
Slice of Life: Porcine Kidney Slices for Testing Antifibrotic Drugs in a Transplant Setting



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Hypothermic Machine Perfusion
2.3. Precision-Cut Kidney Slices
2.4. Inducing Fibrosis in PCKS
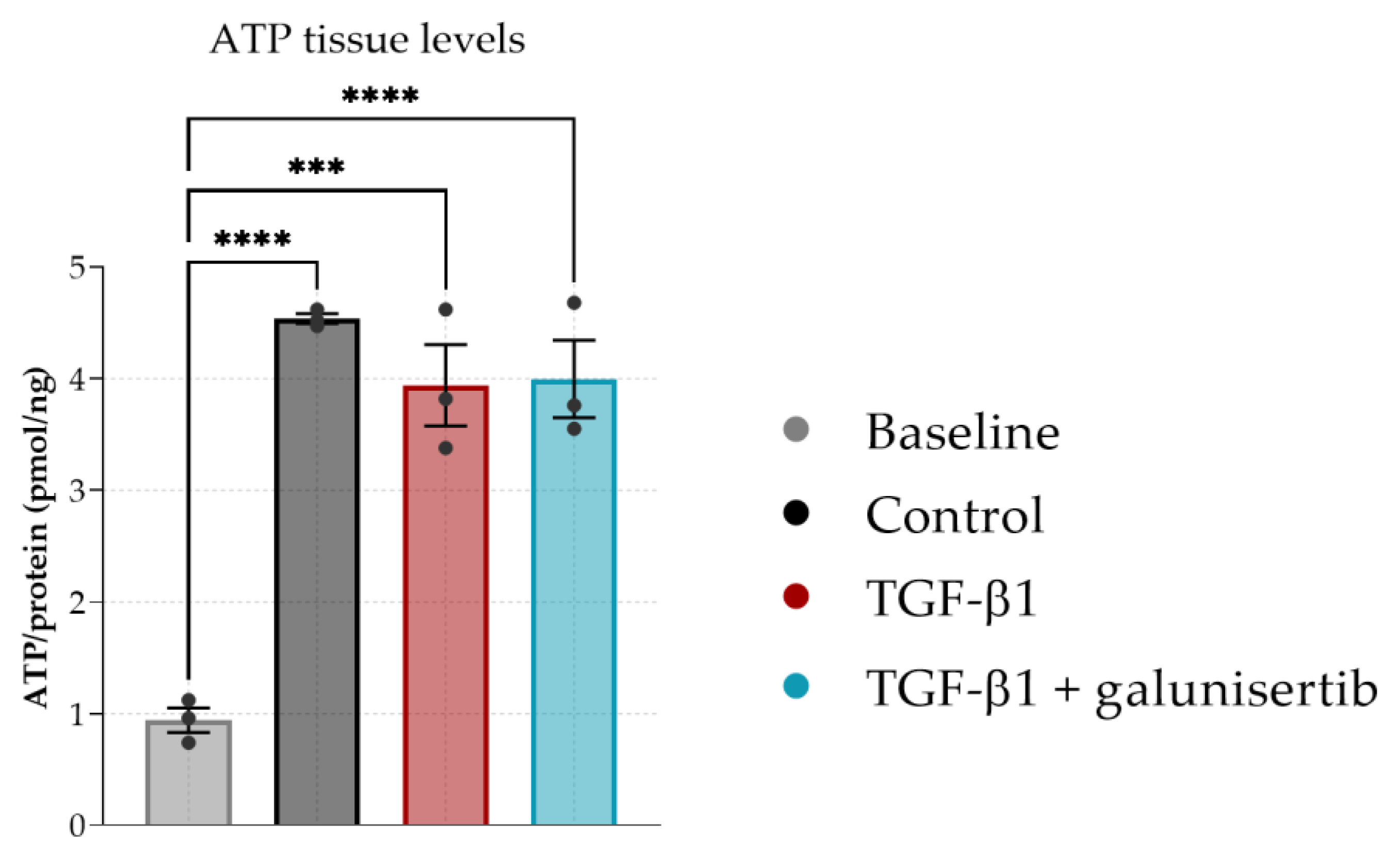
2.5. Viability Analysis
2.6. Gene Expression Analysis
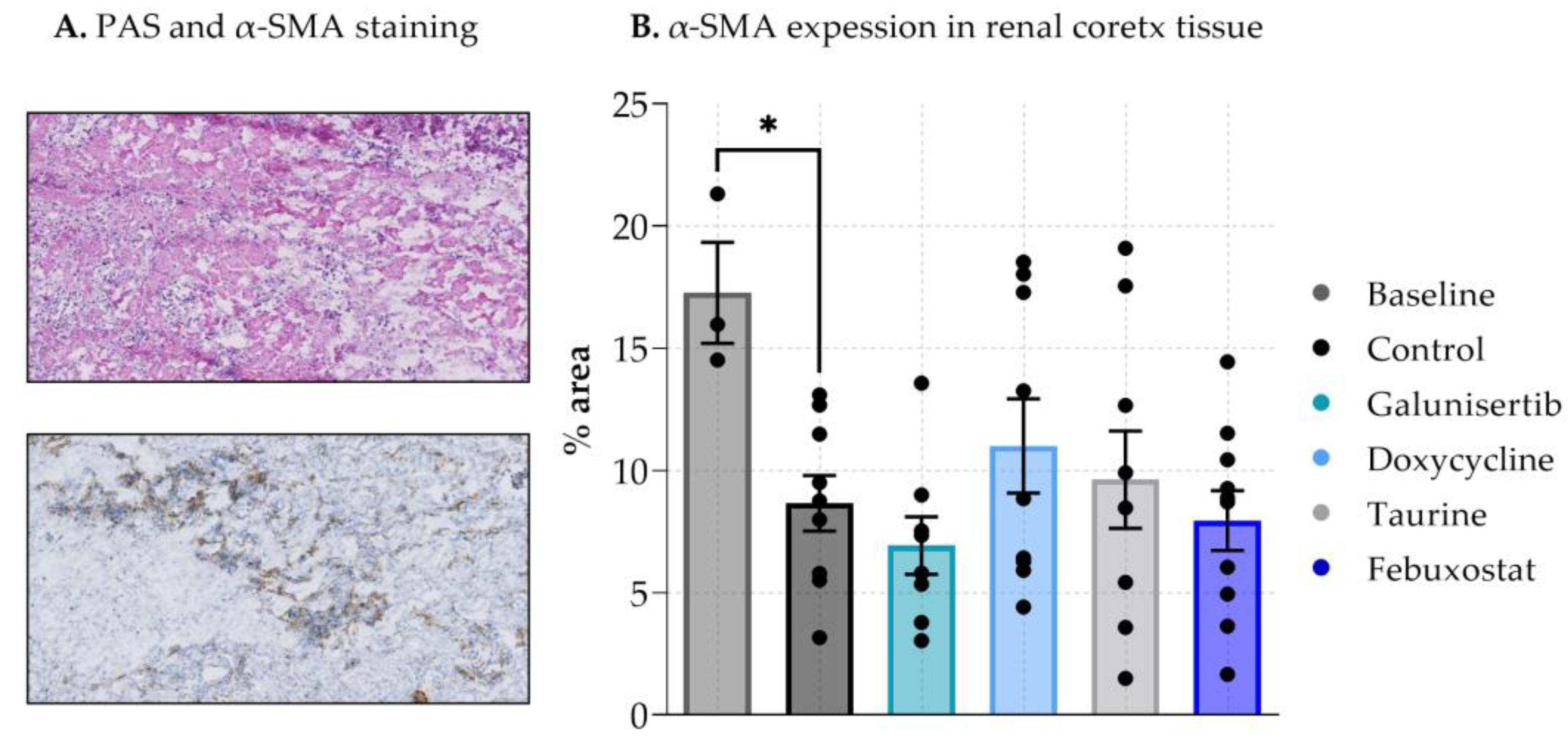
2.7. Histological Analysis
2.8. Statistical Analysis
3. Results
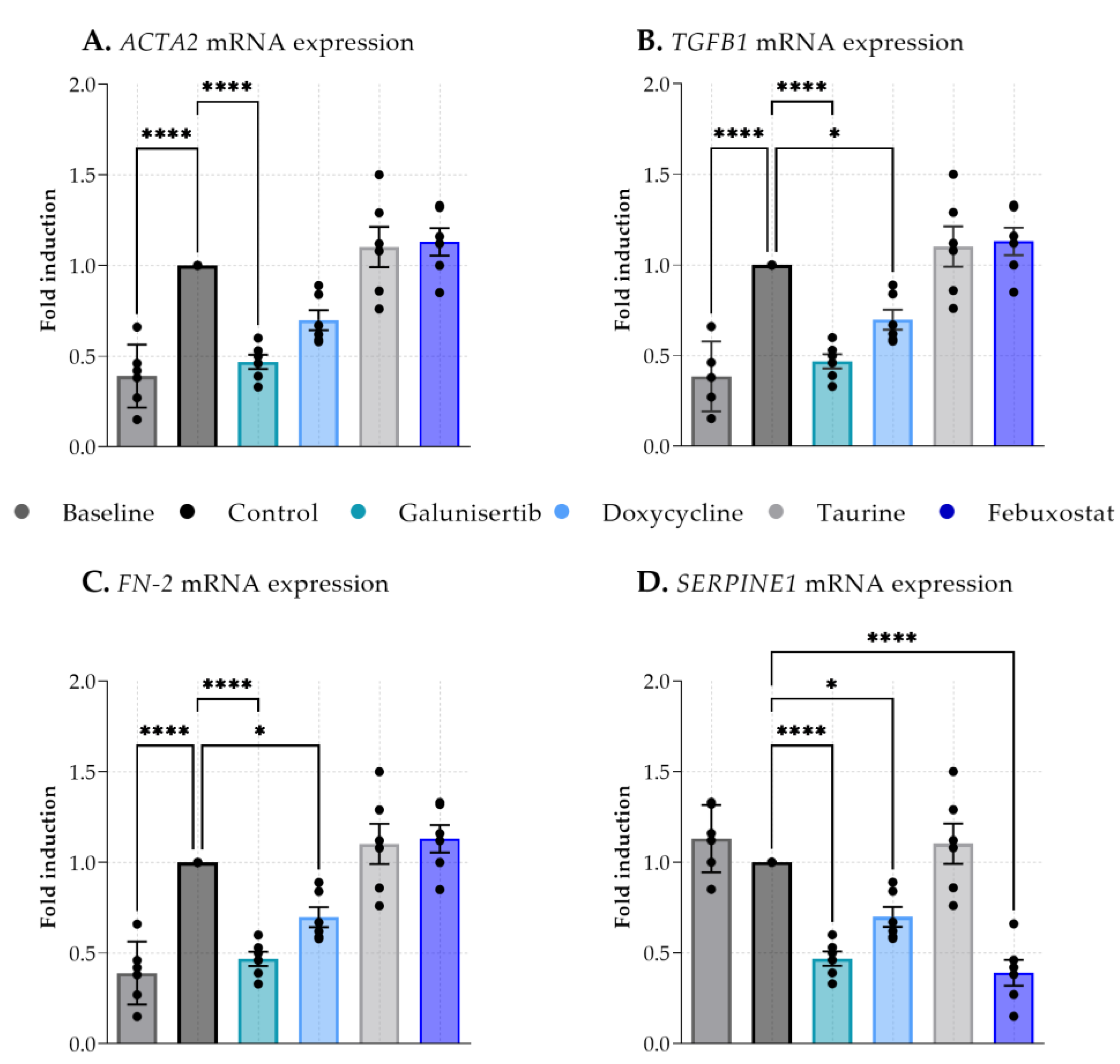
3.1. The Effect of Antifibrotic Drugs on DCD-PCKS
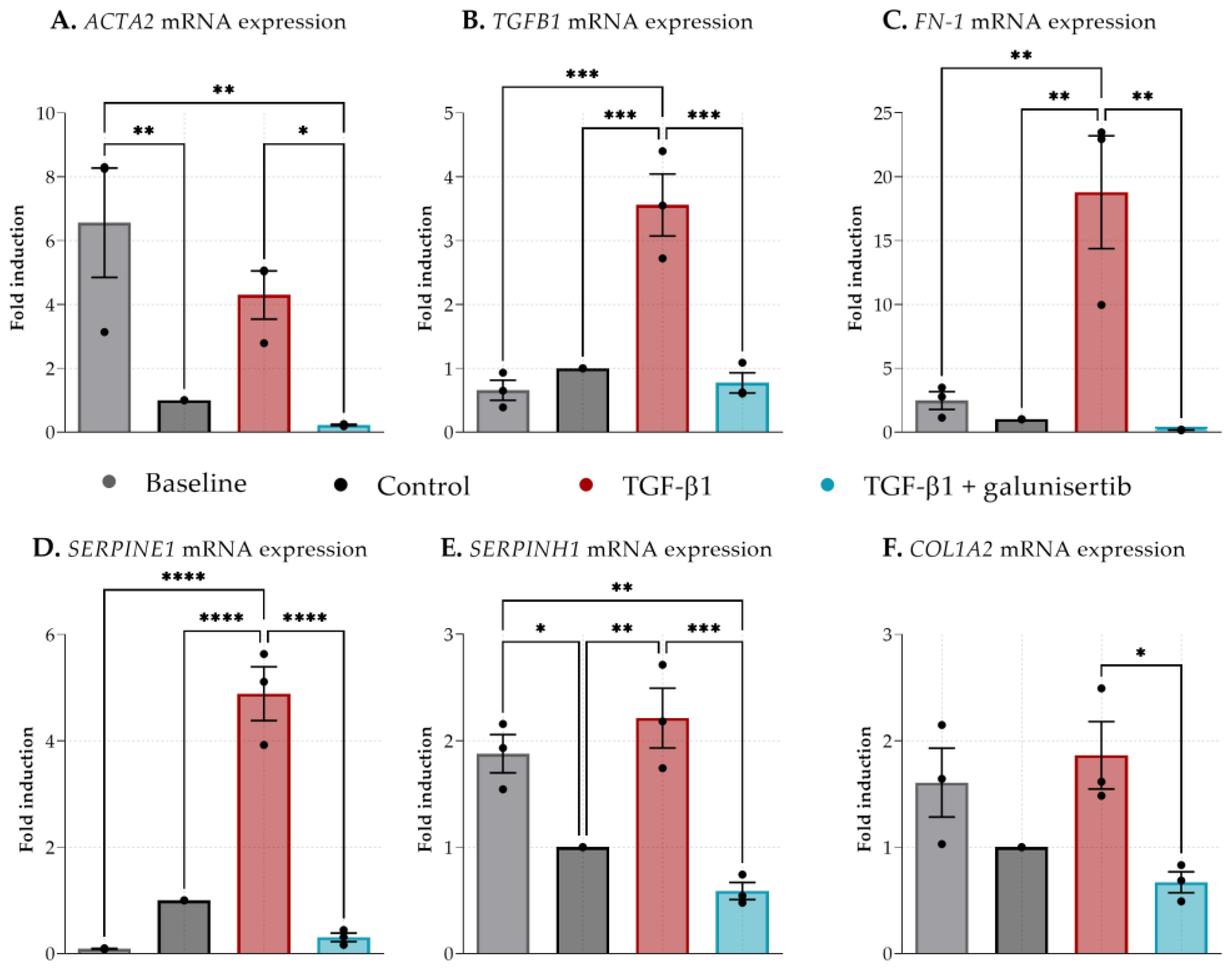
3.2. The Effect of Galunisertib on Fibrotic PCKS
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef] [PubMed]
- Schaubel, D.; Desmeules, M.; Mao, Y.; Jeffery, J.; Fenton, S. Survival experience among elderly end-stage renal disease patients. A controlled comparison of transplantation and dialysis. Transplantation 1995, 60, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Filiopoulos, V.; Boletis, J.N. Renal transplantation with expanded criteria donors: Which is the optimal immunosuppression? World J. Transplant. 2016, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Saidi, R.F.; Elias, N.; Kawai, T.; Hertl, M.; Farrell, M.L.; Goes, N.; Wong, W.; Hartono, C.; Fishman, J.A.; Kotton, C.N.; et al. Outcome of Kidney Transplantation Using Expanded Criteria Donors and Donation After Cardiac Death Kidneys: Realities and Costs. Am. J. Transplant. 2007, 7, 2769–2774. [Google Scholar] [CrossRef] [PubMed]
- Venkat, K.K.; Eshelman, A.K. The evolving approach to ethical issues in living donor kidney transplantation: A review based on illustrative case vignettes. Transplant. Rev. 2014, 28, 134–139. [Google Scholar] [CrossRef]
- Li, X.; Zhuang, S. Recent advances in renal interstitial fibrosis and tubular atrophy after kidney transplantation. Fibrogenesis Tissue Repair 2014, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Boor, P.; Floege, J. Renal Allograft Fibrosis: Biology and Therapeutic Targets. Am. J. Transplant. 2015, 15, 863–886. [Google Scholar] [CrossRef]
