1. Introduction
Posidonia oceanica (L) Delile is a Mediterranean endemic seagrass and, in optimal conditions, is considered the main habitat-forming species of one of the climax communities of some Mediterranean infralittoral areas due to the high productivity, complexity and stability of this vascular plant [
1,
2]. Meadows of this seagrass have an estimated coverage of 25–50% of infralittoral bottoms of the Mediterranean Sea (down to 50 m in certain areas), corresponding to the most extensive seagrass meadows for this basin [
2,
3]. These meadows support a highly diverse associated biotic community [
1], providing an elevated number of ecosystem services such as the maintenance of marine biodiversity (including species of commercial interest), buffering of strong water movement leading to protection of the coastline against erosion, nutrient cycling and carbon sequestration, among others [
2,
4,
5,
6,
7]. Despite all these services,
P. oceanica has suffered a strong decline over the last centuries [
7,
8,
9], with an estimated regression of meadows at 34% in the last 50 years as a consequence of the cumulative effects of multiple local stressors [
10].
The decline and negative impacts suffered by this ecologically important species have led the EC to include P. oceanica meadows as priority habitat Posidonia beds (Posidonion oceanicae) (Habitat 1120) for conservation purposes within the EU Habitats Directive (92/43/EC). In addition, it is an indicator species and habitat for evaluating the “Good Environmental Status” of European waters in line with the Marine Strategy Framework Directive (MSFD, 2008/56/EC). Therefore, it is necessary to study the phenological parameters and the spatio-temporal variability of P. oceanica meadows across its biogeographical distribution, including the boundaries of its geographical distribution, as a preliminary step for further monitoring programs in order to improve the knowledge and management of this important marine ecosystem element.
Posidonia oceanica has its westernmost distribution limit close to the Strait of Gibraltar (SoG) in the northern Alboran Sea, being generally absent along the Moroccan coastline except for the Chafarinas Islands (35°05′ N, 02°25′ E) [
10,
11]. At its easternmost limit,
P. oceanica occurs in a few localities along the southern Levantine part of Turkey close to the Kizilliman marine protected area and Alexandria (Egypt) on the southern coast [
10,
12,
13]. In the Alboran Sea,
P. oceanica forms extensive beds in its easternmost sector, close to the Almeria-Oran Front. In the central and western sectors, these beds are reduced from usually fragmented meadows to small patches [
5,
11]. Despite the fragmentation experienced by
P. oceanica meadows close to the SoG, several authors have documented highly diverse associated invertebrate assemblages (e.g., molluscs [
14] and crustacean decapods [
15]). This fosters the idea that
P. oceanica patches have an important ecological and conservation value whatever their size, as even small patches comprising a low number of shoots still support a higher abundance and diversity of invertebrates than adjacent sedimentary bottoms, as observed in other seagrass species [
14,
15].
The Alboran Sea displays transitional characteristics between the Atlantic Ocean and the Mediterranean Sea, with a predominance of colder and less salty water masses than other Mediterranean areas [
16]. Moreover, the entrance of surface Atlantic waters through the SoG generates a series of anticyclonic gyres due to several physical factors (northeast orientation of the SoG, the Coriolis effect, the topography of the Alboran basin, the local climatology) that produce upwellings of cold and nutrient enriched Mediterranean deep waters in the northwestern Alboran coasts [
17,
18] (
Figure 1). This sector of the Mediterranean Sea supports one of the highest productive areas within the region [
19,
20], which, together with the complex seafloor heterogeneity and the geographical location of the Alboran basin, promotes a high diversity of habitats and species [
21,
22], including ones of commercial importance. The latter has allowed the development of an intensive fishing activity targeting demersal, small pelagic and benthic species [
23,
24,
25,
26], with potentially adverse effects on vegetated benthic assemblages [
27,
28].
