
Makeup Water Addition Can Affect the Growth of Scenedesmus dimorphus in Photobioreactors

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strain
2.2. Algal Cultivation with Makeup Water and Growth Medium
- Add makeup water every day (1D). Autoclaved deionized water was added to compensate for water loss and to maintain a 2000 mL algal culture in each bottle. Before water addition, several parameters were measured, including temperature, pH, algal biomass, OD, and cell counts. The tests consumed ~26 mL of algal culture. This volume was also included in the water loss calculation. The entire experiment lasted 14 days.
- Add makeup water every four days (4D). The experiment started with another 1200 mL healthy algal culture that was independent of the previous batch. Again, deionized water was added to maintain a 2000 mL algal culture in each bottle. Temperature, pH, algal biomass, OD, and cell counts were measured every day, and on the day when water was added, the measurement was performed right before water addition. The entire experiment lasted 21 days.
2.3. Analytical Methods
2.4. Data Analysis
3. Results and Discussion
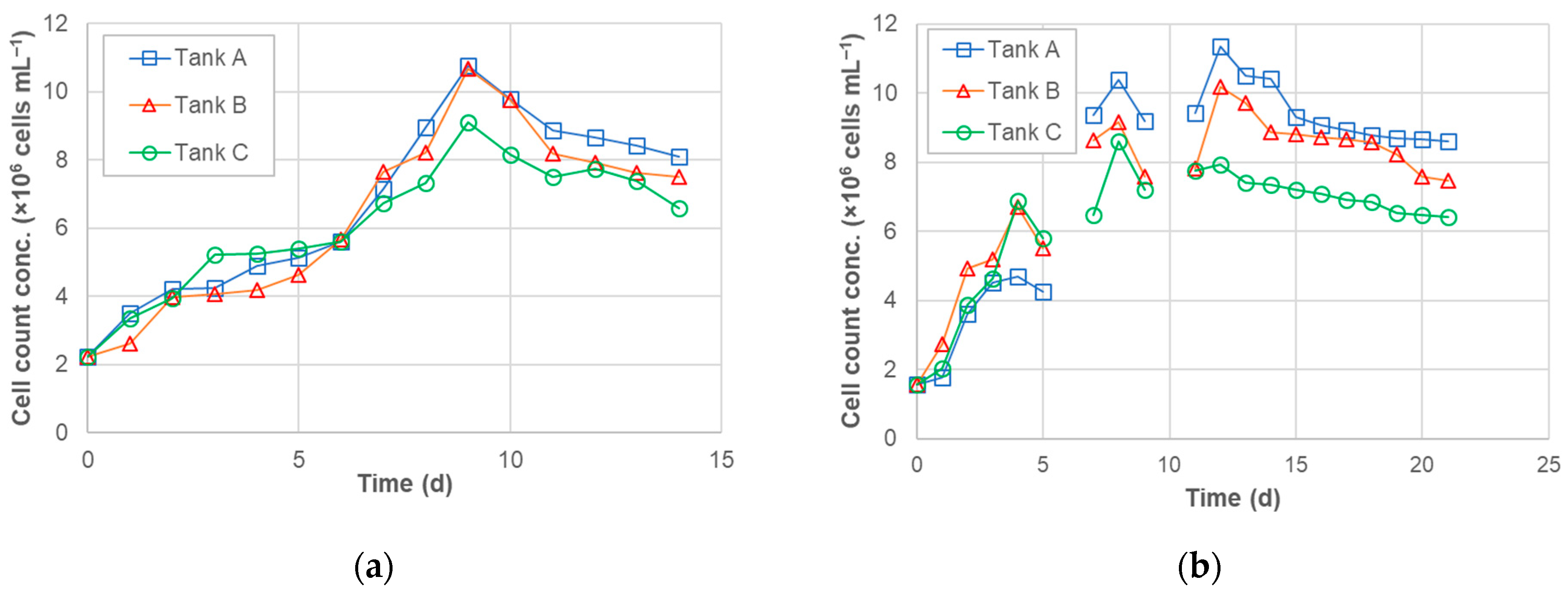
3.1. Algal Cell Counts