- Cosio, F.G.; Grande, J.P.; Larson, T.S.; Gloor, J.M.; Velosa, J.A.; Textor, S.C.; Griffin, M.D.; Stegall, M.D. Kidney Allograft Fibrosis and Atrophy Early After Living Donor Transplantation. Am. J. Transplant. 2005, 5, 1130–1136. [Google Scholar] [CrossRef]
- Yuan, Q.; Tan, R.J.; Liu, Y. Myofibroblast in Kidney Fibrosis: Origin, Activation, and Regulation. Ren. Fibros. Mech. Therapies 2019, 1165, 253–283. [Google Scholar]
- LeBleu, V.; Taduri, G.; O’Connell, J.; Teng, Y.; Cooke, V.; Woda, C.; Sugimoto, H.; Kalluri, R. Origin and function of myofibroblasts in kidney fibrosis. Nat. Med. 2013, 19, 1047–1053. [Google Scholar] [CrossRef]
- Mackinnon, A.; Forbes, S. Bone marrow contributions to fibrosis. Biochim. et Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 955–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Zhang, Z.; Jia, L.; Wang, Y. Role of bone marrow-derived fibroblasts in renal fibrosis. Front. Physiol. 2016, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, A.A. Molecular insights into renal interstitial fibrosis. J. Am. Soc. Nephrol. 1996, 7, 2495–2508. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A. Overview of the cellular and molecular basis of kidney fibrosis. Kidney Int. Suppl. 2014, 4, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Renal fibrosis: New insights into the pathogenesis and therapeutics. Kidney Int. 2006, 69, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Dong, Y.; Tian, X.; Tan, T.K.; Liu, Z.; Zhao, Y.; Zhang, Y.; Harris, D.C.; Zheng, G. Matrix metalloproteinases contribute to kidney fibrosis in chronic kidney diseases. World J. Nephrol. 2013, 2, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.S.; Rasmussen, D.G.K.; Yazdani, S.; Navis, G.; van Goor, H.; Karsdal, M.A.; Genovese, F.; van den Born, J. Urinary collagen degradation products as early markers of progressive renal fibrosis. J. Transl. Med. 2017, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Abrass, C.K.; Hansen, K.M.; Patton, B.L. Laminin α4-Null Mutant Mice Develop Chronic Kidney Disease with Persistent Overexpression of Platelet-Derived Growth Factor. Am. J. Pathol. 