Meadows of
P. oceanica have been widely studied in the Mediterranean Sea, with most studies being focused on seasonal and monthly variations [
28,
29,
30,
31,
32,
33], whereas few of them have considered interannual differences [
34,
35,
36]. Despite the ecological importance of
P. oceanica, studies on phenological aspects are scarce at the limits of the seagrass distribution range, including the Alboran Sea, where hydrological characteristics of the basin are very different from those in other parts of the Mediterranean Sea due to the Atlantic influence. Within this context, peripheral or edge populations are important for a species’ long-term survival and evolution [
37,
38], as these populations usually present a high genetic differentiation compared to central populations due to reduced population size and genetic drift in fragmented habitats, which results in a lower genetic diversity [
39]. Ecotypes derived from this genetic differentiation respond differently to environmental changes, being able to show even better tolerance to thermal shock than central populations [
40]; however, their response depends on local population traits, as has been observed in some marine macrophytes (e.g.,
Laminaria digitata,
Fucus vesiculosus,
Zostera marina,
P. oceanica) [
40,
41,
42]. This highlights the importance of studying and conserving edge populations of marine macrophytes.
The main objectives of this study are (1) to characterize P. oceanica meadows at its westernmost gradient of distribution in the Mediterranean Sea; (2) to analyze the seasonal and interannual variation of different phenological parameters in the Mediterranean–Atlantic transition region in comparison to other Mediterranean areas; and (3) to evaluate flowering events in different P. oceanica meadows of the northwestern Alboran coasts.
4. Discussion
The distribution of
Posidonia oceanica along the northern Alboran Sea, where its western limit of distribution is located, is not uniform. Here, the most extensive and continuous beds occur off El Ejido (Almería) at the Site of Community Importance, “Fondos Marinos de Punta Entinas-Sabinar” (code ES6110009) [
11]. From here, towards the Strait of Gibraltar (SoG) and the Atlantic Ocean,
P. oceanica meadows are becoming increasingly fragmented, and their extent is considerably reduced. The
P. oceanica meadows studied along the northwestern Alboran Sea display a decreasing trend of patch size, bathymetric range and the number of leaves per shoot on moving toward the SoG, as well as an increasing trend of shoot density and leaf height. Similarly, the maximum depth reached by
P. oceanica meadows decreases westwards from Punta Entinas-Sabinar (ca. 15 m depth) [
57] to Chullera (1–1.5 m depth). The influence of surface Atlantic water entering eastwards through the SoG, together with the characteristic oceanographic and hydrological conditions of the northern Alboran Sea (e.g., higher turbidity, stronger wave action) [
5,
44,
58], might represent a developmental limiting factor for
P. oceanica in this part of the Mediterranean. In this respect, other studies have documented the presence of other habitat types at unusually shallow bottoms in the northwestern Alboran Sea, such as coralligenous assemblages found at a depth of 15 m, which is usually characteristic of the circalittoral biocenosis [
59,
60]. This indicates lower water transparency in the northwestern Alboran Sea compared to other Mediterranean areas, as has also been detected in the adjacent Gulf of Cádiz. Moreover, the analyzed
P. oceanica meadows are located in one of the highest biological productivity areas within the Mediterranean [
61,
62], which may also induce low water transparency. This was reflected in the low irradiance values obtained at a depth of 4–5 m at the Calahonda SAC, which are similar to those found at a depth of 10 m in the eastern Adriatic Sea [
50].
