
Cyanobacteria Arthospira platensis as an Effective Tool for Gadolinium Removal from Wastewater
 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Object of Study
2.2. Biosorption Experiment
2.3. Bioaccumulation Experiment
2.4. Biomass Productivity and Biochemical Tests
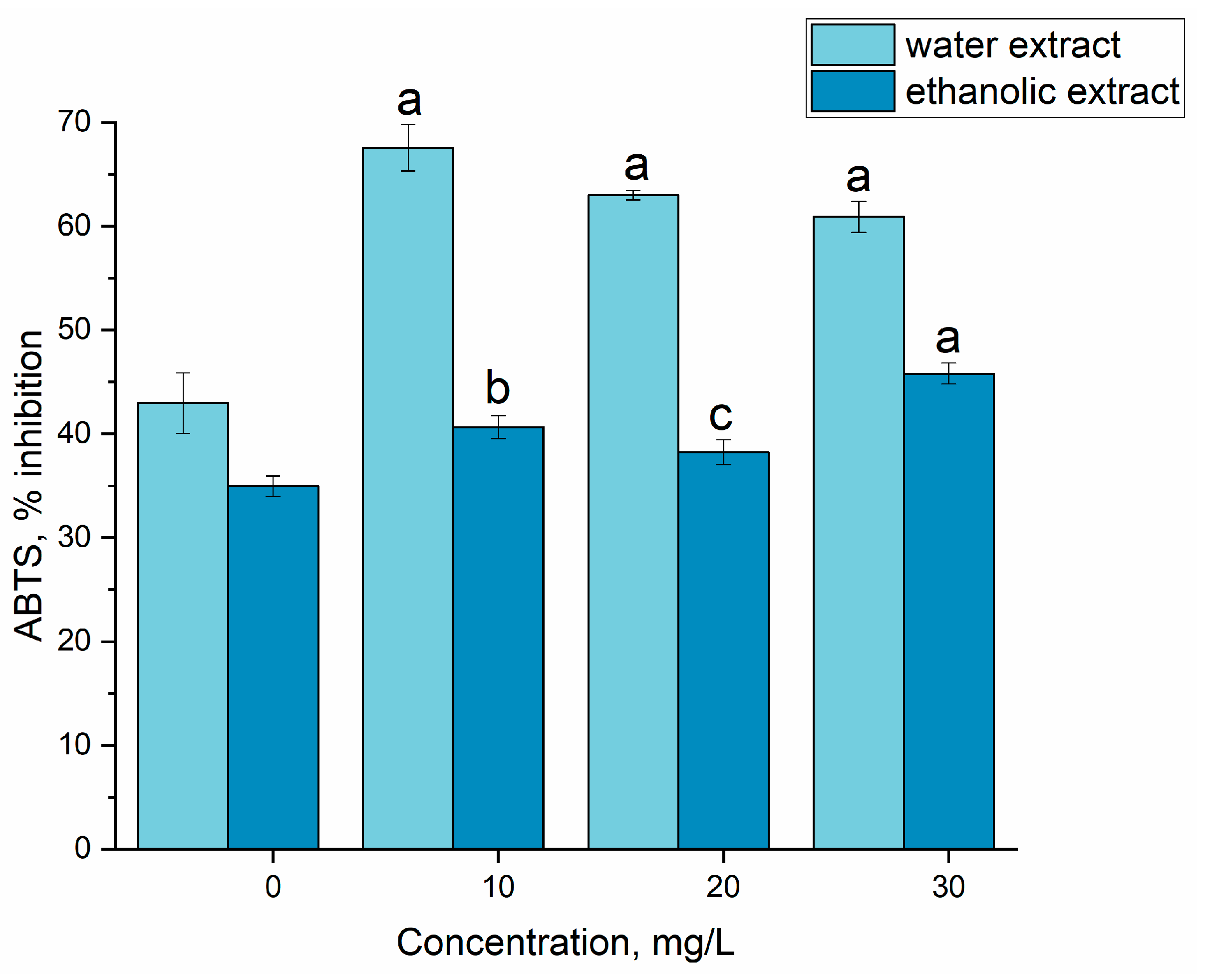
2.5. Antioxidant Activity
2.6. Statistical Analysis
3. Results and Discussion
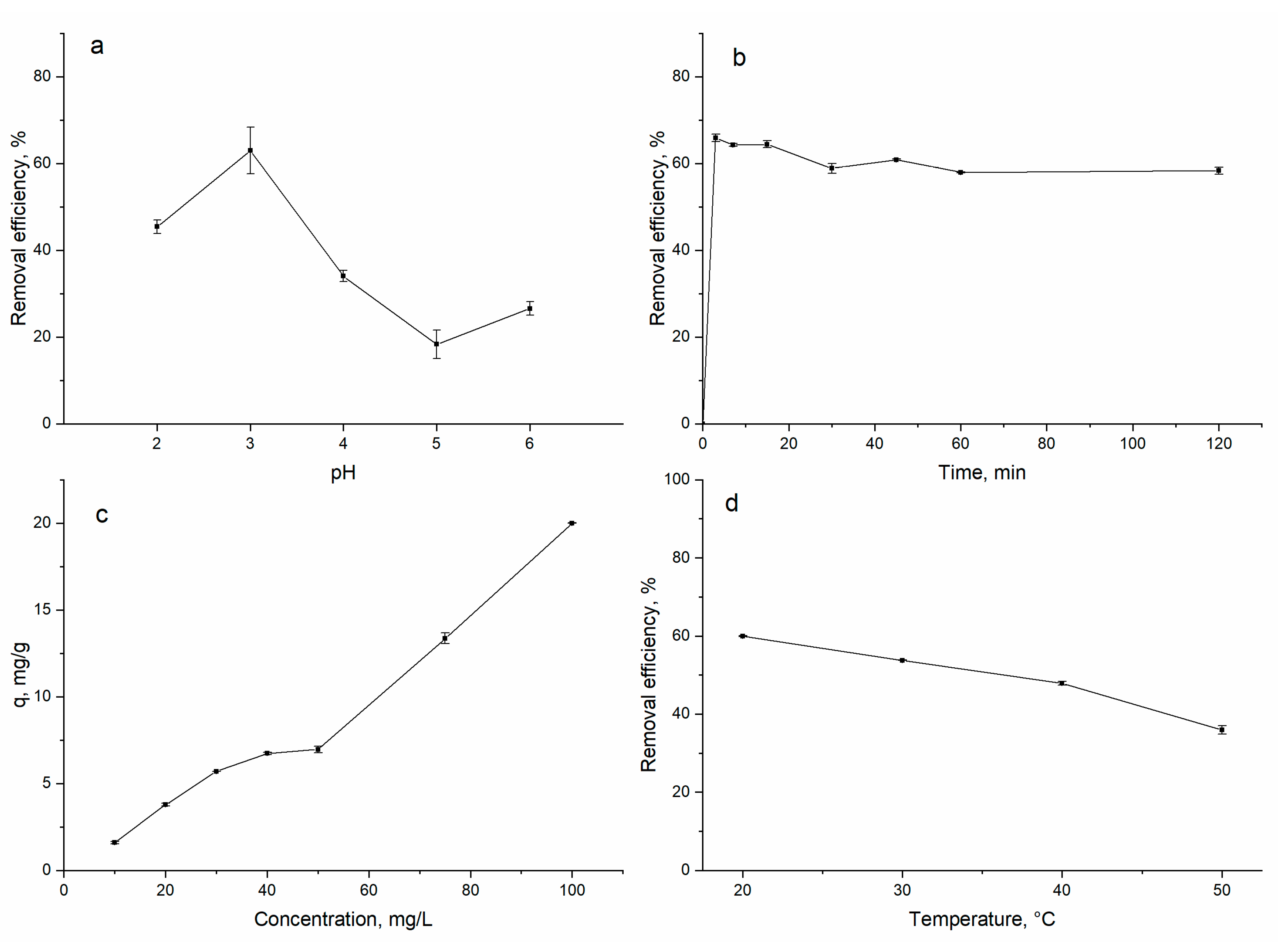
3.1. Biosorption of Gadolinium on A. platensis
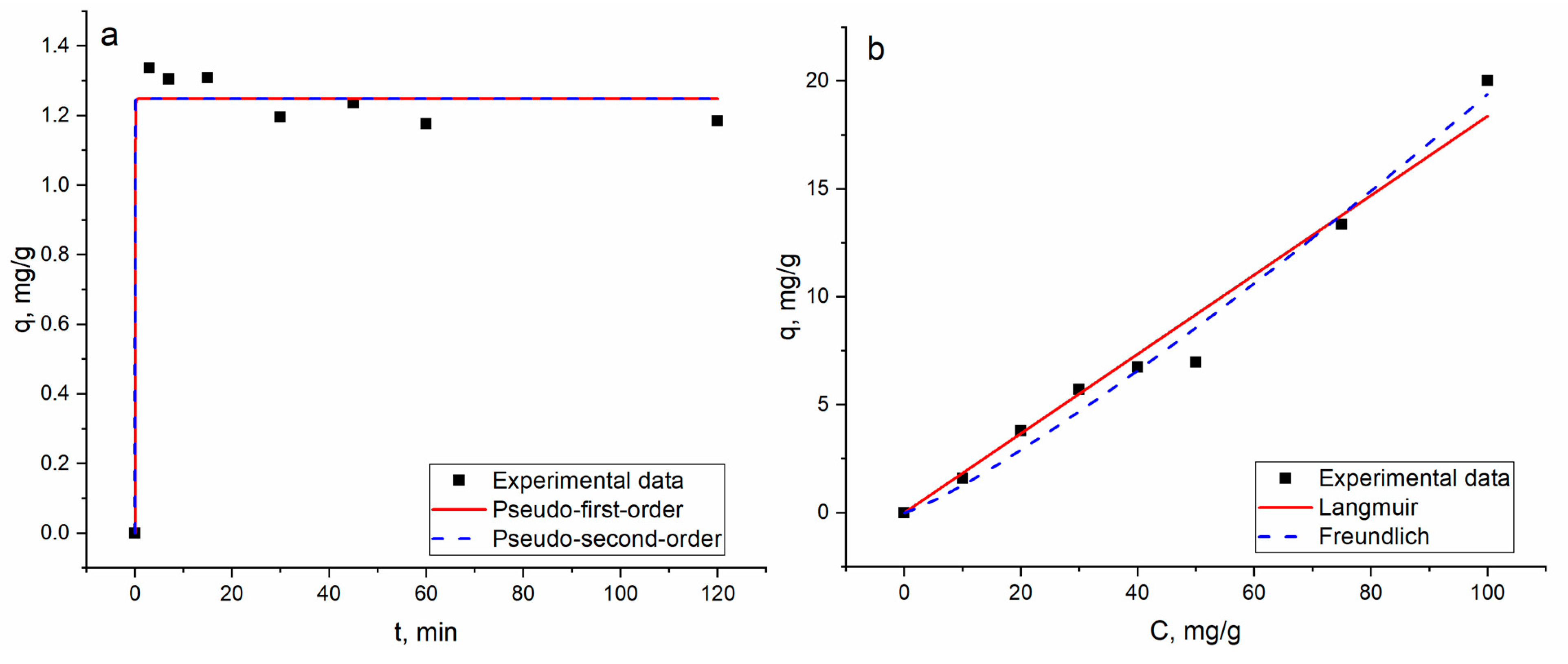
3.2. Equilibrium, Kinetics and Thermodynamic Studies
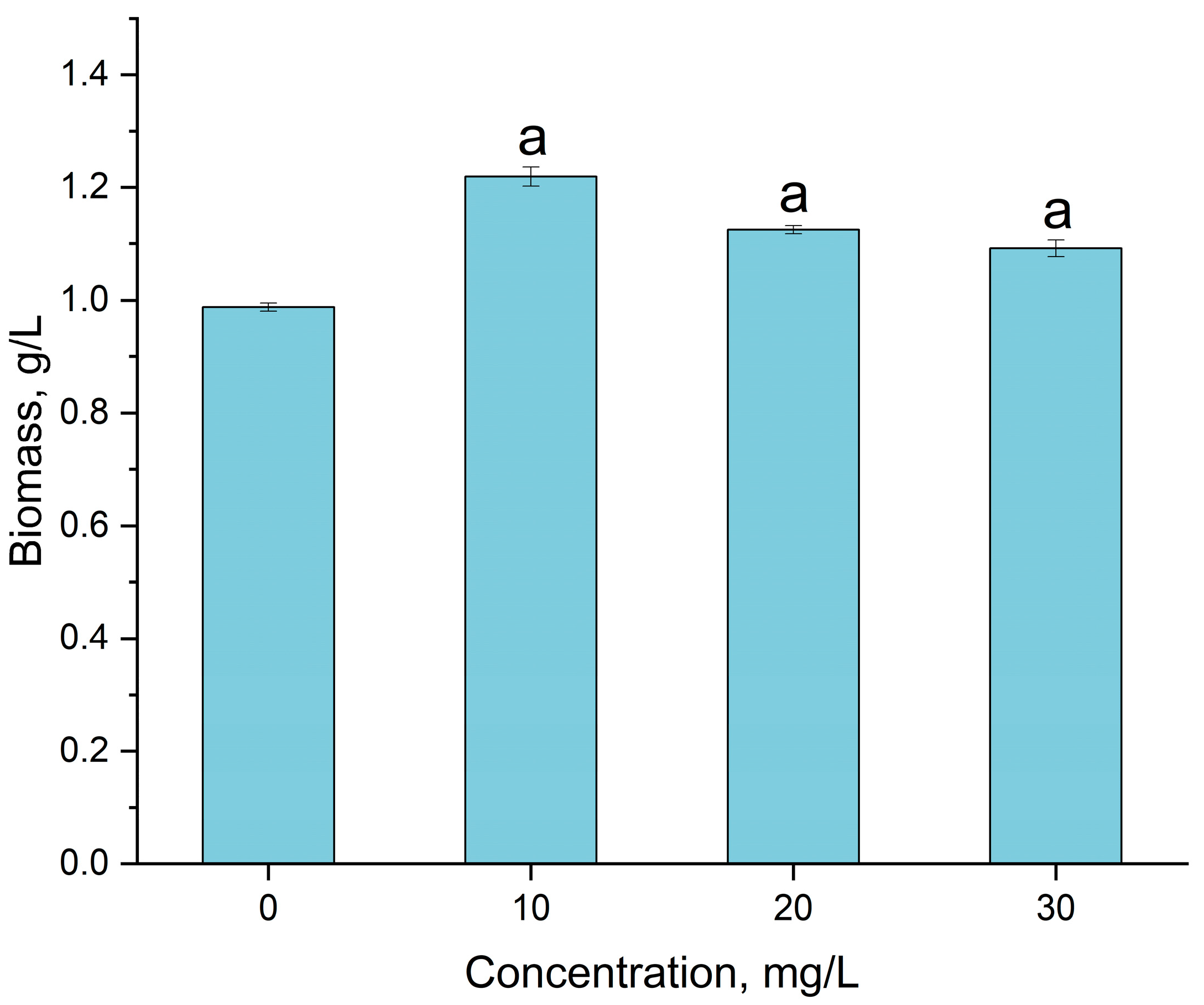
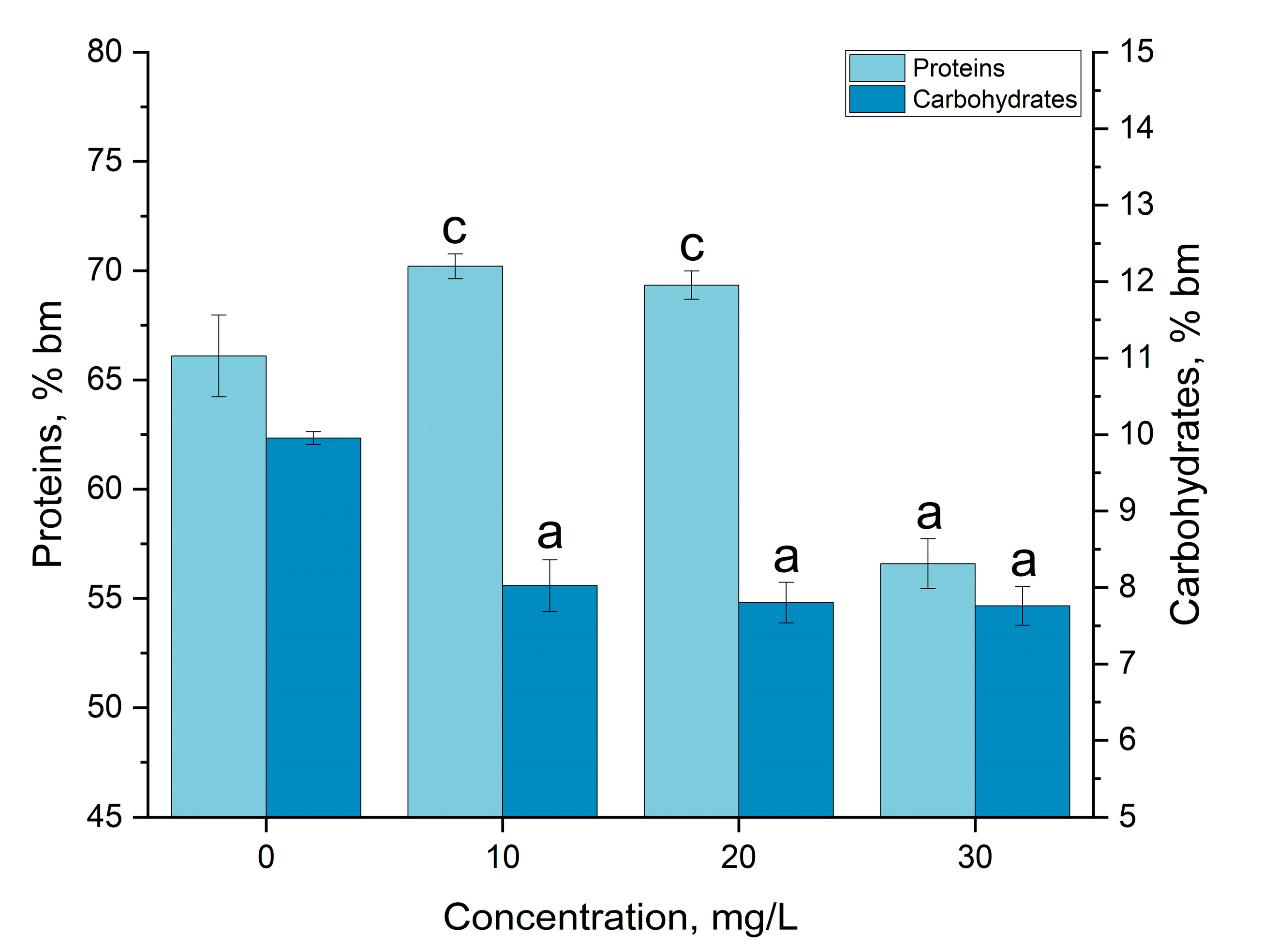
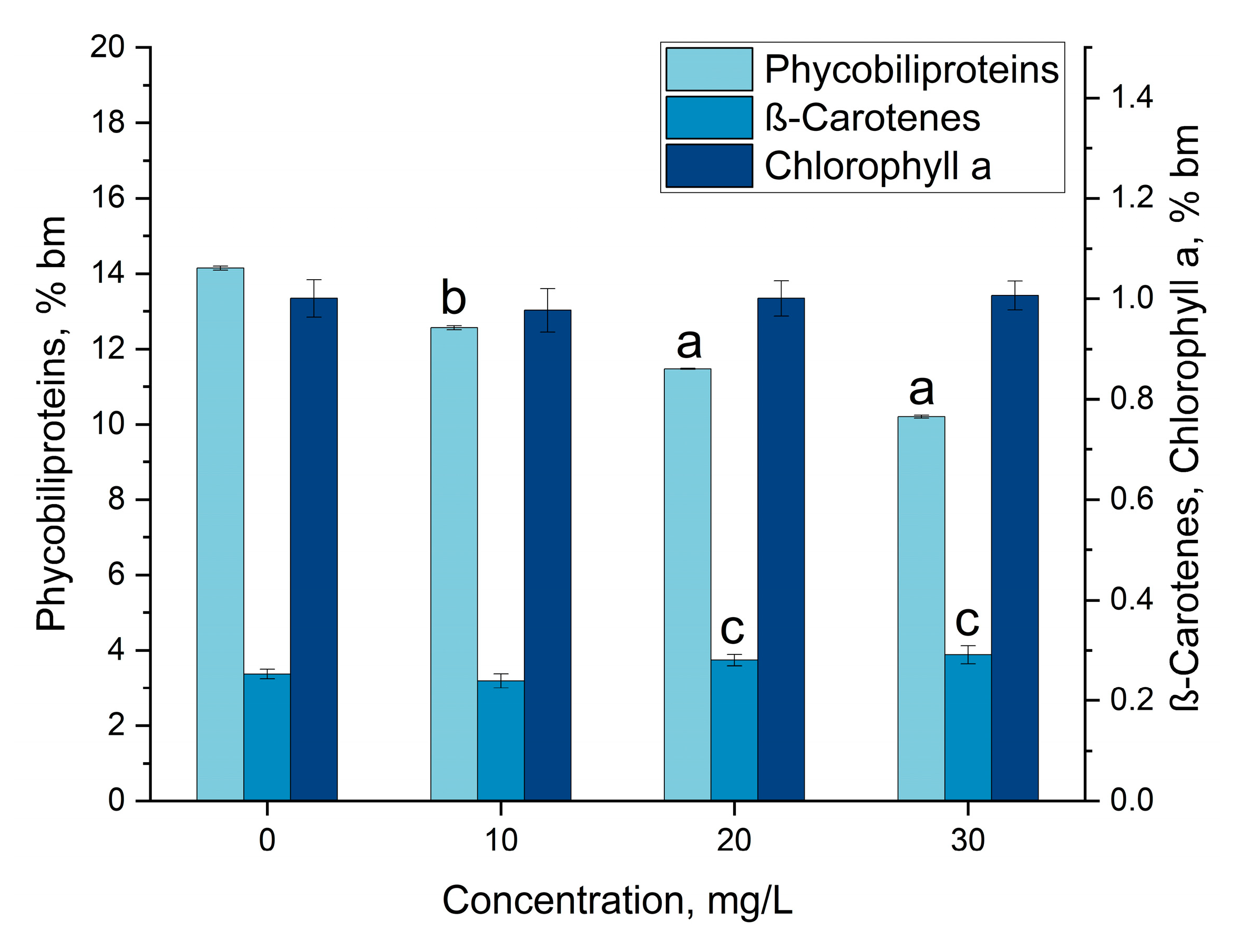
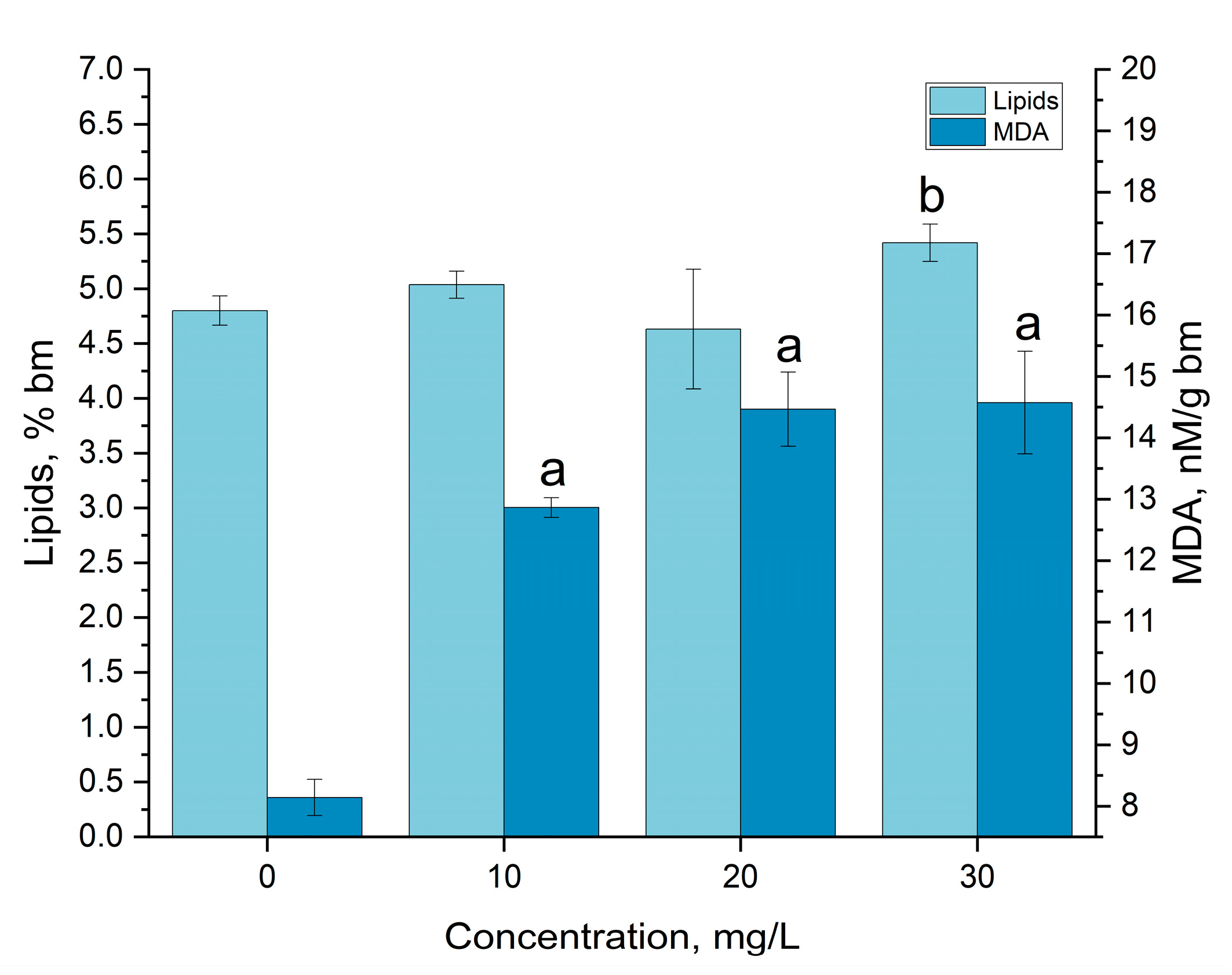
3.3. Gadolinium Bioaccumulation Using A. platensis and Assessment of Biochemical Changes in Biomass
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamed, M.M.; Rizk, S.E.; Nayl, A.A. Adsorption Kinetics and Modeling of Gadolinium and Cobalt Ions Sorption by an Ion-Exchange Resin. Part. Sci. Technol. 2016, 34, 716–724. [Google Scholar] [CrossRef]
