
Photocurrent Production from Cherries in a Bio-Electrochemical Cell


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Fluorescence Measurements
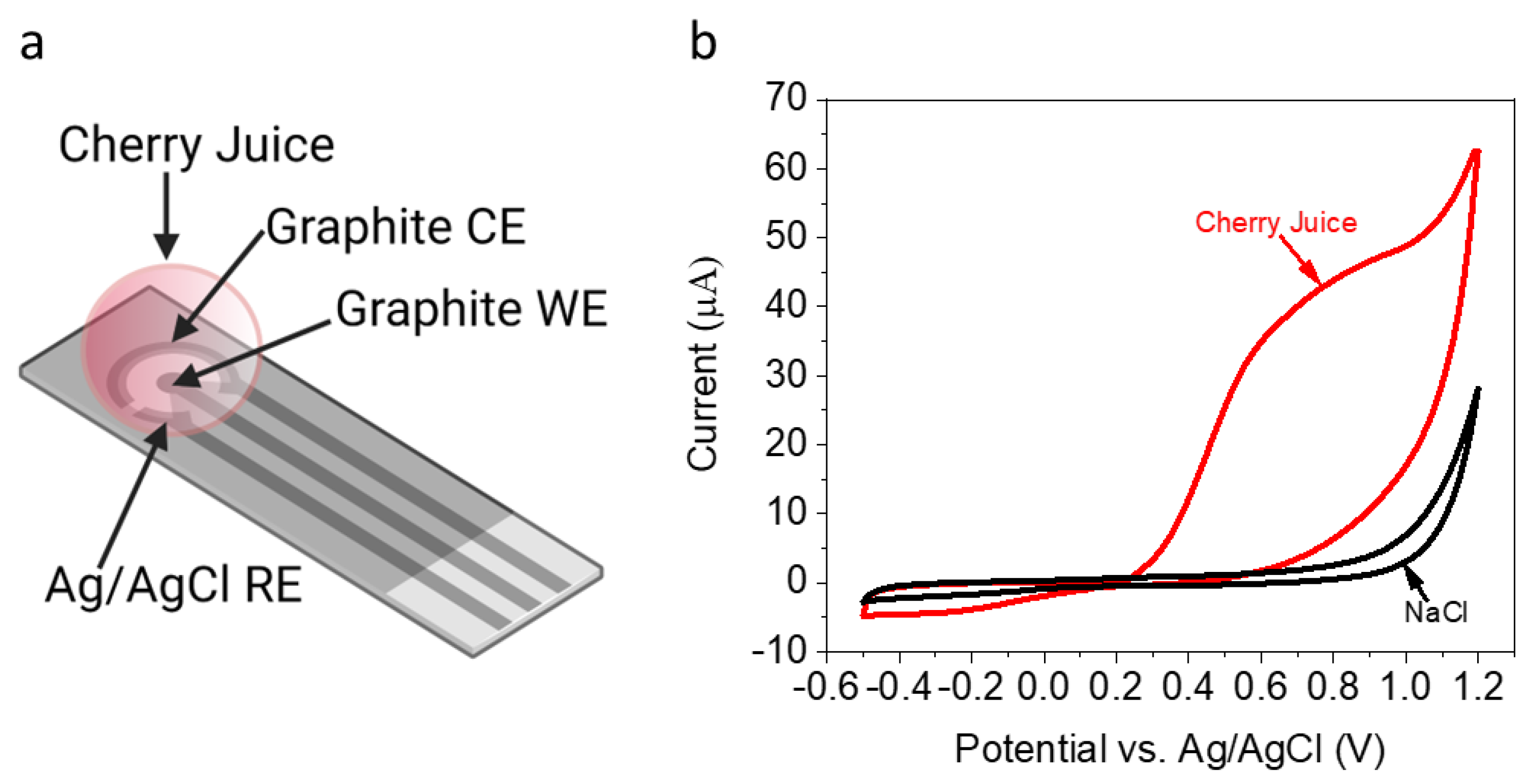
2.3. Electrochemical Apparatus
2.4. Chronoamperometry
2.5. Cyclic Voltammetry
2.6. Electrochemical Impedance Spectroscopy (EIS)
3. Results and Discussion
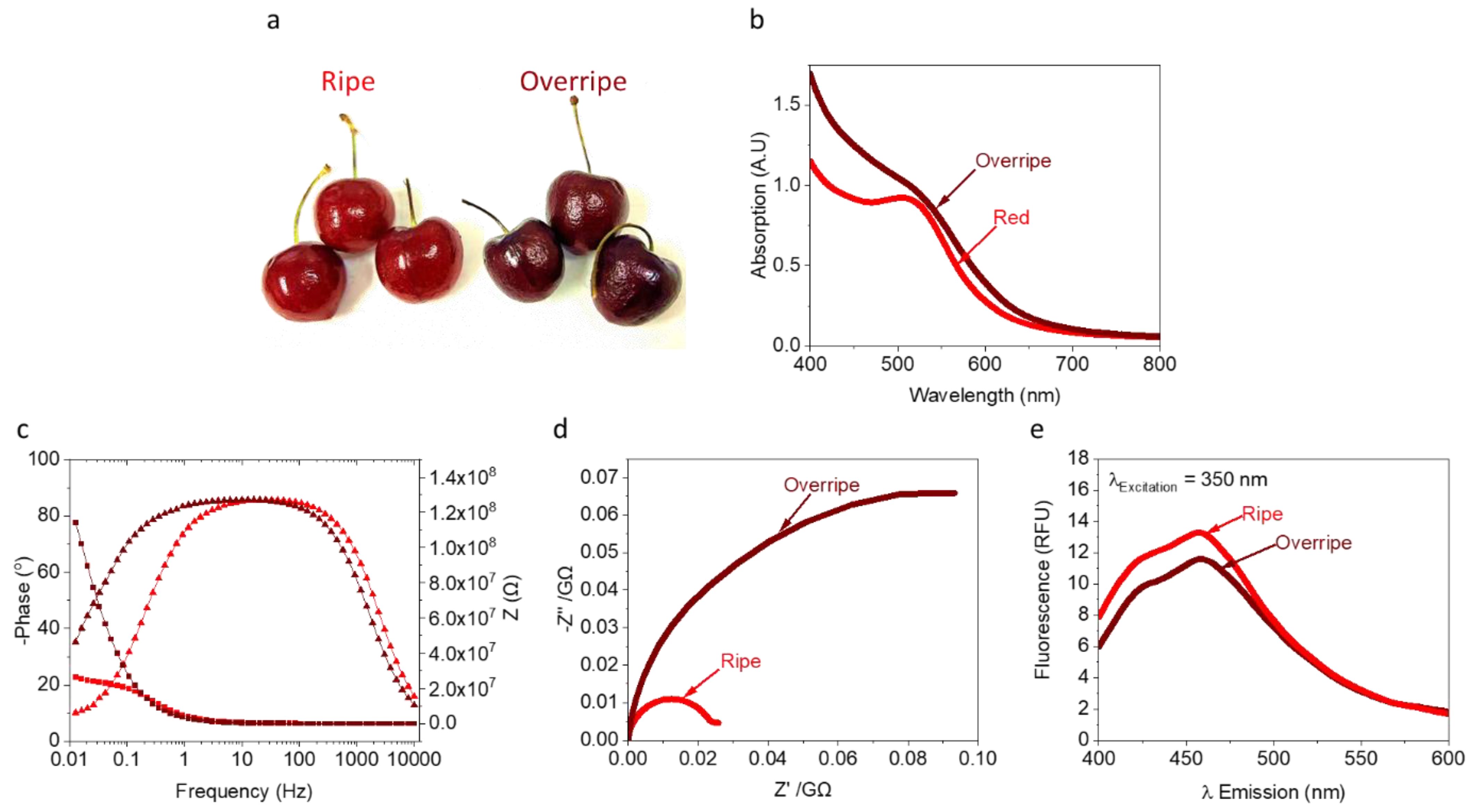
3.1. Identification of Redox-Active Molecules in a Cherry
3.2. Analysis of the Internal Electrolyte in Ripe and Overripe Cherries
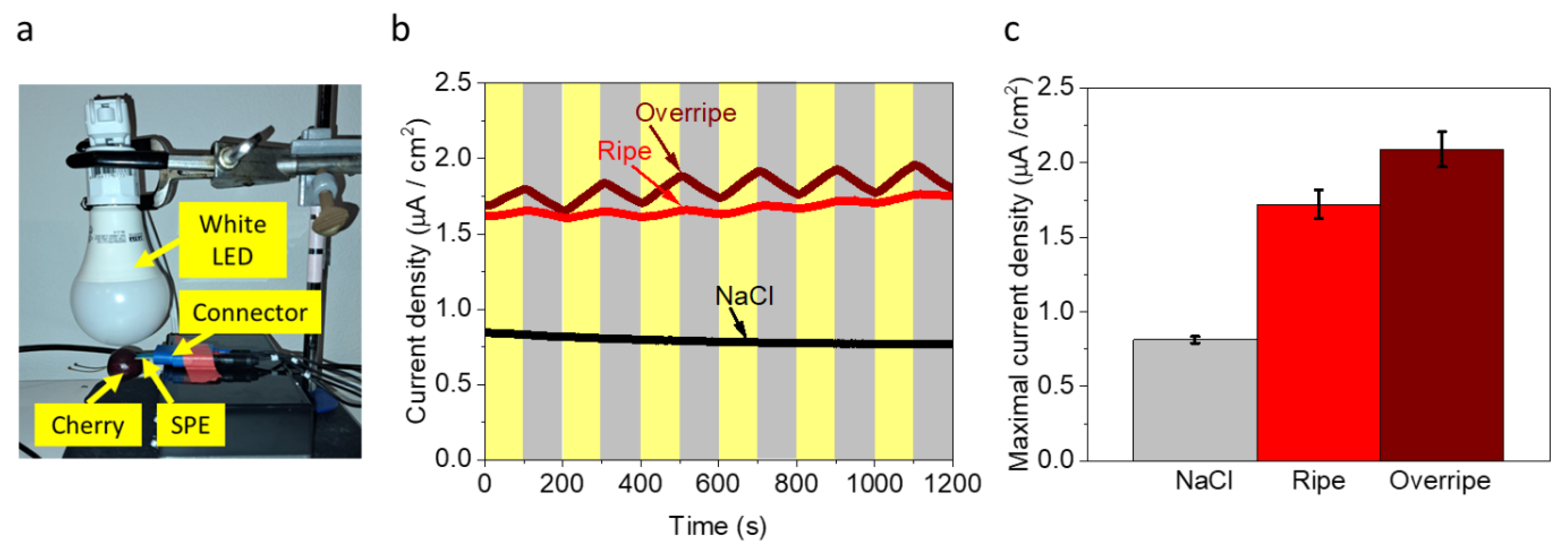
3.3. Direct Photocurrent Production from a Cherry
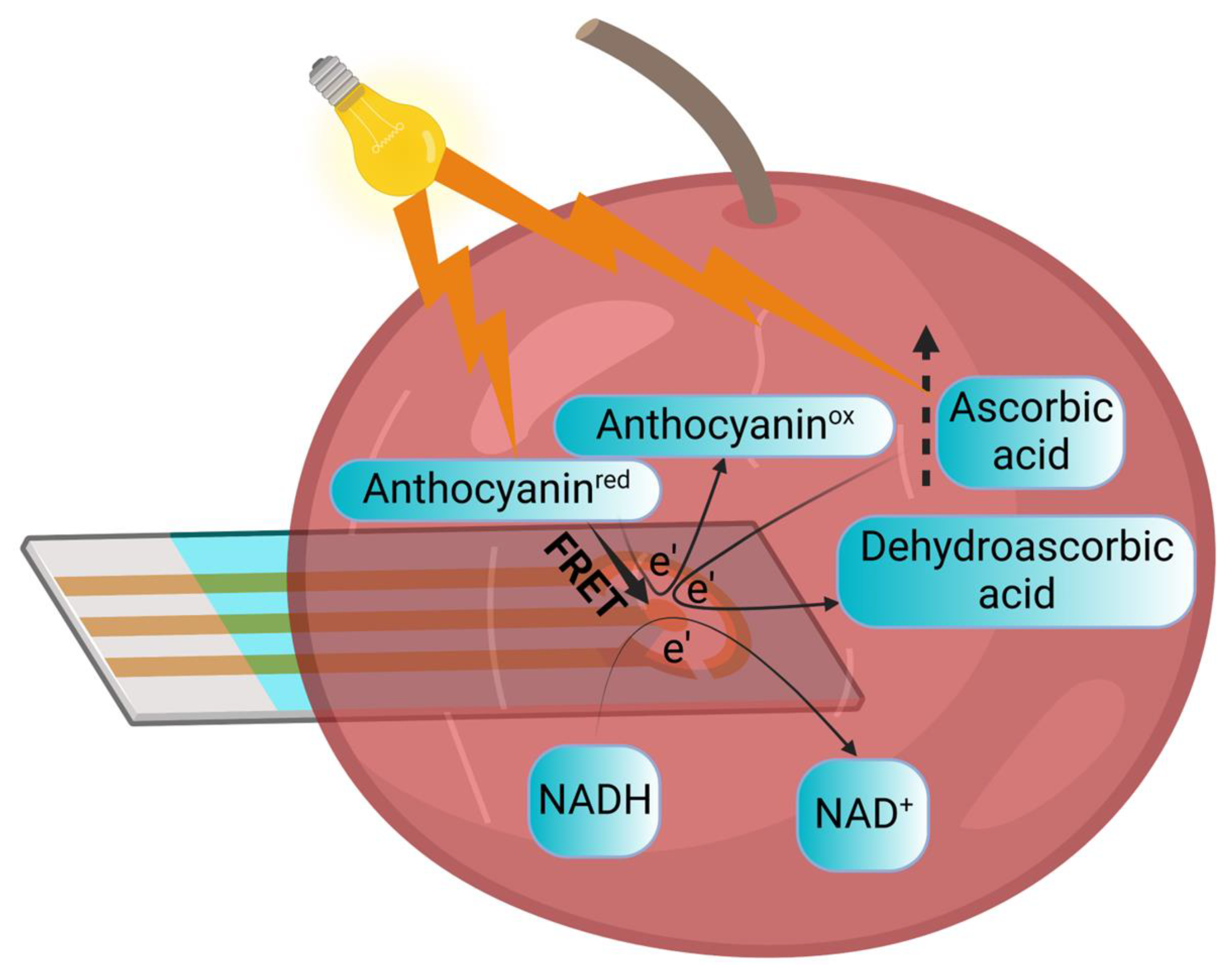
3.4. Proposed Mechanism for Electrical Current and Photocurrent Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obileke, K.; Onyeaka, H.; Meyer, E.L.; Nwokolo, N. Microbial Fuel Cells, a Renewable Energy Technology for Bio-Electricity Generation: A Mini-Review. Electrochem. Commun. 2021, 125, 107003. [Google Scholar] [CrossRef]
- Hartshorne, R.S.; Jepson, B.N.; Clarke, T.A.; Field, S.J.; Fredrickson, J.; Zachara, J.; Shi, L.; Butt, J.N.; Richardson, D.J. Characterization of Shewanella Oneidensis MtrC: A Cell-Surface Decaheme Cytochrome Involved in Respiratory Electron Transport to Extracellular Electron Acceptors. JBIC J. Biol. Inorg. Chem. 2007, 12, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Rosso, K.M.; Zachara, J.M.; Fredrickson, J.K. Mtr Extracellular Electron-Transfer Pathways in Fe(III)-Reducing or Fe(II)-Oxidizing Bacteria: A Genomic Perspective. Biochem. Soc. Trans. 2012, 40, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Heidary, N.; Kornienko, N.; Kalathil, S.; Fang, X.; Ly, K.H.; Greer, H.F.; Reisner, E. Disparity of Cytochrome Utilization in Anodic and Cathodic Extracellular Electron Transfer Pathways of Geobacter Sulfurreducens Biofilms. J. Am. Chem. Soc. 2020, 142, 5194–5203. [Google Scholar] [CrossRef]