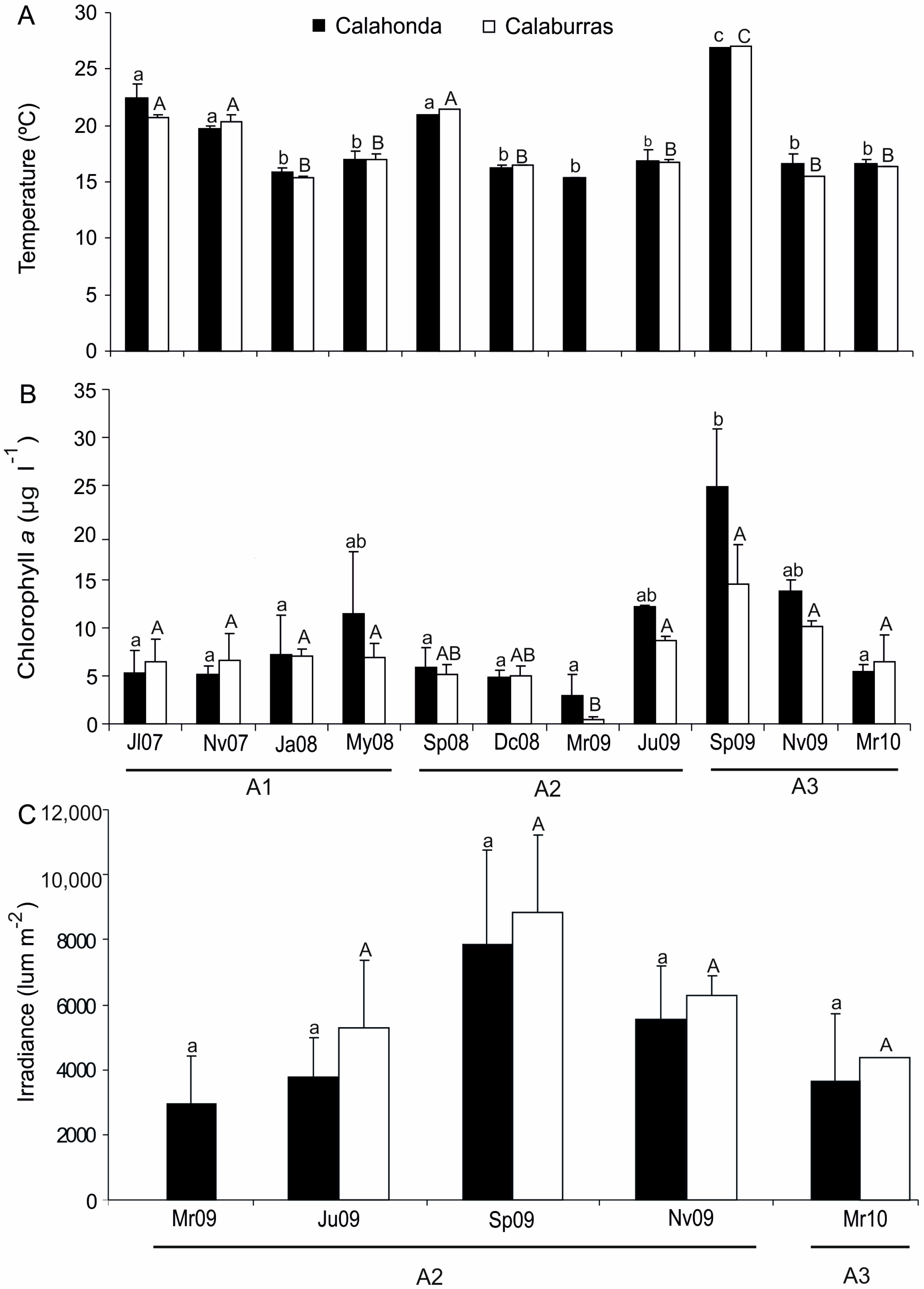
Solar irradiance and seawater temperature are key factors modulating the abundance and productivity of seagrasses [
8,
12,
63]. Indeed, light and surface seawater temperature explained a high percentage of variability of leaf number and shoot density of
P. oceanica at the Calahonda SAC throughout the year. On the other hand, a narrower bathymetric distribution (ranging between 1–5 m depth) exposes the meadows to continuous wave action and turbulence, especially in the areas close to the SoG, where patches were very close to the surf zone (1–1.5 m depth). Regarding this aspect, shallow patchy meadows of different seagrasses have been reported to be affected by such extreme conditions (e.g., low water transparency and strong wave action), including
P. oceanica meadows at its northern distributional boundary in the Adriatic Sea [
64,
65,
66,
67]. Within this context, studies carried out on
P. oceanica, and other seagrasses show a negative relationship between seagrass biomass, leaf height, epiphyte biomass, leaf width and shoot area with wave energy [
68,
69]. Moreover, high-energy environments support
P. oceanica meadows landscapes with higher patch numbers, lower coverage, more complex patch shapes and lower intra-patch architecture than sites exposed to lower wave action [
69].