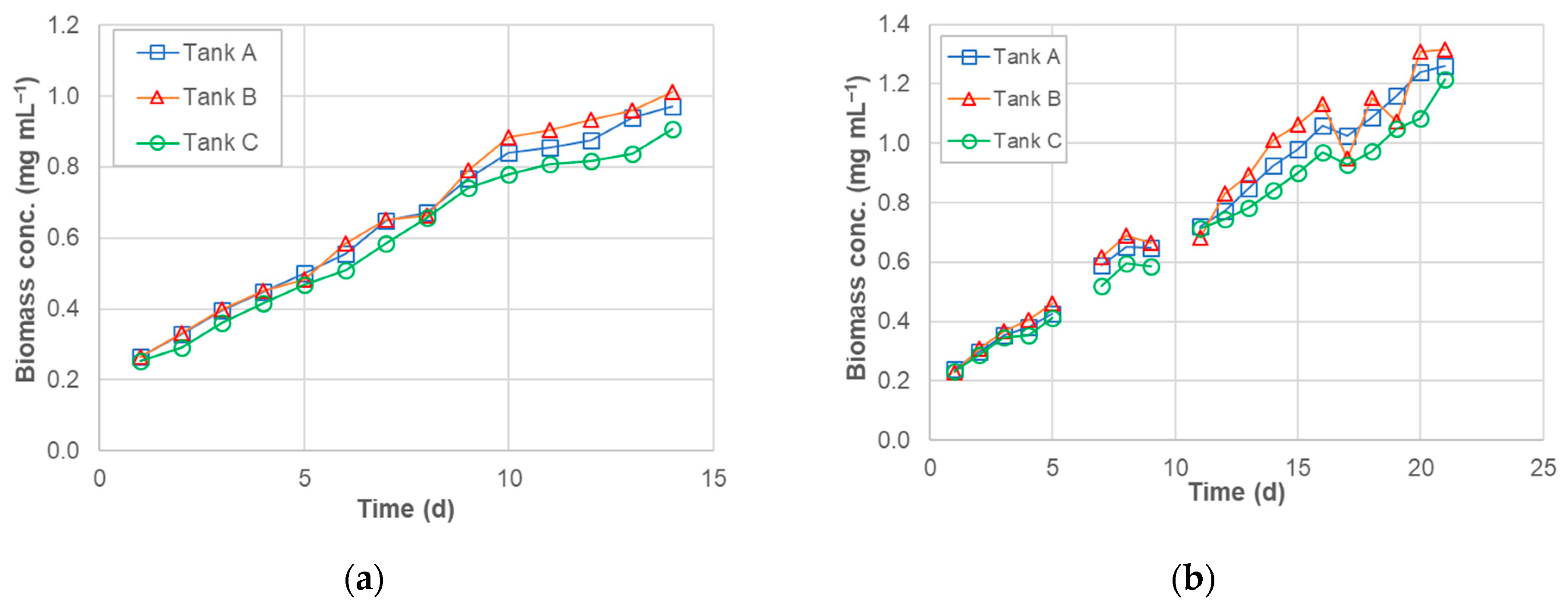
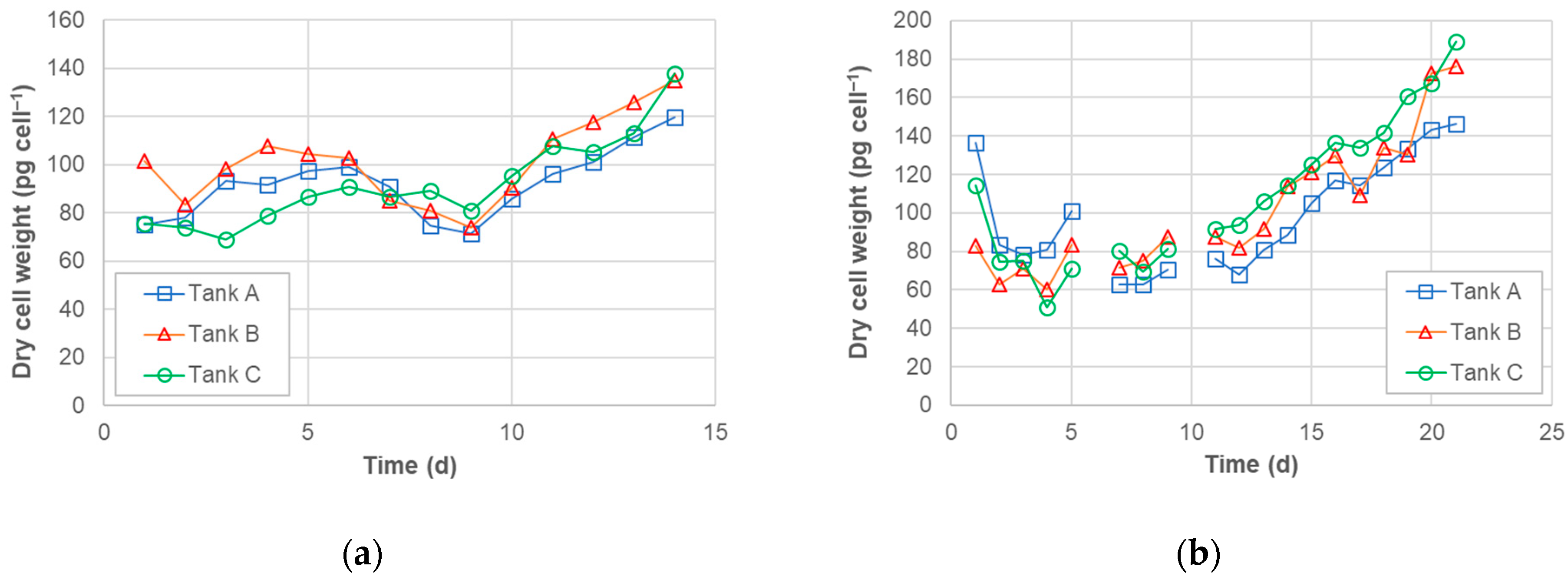
3.2. Dry Algal Biomass (DAB)
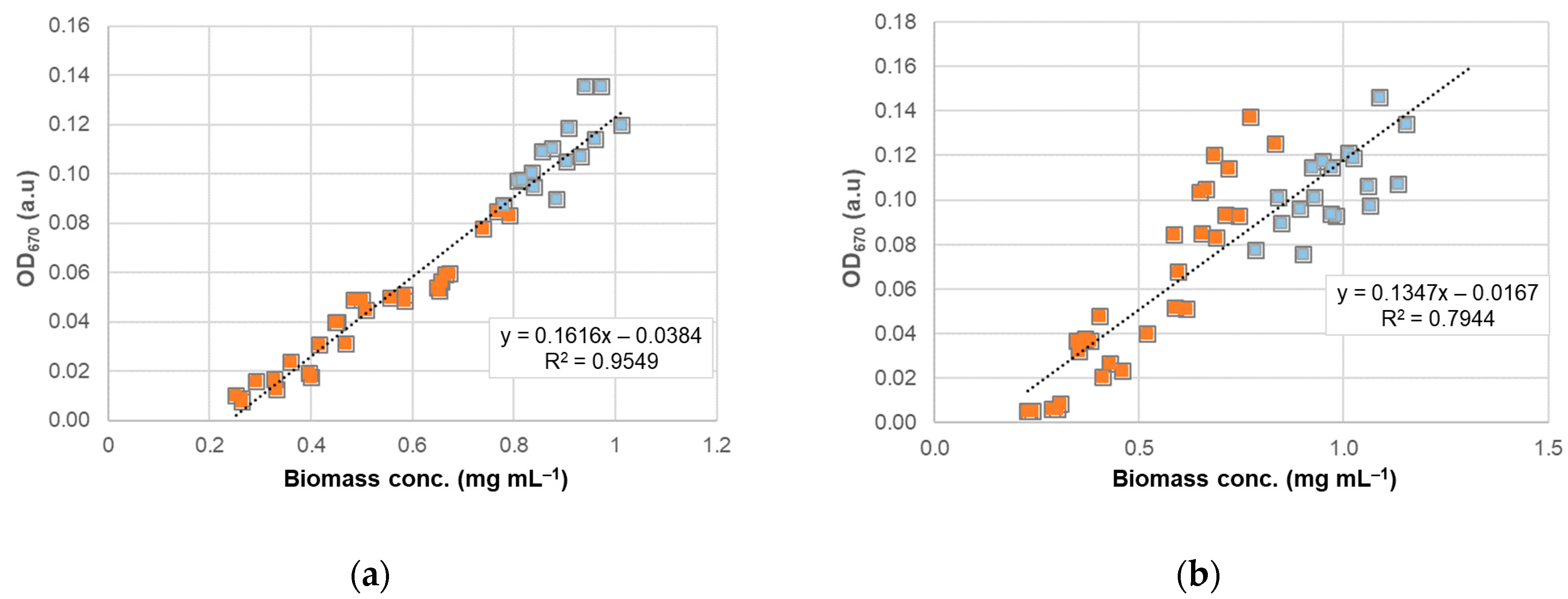
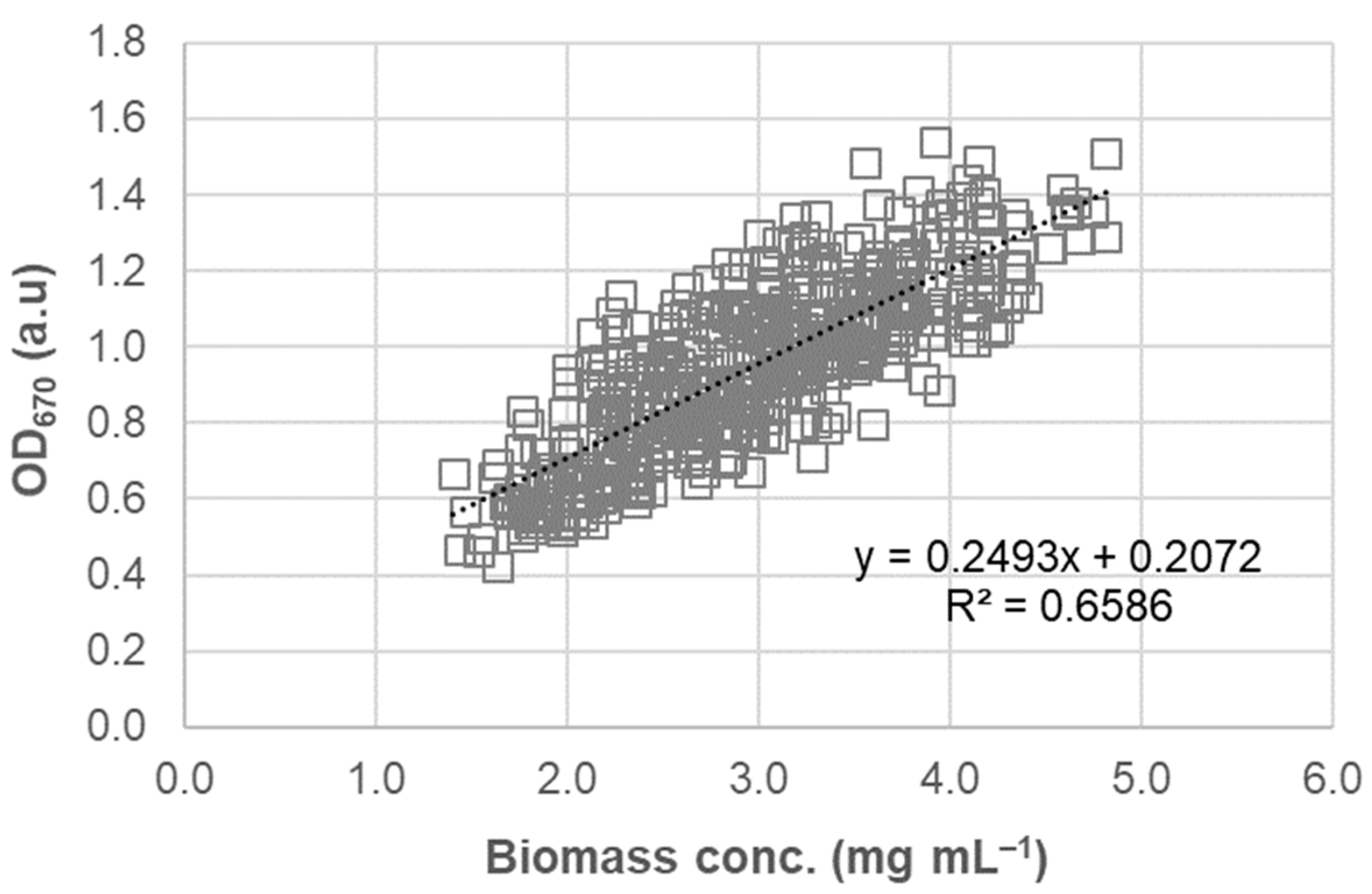
3.3. OD670 as a Surrogate for Algal Biomass
3.4. Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Mascarelli, A.L. Algae: Fuel of the future? Environ. Sci. Technol. 2009, 43, 7160–7161. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Kuo, E.W.; Nagarajan, D.; Ho, S.H.; Dong, C.D.; Lee, D.J.; Chang, J.S. Cultivating Chlorella sorokiniana AK-1 with swine wastewater for simultaneous wastewater treatment and algal biomass production. Bioresour. Technol. 2020, 302, 122814. [Google Scholar] [CrossRef] [PubMed]
- Uguz, S.; Anderson, G.; Yang, X.; Simsek, E.; Osabutey, A. Cultivation of Scenedesmus dimorphus with air contaminants from a pig confinement building. J. Environ. Manag. 2022, 314, 115129. [Google Scholar] [CrossRef]
- Sirakov, M.; Palmieri, M.; Iovinella, M.; Davis, S.J.; Petriccione, M.; di Cicco, M.R.; De Stefano, M.; Ciniglia, C. Cyanidiophyceae (rhodophyta) tolerance to precious metals: Metabolic response to palladium and gold. Plants 2021, 10, 2367. [Google Scholar] [CrossRef]