2010, 176, 839–849. [Google Scholar] [CrossRef]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.K.; Zheng, G.; Hsu, T.T.; Wang, Y.; Lee, V.W.; Tian, X.; Wang, Y.; Cao, Q.; Wang, Y.; Harris, D.C. Macrophage Matrix Metalloproteinase-9 Mediates Epithelial-Mesenchymal Transition in Vitro in Murine Renal Tubular Cells. Am. J. Pathol. 2010, 176, 1256–1270. [Google Scholar] [CrossRef] [Green Version]
- Kui Tan, T.; Zheng, G.; Hsu, T.T.; Ra Lee, S.; Zhang, J.; Zhao, Y.; Tian, X.; Wang, Y.; Min Wang, Y.; Cao, Q.; et al. Matrix metalloproteinase-9 of tubular and macrophage origin contributes to the pathogenesis of renal fibrosis via macrophage recruitment through osteopontin cleavage. Lab. Investig. 2013, 93, 434–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaka, Y. Targeting TGF-β Signaling in Kidney Fibrosis. Int. J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luangmonkong, T.; Suriguga, S.; Bigaeva, E.; Boersema, M.; Oosterhuis, D.; de Jong, K.P.; Schuppan, D.; Mutsaers, H.A.; Olinga, P. Evaluating the antifibrotic potency of galunisertib in a human ex vivo model of liver fibrosis. Br. J. Pharmacol. 2017, 174, 3107–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stribos, E.G.; Seelen, M.A.; van Goor, H.; Olinga, P.; Mutsaers, H.A. Murine Precision-Cut Kidney Slices as an ex vivo Model to Evaluate the Role of Transforming Growth Factor-β1 Signaling in the Onset of Renal Fibrosis. Front. Physiol. 2017, 8, 1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavdar, Z.A.H.İ.D.E.; Ural, C.; Celik, A.; Arslan, S.; Terzioglu, G.; Ozbal, S.; Yildiz, S.; Ergur, U.B.; Guneli, E.; Camsari, T.; et al. Protective effects of taurine against renal ischemia/reperfusion injury in rats by inhibition of gelatinases, MMP-2 and MMP-9, and p38 mitogen-activated protein kinase signaling. Biotech. Histochem. 2017, 92, 524–535. [Google Scholar] [CrossRef]
- Karbalay-Doust, S.; Noorafshan, A.; Pourshahid, S.M. Taxol and taurine protect the renal tissue of rats after unilateral ureteral obstruction: A stereological survey. Korean J. Urol. 2012, 53, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Yamate, J.; Saito, T.; Hosokawa, T.; Saito, S.; Kurasaki, M. Protective effect of taurine against renal interstitial fibrosis of rats induced by cisplatin. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2002, 365, 277–283. [Google Scholar] [CrossRef]