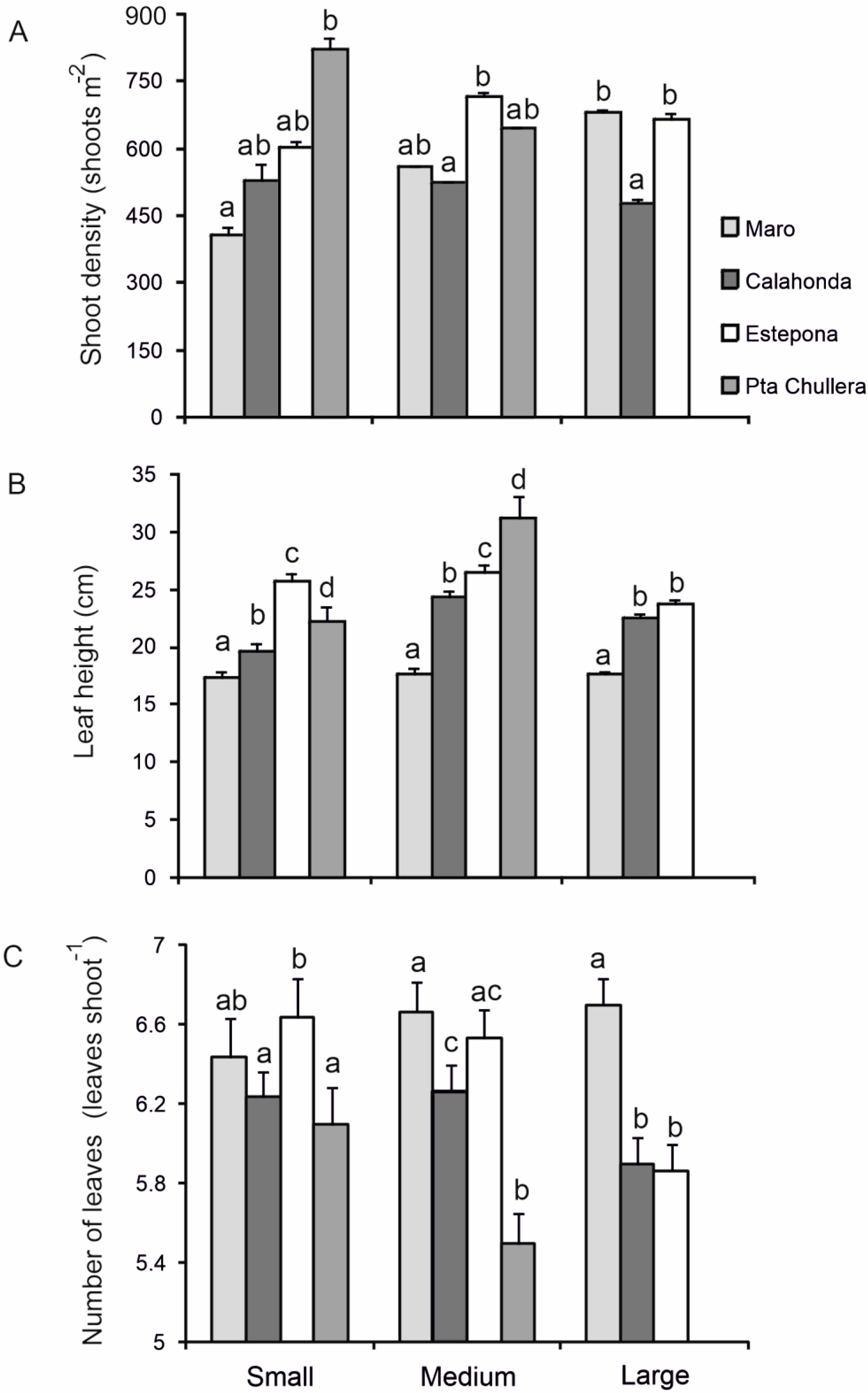
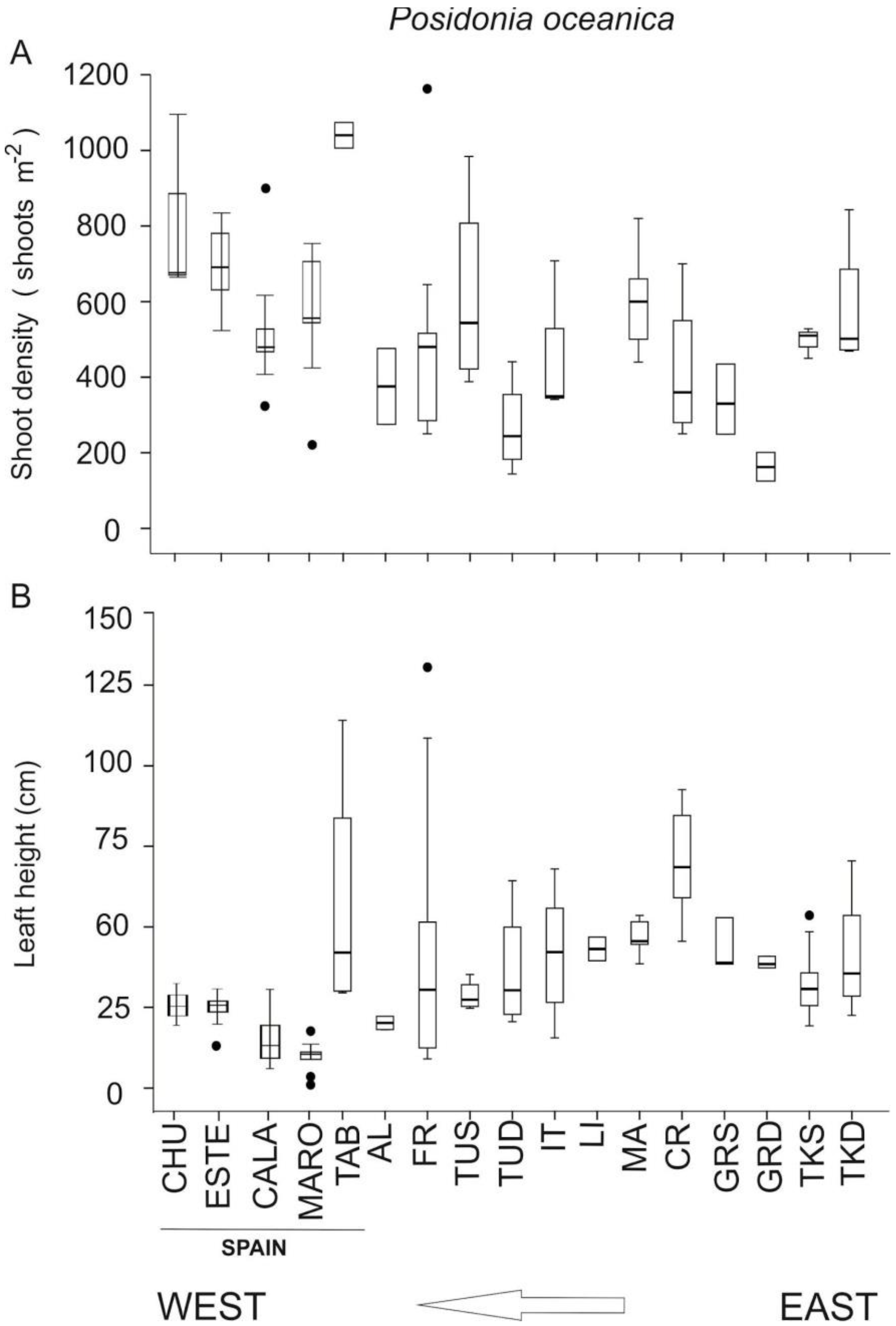
Shoot densities of the studied
P. oceanica meadows were generally higher than those located in other Mediterranean areas at similar depths, including the north Adriatic Sea [
66]) and the northeastern Levantine Sea (eastern distributional boundary [
12]), but similar to those reported in meadows in Tunisia growing on a rocky substratum, as the ones studied here [
70] (see
Figure 6). Regarding leaf height, values recorded in the present study were lower than those documented in other Mediterranean areas (see
Figure 6). Regarding this, Marbà et al. [
8] reported
P. oceanica meadows with high shoot densities and low leaf height values in other northern Alboran Sea locations [
8]. Several physical factors acting together seem to highly influence the westernmost
P. oceanica meadows, such as the shallow location and the rocky nature of the substratum, together with higher hydrodynamic exposure due to wave action. Furthermore, they may represent key drivers structuring ecological features and phenological parameters of these meadows along the northern Alboran Sea [
11,
68,
69], being particularly evident at the westernmost meadows of Estepona SAC and especially of Chullera. This could represent a stress reduction strategy that involves increasing rhizome growth for anchorage as well as self-shading effects [
35,
71,
72], as suggested by the negative correlations observed at Calahonda SAC between shoot densities, leaf height and the number of leaves per shoot.