- Zhao, B.; Su, Y. Process effect of microalgal-carbon dioxide fixation and biomass production: A review. Renew. Sustain. Energy Rev. 2014, 31, 121–132. [Google Scholar] [CrossRef]
- Murphy, C.F.; Allen, D.T. Energy-water nexus for mass cultivation of algae. Environ. Sci. Technol. 2011, 45, 5861–5868. [Google Scholar] [CrossRef]
- Moglia, M.; Cook, S.; Tapsuwan, S. Promoting water conservation: Where to from here? Water 2018, 10, 1510. [Google Scholar] [CrossRef]
- Feng, P.Z.; Zhu, L.D.; Qin, X.X.; Li, Z.H. Water footprint of biodiesel production from microalgae cultivated in photobioreactors. J. Environ. Eng. 2016, 142, 04016067. [Google Scholar] [CrossRef]
- Clarens, A.F.; Resurreccion, E.P.; White, M.A.; Colosi, L.M. Environmental life cycle comparison of algae to other bioenergy feedstocks. Environ. Sci. Technol. 2010, 44, 1813–1819. [Google Scholar] [CrossRef]
- Yang, J.; Xu, M.; Zhang, X.; Hu, Q.; Sommerfeld, M.; Chen, Y. Life-cycle analysis on biodiesel production from microalgae: Water footprint and nutrients balance. Bioresour. Technol. 2011, 102, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, D.; Greene, J.M.; McGowen, J.; Quinn, J.C. Geographical assessment of open pond algal productivity and evaporation losses across the United States. Algal Res. 2021, 60, 102483. [Google Scholar] [CrossRef]