- Cao, J.; Li, Y.; Peng, Y.; Zhang, Y.; Li, H.; Li, R.; Xia, A. Febuxostat Prevents Renal Interstitial Fibrosis by the Activation of BMP-7 Signaling and Inhibition of USAG-1 Expression in Rats. Am. J. Nephrol. 2015, 42, 369–378. [Google Scholar] [CrossRef]
- Roach, D.M.; Fitridge, R.A.; Laws, P.E.; Millard, S.H.; Varelias, A.; Cowled, P.A. Up-regulation of MMP-2 and MMP-9 Leads to Degradation of Type IV Collagen During Skeletal Muscle Reperfusion Injury; Protection by the MMP Inhibitor, Doxycycline. Eur. J. Vasc. Endovasc. Surg. 2002, 23, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Saglam, F.; Celik, A.; Tayfur, D.; Cavdar, Z.; Yilmaz, O.; Sarioglu, S.; Kolatan, E.; Oktay, G.; Camsari, T. Decrease in cell proliferation by an matrix metalloproteinase inhibitor, doxycycline, in a model of immune-complex nephritis. Nephrology 2010, 15, 560–567. [Google Scholar] [CrossRef]
- Cortes, A.L.; Gonsalez, S.R.; Rioja, L.S.; Oliveira, S.S.; Santos, A.L.; Prieto, M.C.; Melo, P.A.; Lara, L.S. Protective outcomes of low-dose doxycycline on renal function of Wistar rats subjected to acute ischemia/reperfusion injury. Biochim. et Biophys. Acta (BBA)-Mol. Basis Dis. 2018, 1864, 102–114. [Google Scholar] [CrossRef]
- van Leeuwen, L.; Venema, L.H.; Heilig, R.; Leuvenink, H.G.; Kessler, B.M. Doxycycline Alters the Porcine Renal Proteome and Degradome during Hypothermic Machine Perfusion. Curr. Issues Mol. Biol. 2022, 44, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.A.; Arcand, S.; Lin, H.B.; Wojnarowicz, C.; Sawicka, J.; Banerjee, T.; Luo, Y.; Beck, G.R.; Luke, P.P.; Sawicki, G. Protection of the Transplant Kidney from Preservation Injury by Inhibition of Matrix Metalloproteinases. PLoS ONE 2016, 11, e0157508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosgood, S.A.; Nicholson, M.L. First in man renal transplantation after ex vivo normothermic perfusion. Transplantation 2011, 92, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Hosgood, S.A.; Saeb-Parsy, K.; Hamed, M.O.; Nicholson, M.L. Successful Transplantation of Human Kidneys Deemed Untransplantable but Resuscitated by Ex Vivo Normothermic Machine Perfusion. Am. J. Transplant. 