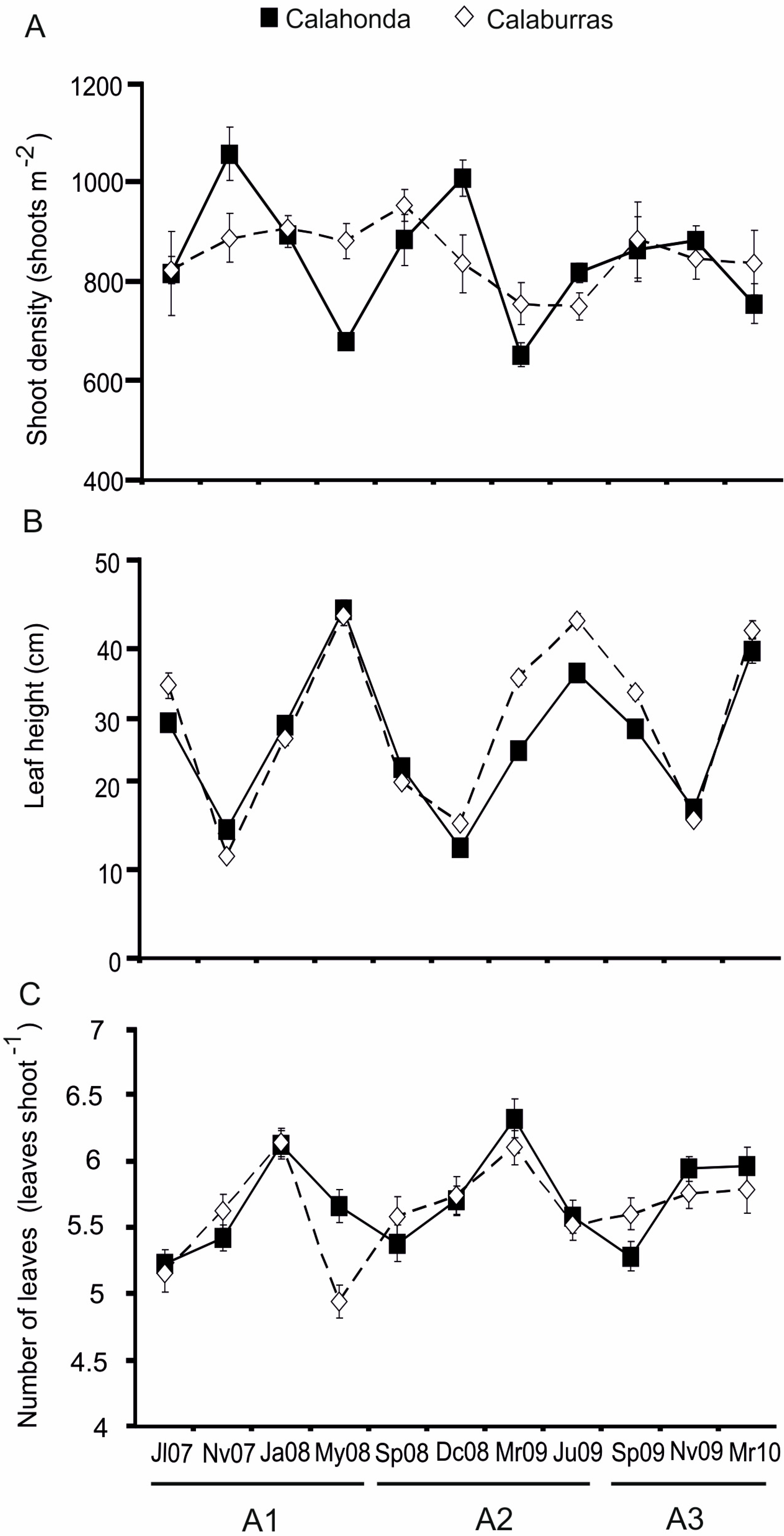
The temporal trend observed for leaf height and number of leaves per shoot is similar to that reported for other
P. oceanica meadows of both European and African coasts [
30,
31,
32,
36,
73,
74] Regarding shoot density and despite being a more stable descriptor, higher values were observed in cold months in Calahonda SAC. Moreover, an unusual seasonal trend was observed during the first two years, similar to that previously documented in other meadows, some of which were located close to the northern and eastern distribution edges of
P. oceanica in Tunisia and Turkey [
36,
71,
75]. Nevertheless, shoot density and number of leaves per shoot displayed lower intra-annual fluctuations when compared to other
P. oceanica meadows (see
Table 6), probably linked to the particular environmental characteristics of the northwestern Alboran Sea, including lower intra-annual temperature changes (15.3–22.5 °C) in comparison to other Mediterranean areas (e.g., Adriatic Sea: 9–24 °C; Tunisian coasts: 14–31 °C; southern Italian coasts: 11.8–24.8 °C [
33,
73,
76]), which may highly influence these meadows due to their shallow location. On the other hand, maximum leaf height displayed high differences amongst years, as observed previously [
8,
35,
36]. Interannual studies of phenological parameters are scarce, but all of them have indicated some interannual variability, highlighting the need for performing long-term studies of meadow population characters and phenological parameters [
8,
35,
36].