- Martins, A.A.; Marques, F.; Cameira, M.; Santos, E.; Badenes, S.; Costa, L.; Vieira, V.V.; Caetano, N.S.; Mata, T.M. Water footprint of microalgae cultivation in photobioreactor. Energy Procedia 2018, 153, 426–431. [Google Scholar] [CrossRef]
- Sander, K.; Murthy, G.S. Life cycle analysis of algae biodiesel. Int. J. Life Cycle Assess. 2010, 15, 704–714. [Google Scholar] [CrossRef]
- Kang, J.; Wang, T.; Xin, H.; Wen, Z. A laboratory study of microalgae-based ammonia gas mitigation with potential application for improving air quality in animal production operations. J. Air Waste Manag. Assoc. 2014, 64, 330–339. [Google Scholar] [CrossRef]
- Xu, X.; Shen, Y.; Chen, J. Cultivation of Scenedesmus dimorphus for C/N/P removal and lipid production. Electron. J. Biotechnol. 2015, 18, 46–50. [Google Scholar] [CrossRef]
- Fábregas, J.; Domínguez, A.; Regueiro, M.; Maseda, A.; Otero, A. Optimization of culture medium for the continuous cultivation of the microalga Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 2000, 53, 530–535. [Google Scholar] [CrossRef]
- Hu, W. Dry Weight and Cell Density of Individual Algal and Cyanobacterial Cells for Algae Research and Development. Master’s Thesis, University of Missouri-Columbia, Columbia, MO, USA, 2014. [Google Scholar]
- Menegazzo, M.L.; Ribeiro, D.M.; De Oliveira, N.N.; Marques, O.G.B.; Fonseca, G.G. Evaluation of Chlorella sorokiniana biomass recovery by using different chemical-based flocculants. J. Appl. Biotechnol. Rep. 2022, 91, 477–483. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Velichkova, K.; Sirakov, I.; Georgiev, G. Cultivation of Scenedesmus dimorphus strain for biofuel production. Agric. Sci. Technol. 2013, 5, 181–185. [Google Scholar]
- Varsharani, H.; Geeta, G. Isolation of microalgae with limitation on fatty acid and lipid content of marine microalgae. Biodiesel Product. Prospects 2011, 24, 585–588. [Google Scholar]
- Padovan, A. Isolation and Culture of Five Species of Freshwater Algae from the Alligator Rivers Region, Northern Territory; Australian Government Publishing Service: Canberra, Australia, 1992. Available online: https://www.awe.gov.au/science-research/supervising-scientist/publications/technical-memoranda/isolation-and-culture-five-species-freshwater-algae-alligator-rivers-region (accessed on 9 March 2022).