- Maksin, D.; Vukčević, M.; Đurkić, T.; Stanišić, I.; Bakić, T.; Radomirović, M.; Onjia, A. Gadolinium sorption on multi-walled carbon nanotubes. Contemp. Mater. 2019, 10, 1. [Google Scholar] [CrossRef]
- Siew, E.L.; Farris, A.F.; Rashid, N.; Chan, K.M.; Rajab, N.F. In Vitro Toxicological Assessment of Gadolinium (III) Chloride in V79-4 Fibroblasts. Genes Environ. 2020, 42, 22. [Google Scholar] [CrossRef]
- Sasikumar, P.; Narasimhan, S.V.; Velmurugan, S. Development of a Modified Ion Exchange Resin Column for Removal of Gadolinium From the Moderator System of PHWRs. Sep. Sci. Technol. 2013, 48, 1220–1225. [Google Scholar] [CrossRef]
- Port, M.; Idée, J.M.; Medina, C.; Robic, C.; Sabatou, M.; Corot, C. Efficiency, Thermodynamic and Kinetic Stability of Marketed Gadolinium Chelates and Their Possible Clinical Consequences: A Critical Review. BioMetals 2008, 21, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Yongxing, W.; Xiaorong, W.; Zichun, H. Genotoxicity of Lanthanum (III) and Gadolinium (III) in Human Peripheral Blood Lymphocytes. Bull. Environ. Contam. Toxicol. 2000, 64, 611–616. [Google Scholar] [CrossRef]