- Lovley, D.R. Electromicrobiology. Annu. Rev. Microbiol. 2012, 66, 391–409. [Google Scholar] [CrossRef]
- Nevin, K.P.; Richter, H.; Covalla, S.F.; Johnson, J.P.; Woodard, T.L.; Orloff, A.L.; Jia, H.; Zhang, M.; Lovley, D.R. Power Output and Columbic Efficiencies from Biofilms of Geobacter Sulfurreducens Comparable to Mixed Community Microbial Fuel Cells. Environ. Microbiol. 2008, 10, 2505–2514. [Google Scholar] [CrossRef]
- Yi, H.; Nevin, K.P.; Kim, B.-C.; Franks, A.E.; Klimes, A.; Tender, L.M.; Lovley, D.R. Selection of a Variant of Geobacter Sulfurreducens with Enhanced Capacity for Current Production in Microbial Fuel Cells. Biosens. Bioelectron. 2009, 24, 3498–3503. [Google Scholar] [CrossRef]
- Neu, J.; Shipps, C.C.; Guberman-Pfeffer, M.J.; Shen, C.; Srikanth, V.; Spies, J.A.; Kirchhofer, N.D.; Yalcin, S.E.; Brudvig, G.W.; Batista, V.S.; et al. Microbial Biofilms as Living Photoconductors Due to Ultrafast Electron Transfer in Cytochrome OmcS Nanowires. Nat. Commun. 2022, 13, 5150. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Limwongyut, J.; Moreland, A.S.; Bazan, G.C. Non-Photosynthetic Bacteria Produce Photocurrent Mediated by NADH. bioRxiv 2023. [Google Scholar] [CrossRef]
- Simoska, O.; Sans, M.; Eberlin, L.S.; Shear, J.B.; Stevenson, K.J. Electrochemical Monitoring of the Impact of Polymicrobial Infections on Pseudomonas Aeruginosa and Growth Dependent Medium. Biosens. Bioelectron. 2019, 142, 111538. [Google Scholar] [CrossRef]
- Ieropoulos, I.; Greenman, J.; Melhuish, C.; Hart, J. Comparative Study of Three Types of Microbial Fuel Cell. Enzyme Microb. Technol. 2005, 37, 238–245. [Google Scholar] [CrossRef]