2016, 16, 3282–3285. [Google Scholar] [CrossRef] [Green Version]
- Hosgood, S.A.; Thompson, E.; Moore, T.; Wilson, C.H.; Nicholson, M.L. Normothermic machine perfusion for the assessment and transplantation of declined human kidneys from donation after circulatory death donors. J. Br. Surg. 2018, 105, 388–394. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, L.L.; Leuvenink, H.G.D.; Olinga, P.; Ruigrok, M.J.R. Shifting paradigms for suppressing fibrosis in kidney transplants: Supplementing perfusion solutions with antifibrotic drugs. Front. Med. 2022, 8, 2917. [Google Scholar] [CrossRef]
- van Furth, L.A.; Leuvenink, H.G.; Seras, L.; de Graaf, I.A.; Olinga, P.; van Leeuwen, L.L. Exploring Porcine Precision-Cut Kidney Slices as a Model for Transplant-Related Ischemia-Reperfusion Injury. Transplantology 2022, 3, 139–151. [Google Scholar] [CrossRef]
- Stribos, E.G.; Luangmonkong, T.; Leliveld, A.M.; De Jong, I.J.; Van Son, W.J.; Hillebrands, J.L.; Seelen, M.A.; Van Goor, H.; Olinga, P.; Mutsaers, H.A. Precision-cut human kidney slices as a model to elucidate the process of renal fibrosis. Transl. Res. 2016, 170, 8–16. [Google Scholar] [CrossRef]
- de Graaf, I.A.; Olinga, P.; de Jager, M.H.; Merema, M.T.; de Kanter, R.; van de Kerkhof, E.G.; Groothuis, G.M. Preparation and incubation of precision-cut liver and intestinal slices for application in drug metabolism and toxicity studies. Nat. Protoc. 2010, 5, 1540–1551. [Google Scholar] [CrossRef]
- Stribos, E.G.; Hillebrands, J.L.; Olinga, P.; Mutsaers, H.A. Renal fibrosis in precision-cut kidney slices. Eur. J. Pharmacol. 2016, 790, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Matas, A.J.; Gillingham, K.J.; Humar, A.; Kandaswamy, R.; Sutherland, D.E.; Payne, W.D.; Dunn, T.B.; Najarian, J.S. 2202 kidney transplant recipients with 10 years of graft function: What happens next? Am. J. Transplant. 2008, 8, 2410–2419. [Google Scholar] [CrossRef] [Green Version]