- Sappidi, P.; Boda, A.; Ali, S.M.; Singh, J.K. Adsorption of Gadolinium (Gd3+) Ions on the Dibenzo Crown Ether (DBCE) and Dicyclo Hexano Crown Ether (DCHCE) Grafted on the Polystyrene Surface: Insights from All Atom Molecular Dynamics Simulations and Experiments. J. Phys. Chem. C 2019, 123, 12276–12285. [Google Scholar] [CrossRef]
- Andrès, Y.; Thouand, G.; Boualam, M.; Mergeay, M. Factors Influencing the Biosorption of Gadolinium by Micro-Organisms and Its Mobilisation from Sand. Appl. Microbiol. Biotechnol. 2000, 54, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Breuker, A.; Ritter, S.F.; Schippers, A. Biosorption of Rare Earth Elements by Different Microorganisms in Acidic Solutions. Metals 2020, 10, 954. [Google Scholar] [CrossRef]
- Kumar, D.; Pandey, L.K.; Gaur, J.P. Metal sorption by algal biomass: From batch to continuous system. Algal Res. 2016, 18, 95–109. [Google Scholar] [CrossRef]
- Torres, E. Biosorption: A Review of the Latest Advances. Processes 2020, 8, 1584. [Google Scholar] [CrossRef]