- Rabaey, K.; Boon, N.; Siciliano, S.D.; Verhaege, M.; Verstraete, W. Biofuel Cells Select for Microbial Consortia That Self-Mediate Electron Transfer. Appl. Environ. Microbiol. 2004, 70, 5373–5382. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Boon, N.; Höfte, M.; Verstraete, W. Microbial Phenazine Production Enhances Electron Transfer in Biofuel Cells. Environ. Sci. Technol. 2005, 39, 3401–3408. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Coates, J.D.; Blunt-Harris, E.L.; Phillips, E.J.P.; Woodward, J.C. Humic Substances as Electron Acceptors for Microbial Respiration. Nature 1996, 382, 445–448. [Google Scholar] [CrossRef]
- Lovley, D.R.; Holmes, D.E.; Nevin, K.P. Dissimilatory Fe(III) and Mn(IV) Reduction; Academic Press: Cambridge, MA, USA, 2004; Volume 49, pp. 219–286. ISBN 0065-2911. [Google Scholar]
- Herkendell, K. Status Update on Bioelectrochemical Systems: Prospects for Carbon Electrode Design and Scale-Up. Catalysts 2021, 11, 278. [Google Scholar] [CrossRef]
- Herkendell, K.; Tel-Vered, R.; Stemmer, A. Switchable Aerobic/Anaerobic Multi-Substrate Biofuel Cell Operating on Anodic and Cathodic Enzymatic Cascade Assemblies. Nanoscale 2017, 9, 14118–14126. [Google Scholar] [CrossRef]
- Herkendell, K.; Stemmer, A.; Tel-Vered, R. Extending the Operational Lifetimes of All-Direct Electron Transfer Enzymatic Biofuel Cells by Magnetically Assembling and Exchanging the Active Biocatalyst Layers on Stationary Electrodes. Nano Res. 2019, 12, 767–775. [Google Scholar] [CrossRef]
- Herkendell, K.; Stemmer, A.; Tel-Vered, R. Magnetically Induced Enzymatic Cascades—Advancing towards Multi-Fuel Direct/Mediated Bioelectrocatalysis. Nanoscale Adv. 2019, 1, 1686–1692. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Spungin, D.; Schuster, G.; Frank, I.-B.; Adir, N. Trichodesmium Erythraeum Produces a Higher Photocurrent than Other Cyanobacterial Species in Bio-Photo Electrochemical Cells. Biochim. Biophys. Acta Bioenerg. 2022, 1863, 148910. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Eichenbaum, B.; Tóth, T.N.; Levin, G.; Liveanu, V.; Schuster, G.; Adir, N. NADPH Performs Mediated Electron Transfer in Cyanobacterial-Driven Bio-Photoelectrochemical Cells. iScience 2020, 24, 101892. [Google Scholar] [CrossRef]