- Dinis, P.; Nunes, P.; Marconi, L.; Furriel, F.; Parada, B.; Moreira, P.; Figueiredo, A.; Bastos, C.; Roseiro, A.; Dias, V.; et al. Kidney Retransplantation: Removal or Persistence of the Previous Failed Allograft? Transplant. Proc. 2014, 46, 1730–1734. [Google Scholar] [CrossRef] [PubMed]
- Boor, P.; Šebeková, K.; Ostendorf, T.; Floege, J. Treatment targets in renal fibrosis. Nephrol. Dial. Transplant. 2007, 22, 3391–3407. [Google Scholar] [CrossRef]
- Razzaque, M.S.; Le, V.T.; Taguchi, T. Heat Shock Protein 47 and Renal Fibrogenesis. Cell. Stress Responses Ren. Dis. 2005, 148, 57–69. [Google Scholar]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in Tissue Fibrosis. J. Cell. Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Golriz, M.; Fonouni, H.; Nickkholgh, A.; Hafezi, M.; Garoussi, C.; Mehrabi, A. Pig Kidney Transplantation: An Up-To-Date Guideline. Eur. Surg. Res. 2012, 49, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Grosse-Siestrup, C.; Unger, V.; Fehrenberg, C.; Baeyer, H.V.; Fischer, A.; Schäper, F.; Groneberg, D.A. A model of isolated autologously hemoperfused porcine slaughterhouse kidneys. Nephron 2002, 92, 414–421. [Google Scholar] [CrossRef]
- Venema, L.H.; Brat, A.; Moers, C.; A‘t Hart, N.; Ploeg, R.J.; Hannaert, P.; Minor, T.; Leuvenink, H.G. Effects of Oxygen During Long-term Hypothermic Machine Perfusion in a Porcine Model of Kidney Donation After Circulatory Death. Transplantation 2019, 103, 2057–2064. [Google Scholar] [CrossRef] [Green Version]
- Hendriks, K.D.; Brüggenwirth, I.M.; Maassen, H.; Gerding, A.; Bakker, B.; Porte, R.J.; Henning, R.H.; Leuvenink, H.G. Renal temperature reduction progressively favors mitochondrial ROS production over respiration in hypothermic kidney preservation. J. Transl. Med. 2019, 17, 1–10. [Google Scholar] [CrossRef]
- Pool, M.B.F.; Hartveld, L.; Leuvenink, H.G.D.; Moers, C. Normothermic machine perfusion of ischaemically damaged porcine kidneys with autologous, allogeneic porcine and human red blood cells. PLoS ONE 2020, 15, e0229566. [Google Scholar] [CrossRef] [PubMed]