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of microalgae cultivation in photobioreactor, open raceway pond, and a two-stage hybrid system. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef]
- Bordoloi, N.; Narzari, R.; Sut, D.; Saikia, R.; Chutia, R.S.; Kataki, R. Characterization of bio-oil and its sub-fractions from pyrolysis of Scenedesmus dimorphus. Renew. Energy 2016, 98, 245–253. [Google Scholar] [CrossRef]
- Latsos, C.; van Houcke, J.; Timmermans, K.R. The effect of nitrogen starvation on biomass yield and biochemical constituents of Rhodomonas sp. Front. Mar. Sci. 2020, 7, 563333. [Google Scholar] [CrossRef]
- Rugnini, L.; Rossi, C.; Antonaroli, S.; Rakaj, A.; Bruno, L. The influence of light and nutrient starvation on morphology, biomass and lipid content in seven strains of green microalgae as a source of biodiesel. Microorganisms 2020, 8, 1254. [Google Scholar] [CrossRef]
- Perdana, B.A.; Dharma, A.; Zakaria, I.J.; Syafrizayanti, S. Freshwater pond microalgae for biofuel: Strain isolation, identification, cultivation and fatty acid content. Biodiversitas 2021, 22, 505–511. [Google Scholar] [CrossRef]
- Ferreira, V.S.; Pinto, R.F.; Sant’Anna, C. Low light intensity and nitrogen starvation modulate the chlorophyll content of Scenedesmus dimorphus. J. Appl. Microbiol. 2016, 120, 661–670. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, W.; Wang, J.; Chen, Y.; Shen, S.; Liu, T. Utilization of simulated flue gas for cultivation of Scenedesmus dimorphus. Bioresour. Technol. 2013, 128, 359–364. [Google Scholar] [CrossRef]
- Cicci, A.; Stoller, M.; Bravi, M. Microalgal biomass production by using ultra-and nanofiltration membrane fractions of olive mill wastewater. Water Res. 2013, 47, 4710–4718. [Google Scholar] [CrossRef]
- Bohutskyi, P.; Kligerman, D.C.; Byers, N.; Nasr, L.K.; Cua, C.; Chow, S.; Su, C.; Tang, Y.; Betenbaugh, M.J.; Bouwer, E.J. Effects of inoculum size, light intensity, and dose of anaerobic digestion centrate on growth and productivity of Chlorella and Scenedesmus microalgae and their poly-culture in primary and secondary wastewater. Algal Res. 2016, 19, 278–290. [Google Scholar] [CrossRef]
- Tran, H.D.; Ong, B.N.; Ngo, V.T.; Tran, D.L.; Nguyen, T.C.; Tran-Thi, B.H.; Do, T.T.; Nguyen, T.M.L.; Nguyen, X.H.; Melkonian, M. New angled twin–layer porous substrate photobioreactors for cultivation of Nannochloropsis oculata. Protist 2022, 173, 125914. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osabutey, A.; Haleem, N.; Uguz, S.; Albert, K.L.; Anderson, G.A.; Min, K.; Yang, X. Makeup Water Addition Can Affect the Growth of Scenedesmus dimorphus in Photobioreactors. AgriEngineering 2023, 5, 982-991. https://doi.org/10.3390/agriengineering5020061
Osabutey A, Haleem N, Uguz S, Albert KL, Anderson GA, Min K, Yang X. Makeup Water Addition Can Affect the Growth of Scenedesmus dimorphus in Photobioreactors. AgriEngineering. 2023; 5(2):982-991. https://doi.org/10.3390/agriengineering5020061
Chicago/Turabian StyleOsabutey, Augustina, Noor Haleem, Seyit Uguz, Karlee L. Albert, Gary A. Anderson, Kyungnan Min, and Xufei Yang. 2023. "Makeup Water Addition Can Affect the Growth of Scenedesmus dimorphus in Photobioreactors" AgriEngineering 5, no. 2: 982-991. https://doi.org/10.3390/agriengineering5020061