- Saper, G.; Kallmann, D.; Conzuelo, F.; Zhao, F.; Tóth, T.N.; Liveanu, V.; Meir, S.; Szymanski, J.; Aharoni, A.; Schuhmann, W.; et al. Live Cyanobacteria Produce Photocurrent and Hydrogen Using Both the Respiratory and Photosynthetic Systems. Nat. Commun. 2018, 9, 2168. [Google Scholar] [CrossRef] [PubMed]
- Karkos, P.D.; Leong, S.C.; Karkos, C.D.; Sivaji, N.; Assimakopoulos, D.A. Spirulina in Clinical Practice: Evidence-Based Human Applications. Evid.-Based Complement. Altern. Med. 2011, 2011, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Shlosberg, Y.; Tóth, T.N.; Eichenbaum, B.; Keysar, L.; Schuster, G.; Adir, N. Electron Mediation and Photocurrent Enhancement in Dunalliela Salina Driven Bio-Photo Electrochemical Cells. Catalysts 2021, 11, 1220. [Google Scholar] [CrossRef]
- McCormick, A.J.J.; Bombelli, P.; Scott, A.M.M.; Philips, A.J.J.; Smith, A.G.G.; Fisher, A.C.C.; Howe, C.J.J. Photosynthetic Biofilms in Pure Culture Harness Solar Energy in a Mediatorless Bio-Photovoltaic Cell (BPV) System. Energy Environ. Sci. 2011, 4, 4699–4709. [Google Scholar] [CrossRef]
- Santoro, C.; Arbizzani, C.; Erable, B.; Ieropoulos, I. Microbial Fuel Cells: From Fundamentals to Applications. A Review. J. Power Sources 2017, 356, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Ng, F.L.; Phang, S.M.; Periasamy, V.; Yunus, K.; Fisher, A.C. Enhancement of Power Output by Using Alginate Immobilized Algae in Biophotovoltaic Devices. Sci. Rep. 2017, 7, 16237. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Krupnik, N.; Tóth, T.N.; Eichenbaum, B.; Meirovich, M.; Meiri, D.; Yehezkeli, O.; Schuster, G.; Israel, Á.; Adir, N. Bioelectricity Generation from Live Marine Photosynthetic Macroalgae. Biosens. Bioelectron. 2021, 198, 113824. [Google Scholar] [CrossRef]