- Chojnacka, K. Biosorption and Bioaccumulation—The Prospects for Practical Applications. Environ. Int. 2010, 36, 299–307. [Google Scholar] [CrossRef]
- Dev, S.; Sachan, A.; Dehghani, F.; Ghosh, T.; Briggs, B.R.; Aggarwal, S. Mechanisms of Biological Recovery of Rare-Earth Elements from Industrial and Electronic Wastes: A Review. Chem. Eng. J. 2020, 397, 124596. [Google Scholar] [CrossRef]
- Fujita, Y.; Walton, M.; Das, G.; Dohnalkova, A.; Vanzin, G.; Anderko, A. Impacts of Anthropogenic Gadolinium on the Activity of the Ammonia Oxidizing Bacterium Nitrosomonas Europaea. Chemosphere 2020, 257, 127250. [Google Scholar] [CrossRef]
- Ferreira, N.; Ferreira, A.; Viana, T.; Lopes, C.B.; Costa, M.; Pinto, J.; Soares, J.; Pinheiro-Torres, J.; Henriques, B.; Pereira, E. Assessment of Marine Macroalgae Potential for Gadolinium Removal from Contaminated Aquatic Systems. Sci. Total Environ. 2020, 749, 141488. [Google Scholar] [CrossRef]
- Ojima, Y.; Kosako, S.; Kihara, M.; Miyoshi, N.; Igarashi, K.; Azuma, M. Recovering Metals from Aqueous Solutions by Biosorption onto Phosphorylated Dry Baker’s Yeast. Sci. Rep. 2019, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Al-Amin, A.; Parvin, F.; Chakraborty, J.; Kim, Y.I. Cyanobacteria Mediated Heavy Metal Removal: A Review on Mechanism, Biosynthesis, and Removal Capability. Environ. Technol. Rev. 2021, 10, 44–57. [Google Scholar] [CrossRef]
- Kulal, D.K.; Loni, P.C.; Dcosta, C.; Some, S.; Kalambate, P.K. Cyanobacteria: As a Promising Candidate for Heavy-Metals Removal. In Advances in Cyanobacterial Biology; Academic Press: Cambridge, MA, USA, 2020; pp. 291–300. ISBN 9780128193112. [Google Scholar]
- Sarup, R.; Behl, K.; Joshi, M.; Nigam, S. Heavy Metal Removal by Cyanobacteria. In New Trends in Removal of Heavy Metals from Industrial Wastewater; Elsevier: Amsterdam, The Netherlands, 2021; pp. 441–466. ISBN 9780128229651. [Google Scholar]
- Ghorbani, E.; Nowruzi, B.; Nezhadali, M.; Hekmat, A. Metal Removal Capability of Two Cyanobacterial Species in Autotrophic and Mixotrophic Mode of Nutrition. BMC Microbiol. 