The inflorescences followed by fruit development observed at Calahonda SAC would indicate that sexual reproduction of
P. oceanica may potentially occur in meadows at their western limit of geographical distribution [
80]. Flowering of
P. oceanica is highly variable within and between meadows, with differences even at small spatial scales of a few meters [
81]. Despite this, the flowering and recruitment of this Mediterranean seagrass seem to be more frequent than previously expected [
82,
83]. The available information seems to indicate that flowering represents an irregular event mainly related to high seawater temperature [
84,
85]. In fact, three flowering events were registered in the meadows at Calahonda SAC between 2006 and 2015, with the highest flowering episode coinciding with the warmest seawater temperature values ever recorded for the area at that time. Regarding this, it is remarkable that the high flowering frequency observed in 2015, with similar or even higher values than those reported in meadows of the central and eastern Mediterranean basin [
75,
86,
87]. Nevertheless, no flowering events were detected in Maro MPA, Estepona SAC, or Chullera during that year. This is of interest as Calahonda SAC meadows are located close to the westernmost distributional limit of
P. oceanica, which would indicate (1) that these meadows present an adequate health status for sexual reproduction; or (2) a survival mechanism in response to natural or anthropogenic disturbances [
85,
88]. Furthermore, the intensity of the flowering and the absence of significant differences amongst patches of different sizes in relation to inflorescence density may be related to the latter notion. Fruit production and seed germination still need to be monitored in these
P. oceanica meadows in long-term studies in order to evaluate the extent of sexual reproduction in the formation of new patches. This is of importance to evaluate the survival capacity of
P. oceanica, as edge populations could play an important role in the long-term survival of this seagrass; in fact, a recent study shows that thermal edge populations of
P. oceanica have a greater resilience against thermal stress than central populations [
42].
Disentangling natural, anthropogenic impacts on different organisms and habitats is still very difficult to achieve. In spite of being exposed to high anthropogenic pressure (intense coastal development and high human population), most of the studied
P. oceanica meadows were in a good conservation status as reflected in the BiPo index, with EQR values similar to those of other meadows located in areas that are more pristine and where the human population density is low [
55]. This could be related to the peculiar characteristics of the
P. oceanica meadows at our study sites compared to other sites in the Mediterranean Sea (see
Figure 6). Nevertheless, the Maro MPA meadow showed evidence of decline, which could be partly related to large flooding linked to a torrential rainfall (200 mm per m
2) recorded in the area in September 2007. This had serious consequences for the
P. oceanica meadow located here as several streams flow into the small cove of “Molino de Papel”. Flooding of these watercourses led to the transport of sand and mud, plant debris, and waste of different kinds into the sea, which were deposited in a large part of the meadow. This caused the burial and subsequent death of approximately half the existing meadow [
89] (see
Figure 2D). The Calahonda SAC meadows also show a high presence of old and dead rhizomes, with no leaves and the matte covered by macroalgae, which may indicate that
P. oceanica has suffered a regression and it was more abundant in the past (see
Figure 2C).

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}