- Hubenova, Y.; Mitov, M. Conversion of Solar Energy into Electricity by Using Duckweed in Direct Photosynthetic Plant Fuel Cell. Bioelectrochemistry 2012, 87, 185–191. [Google Scholar] [CrossRef]
- Hubenova, Y.; Mitov, M. Enhanced Metabolic and Redox Activity of Vascular Aquatic Plant Lemna Valdiviana under Polarization in Direct Photosynthetic Plant Fuel Cell. Bioelectrochemistry 2015, 106, 226–231. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Meirovich, M.; Yehezkeli, O.; Schuster, G.; Adir, N. Production of Photocurrent and Hydrogen Gas from Intact Plant Leaves. Biosens. Bioelectron. 2022, 215, 114558. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Schuster, G.; Adir, N. Self-Enclosed Bio-Photoelectrochemical Cell in Succulent Plants. ACS Appl. Mater. Interfaces 2022, 14, 53761–53766. [Google Scholar] [CrossRef] [PubMed]
- Shlosberg, Y.; Schuster, G.; Adir, N. Harnessing Photosynthesis to Produce Electricity Using Cyanobacteria, Green Algae, Seaweeds and Plants. Front. Plant Sci. 2022, 13, 955843. [Google Scholar] [CrossRef] [PubMed]
- Gvamichava, N.E. Vitamin Secretion by Roots. Fiziol. Drev. Rastenii 1966, 2, 5–16. [Google Scholar]
- Pardha-Saradhi, P.; Yamal, G.; Peddisetty, T.; Sharmila, P.; Nagar, S.; Singh, J.; Nagarajan, R.; Rao, K.S. Reducing Strength Prevailing at Root Surface of Plants Promotes Reduction of Ag+ and Generation of Ag0/Ag2O Nanoparticles Exogenously in Aqueous Phase. PLoS ONE 2014, 9, e106715. [Google Scholar] [CrossRef]
- Shlosberg, Y. Direct Electricity Production from Green Onion’s Roots. bioRxiv 2022. [Google Scholar] [CrossRef]