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Application of Cyanobacteria Arthospira Platensis for Bioremediation of Erbium-Contaminated Wastewater. Materials 2022, 15, 6101. [Google Scholar] [CrossRef]
- Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Biosorption and Bioaccumulation Capacity of Arthospira Platensis toward Europium Ions. Water 2022, 14, 2128. [Google Scholar] [CrossRef]
- Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Biosorption and Bioaccumulation Capacity of Arthrospira Platensis toward Yttrium Ions. Metals 2022, 12, 1465. [Google Scholar] [CrossRef]
- Sayed, M.A.; Helal, A.I.; Abdelwahab, S.M.; Mahmoud, H.H.; Aly, H.F. Sorption and Possible Preconcentration of Europium and Gadolinium Ions from Aqueous Solutions by Mn3O4 Nanoparticles. Chem. Pap. 2020, 74, 619–630. [Google Scholar] [CrossRef]
- Abass, M.R.; El-Kenany, W.M.; Eid, M.A. Sorption of Cesium and Gadolinium Ions onto Zirconium Silico Antimonate Sorbent from Aqueous Solutions. Appl. Radiat. Isot. 2022, 192, 110542. [Google Scholar] [CrossRef] [PubMed]
- Cepoi, L.; Zinicovscaia, I.; Rudi, L.; Chiriac, T.; Rotari, I.; Turchenko, V.; Djur, S. Effects of PEG-Coated Silver and Gold Nanoparticles on Spirulina Platensis Biomass during Its Growth in a Closed System. Coatings 2020, 10, 717. [Google Scholar] [CrossRef]
- Cepoi, L.; Rudi, L.; Miscu, V.; Cojocari, A.; Chiriac, T.; Sadovnic, D. Antioxidative Activity of Ethanol Extracts from Spirulina Platensis and Nostoc Linckia Measured by Various Methods. An. Univ. Din Oradea Fasc. Biol. 2009, 16, 43–48. [Google Scholar]
- Lee, H.S.; Suh, J.H.; Kim, I.B.; Yoon, T. Effect of Aluminum in Two-Metal Biosorption by an Algal Biosorbent. Miner. Eng. 2004, 17, 487–493. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Safonov, A.; Tregubova, V.; Ilin, V.; Cepoi, L.; Chiriac, T.; Rudi, L.; Frontasyeva, M.V. Uptake of Metals from Single and Multi-Component Systems by Spirulina Platensis Biomass. Ecol. Chem. Eng. S 2016, 23, 401–412. [Google Scholar] [CrossRef]