- Karuthu, S.; Blumberg, E.A. Common infections in kidney transplant recipients. Clin. J. Am. Soc. Nephrol. 2012, 7, 2058–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, M.F.; Liu, Q.; Soliman, B.; Dreher, P.; Okamoto, T.; Poggio, E.D.; Goldfarb, D.A.; Baldwin III, W.M.; Quintini, C. Comparison of normothermic and hypothermic perfusion in porcine kidneys donated after cardiac death. J. Surg. Res. 2017, 216, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Bigaeva, E. Ex Vivo Fibrosis Research: 5 MM Closer to Human Studies; University of Groningen: Groningen, The Netherlands, 2019; pp. 1–301. [Google Scholar]
- Bonora, M.; Patergnani, S.; Rimessi, A.; de Marchi, E.; Suski, J.M.; Bononi, A.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. ATP synthesis and storage. Purinergic Signal 2012, 8, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Cisewski, S.E.; Zhang, L.; Kuo, J.; Wright, G.J.; Wu, Y.; Kern, M.J.; Yao, H. The effects of oxygen level and glucose concentration on the metabolism of porcine TMJ disc cells. Osteoarthr. Cartil. 2015, 23, 1790–1796. [Google Scholar] [CrossRef] [Green Version]
- Brat, A.; Pol, R.A.; Leuvenink, H.G.D. Novel preservation methods to increase the quality of older kidneys. Curr. Opin. Organ Transplant. 2015, 20, 438–443. [Google Scholar] [CrossRef]
- Hosgood, S.A.; van Heurn, E.; Nicholson, M.L. Normothermic machine perfusion of the kidney: Better conditioning and repair? Transpl. Int. 2015, 28, 657–664. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group (n = 7) | Treatment |
|---|---|
| Control | Williams Medium E (1X) + GlutaMAX (WME) (Gibco) + 10 uG/mL ciprofloxacin (Fresenius Kabi, France) + 1.2 mG/mL D-(+)-glucose solution (Sigma-Aldrich, UK) + <1% DMSO |
| Galunisertib | WME + 10 uG/mL ciprofloxacin + 1.2 mG/mL D-(+)-glucose solution + 10 μM galunisertib (Selleckchem, Munich, Germany) in DMSO (<1%) |
| Doxycycline | WME + 10 uG/mL ciprofloxacin + 1.2 mG/mL D-(+)-glucose solution + <1% DMSO + 113 μM doxycycline (Vibramycin SF 100 mg/5 mL, Pfizer BV, Capelle aan den Ijssel, The Netherlands) |