- Apollon, W.; Luna-Maldonado, A.I.; Kamaraj, S.-K.; Vidales-Contreras, J.A.; Rodríguez-Fuentes, H.; Gómez-Leyva, J.F.; Aranda-Ruíz, J. Progress and Recent Trends in Photosynthetic Assisted Microbial Fuel Cells: A Review. Biomass Bioenergy 2021, 148, 106028. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Brekhman, V.; Lotan, T.; Sepunaru, L. Direct Electricity Production from Nematostella and Arthemia’s Eggs in a Bio-Electrochemical Cell. Int. J. Mol. Sci. 2022, 23, 15001. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Sepunaru, L. Advantages of Imprinted Polymer Electrodes for Electrochemical Pathogen Detection. Curr. Opin. Electrochem. 2022, 36, 101123. [Google Scholar] [CrossRef]
- Li, M.; Li, Y.-T.; Li, D.-W.; Long, Y.-T. Recent Developments and Applications of Screen-Printed Electrodes in Environmental Assays—A Review. Anal. Chim. Acta 2012, 734, 31–44. [Google Scholar] [CrossRef]
- Shlosberg, Y.; Smith, M.; Carlini, A. Redox-Active Molecules in Bacterial Cultivation Media Produce Photocurrent. bioRxiv 2022. [Google Scholar] [CrossRef]
- Šic Žlabur, J.; Bogdanović, S.; Voća, S.; Skendrović Babojelić, M. Biological Potential of Fruit and Leaves of Strawberry Tree (Arbutus Unedo L.) from Croatia. Molecules 2020, 25, 5102. [Google Scholar] [CrossRef] [PubMed]
- Blandón-Naranjo, L.; Della Pelle, F.; Vázquez, M.V.; Gallego, J.; Santamaría, A.; Alzate-Tobón, M.; Compagnone, D. Electrochemical Behaviour of Microwave-Assisted Oxidized MWCNTs Based Disposable Electrodes: Proposal of a NADH Electrochemical Sensor. Electroanalysis 2018, 30, 509–516. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Danet, A.F.; Kalinowski, S. Ascorbic Acid Determination in Commercial Fruit Juice Samples by Cyclic Voltammetry. J. Autom. Methods Manag. Chem. 2008, 2008, 937651. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, M.; Torabi, S.; Tavan, M.; Azizi, A.; Khazalpour, S. Electrochemical Behavior and LC-MS Analysis of Anthocyanin’s in Vaccinium Arctostaphylos L. Extract: The Molecular Modelling of Potential Inhibition to COVID-19 and ROS Generation Receptors. J. Electrochem. Soc. 2020, 167, 155505. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Singh, P.; Bera, T.K.; Ghoshal, D.; Chakraborty, B. Electrical Impedance Spectroscopic Study of Mandarin Orange during Ripening. J. Food Meas. Charact. 2017, 11, 1654–1664. [Google Scholar] [CrossRef]
- Pantelidis, G.E.; Vasilakakis, M.; Manganaris, G.A.; Diamantidis, G. Antioxidant Capacity, Phenol, Anthocyanin and Ascorbic Acid Contents in Raspberries, Blackberries, Red Currants, Gooseberries and Cornelian Cherries. Food Chem. 2007, 102, 777–783. [Google Scholar] [CrossRef]
- Wang, S.Y. Effect of Pre-Harvest Conditions on Antioxidant Capicity in Fruits. In Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 30 June 2006; pp. 299–306. [Google Scholar]
- Singh, V.; Mishra, A.K. White Light Emission from Vegetable Extracts. Sci. Rep. 2015, 5, 11118. [Google Scholar] [CrossRef] [Green Version]
- Potter, M.C. Electrical Effects Accompanying the Decomposition of Organic Compounds. Proc. R. Soc. Lond. B 1911, 84, 260–286. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shlosberg, Y.; Rubino, K.C.; Nasseri, N.S.; Carlini, A.S. Photocurrent Production from Cherries in a Bio-Electrochemical Cell. Electrochem 2023, 4, 47-55. https://doi.org/10.3390/electrochem4010005
Shlosberg Y, Rubino KC, Nasseri NS, Carlini AS. Photocurrent Production from Cherries in a Bio-Electrochemical Cell. Electrochem. 2023; 4(1):47-55. https://doi.org/10.3390/electrochem4010005
Chicago/Turabian StyleShlosberg, Yaniv, Kimi C. Rubino, Nathan S. Nasseri, and Andrea S. Carlini. 2023. "Photocurrent Production from Cherries in a Bio-Electrochemical Cell" Electrochem 4, no. 1: 47-55. https://doi.org/10.3390/electrochem4010005