| Taurine | WME + 10 uG/mL ciprofloxacin + 1.2 mG/mL D-(+)-glucose solution + <1% DMSO + 80 mM taurine (Sigma Aldrich, Darmstadt, Germany) dissolved in WME |
| Febuxostat | WME + 10 uG/mL ciprofloxacin + 1.2 mG/mL D-(+)-glucose solution + 16 μM febuxostat (Adenuric 80 mg, Menarini, Florence, Italy) dissolved in DMSO (<1%) |
| Experimental Group (n = 3) | Treatment |
|---|---|
| Control | DMEM/F-12 medium (Gibco, 31331-028) + 10 uG/mL ciprofloxacin + <1% DMSO |
| TGF-β1 | DMEM/F-12 medium + 10 uG/mL ciprofloxacin + 5 ng/mL TGF-β1 (Roche Diagnostics, Almere The Netherlands) + <1% DMSO |
| TGF-β1 + galunisertib | DMEM/F-12 medium + 10 uG/mL ciprofloxacin + 5 ng/mL TGF-β1 + 10 μM galunisertib in DMSO (<1%) |
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| GAPDH | CCTGCCGTCTGGAGAAACC | CTCGGACGCCTGCTTCA |
| TGF-b1 | GGGAGGGTGTTCATGGTAGGA | AGCTCACCCCAAATTCATCTTC |
| ACTA2 | ACGAAGCCCAAAGCAAAAGA | GTTGGTGATGATGCCGTGTTC |
| SERPINE1 | GCAAGTTCGGGCTCCACTAC | TGCATGCCGTAACCTCCTG |
| Fibronectin 2 | TTAACTGTCTGGCCCGCAATAT | AGGCAATTACCAAAGTCATCTGGA |
| COL1A2 | CAAGAAAGGGCCCAACTGGA | AGGGCCTGGGATACCATCAT |
| Fibronectin 1 | GCACCATCCAACTTGCGTTT | TGTACTCGGTTGCTGGTTCC |
| SERPINH1 | TGCAGTCCATCAACGAGTGG | TGGAATCGCTCATCCCAGTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Leeuwen, L.L.; Ruigrok, M.J.R.; Leuvenink, H.G.D.; Olinga, P. Slice of Life: Porcine Kidney Slices for Testing Antifibrotic Drugs in a Transplant Setting. Transplantology 2023, 4, 59-70. https://doi.org/10.3390/transplantology4020007
van Leeuwen LL, Ruigrok MJR, Leuvenink HGD, Olinga P. Slice of Life: Porcine Kidney Slices for Testing Antifibrotic Drugs in a Transplant Setting. Transplantology. 2023; 4(2):59-70. https://doi.org/10.3390/transplantology4020007
Chicago/Turabian Stylevan Leeuwen, L. Leonie, Mitchel J. R. Ruigrok, Henri G. D. Leuvenink, and Peter Olinga. 2023. "Slice of Life: Porcine Kidney Slices for Testing Antifibrotic Drugs in a Transplant Setting" Transplantology 4, no. 2: 59-70. https://doi.org/10.3390/transplantology4020007