- Sarada, B.; Krishna Prasad, M.; Kishore Kumar, K.; Murthy, C.V.R. Biosorption of Cd+2 by Green Plant Biomass, Araucaria Heterophylla: Characterization, Kinetic, Isotherm and Thermodynamic Studies. Appl. Water Sci. 2017, 7, 3483–3496. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Cepoi, L.; Rudi, L.; Chiriac, T.; Grozdov, D.; Pavlov, S.; Djur, S. Accumulation of Dysprosium, Samarium, Terbium, Lanthanum, Neodymium and Ytterbium by Arthrospira Platensis and Their Effects on Biomass Biochemical Composition. J. Rare Earths 2021, 39, 1133–1143. [Google Scholar] [CrossRef]
- Makhi, V.V.; Ahmad, A.; Chaugule, B.B. Effect of Bismuth, Gadolinium, and Cadmium Nanoparticles on Biomass, Carotenoid, and Lipid Content of Dunaliella Salina (Dunal) Teodoresco. Curr. Microbiol. 2022, 79, 30. [Google Scholar] [CrossRef]
- Goecke, F.; Vítová, M.; Lukavský, J.; Nedbalová, L.; Řezanka, T.; Zachleder, V. Effects of Rare Earth Elements on Growth Rate, Lipids, Fatty Acids and Pigments in Microalgae. Phycol. Res. 2017, 65, 226–234. [Google Scholar] [CrossRef]
- Tai, P.; Zhao, Q.; Su, D.; Li, P.; Stagnitti, F. Biological Toxicity of Lanthanide Elements on Algae. Chemosphere 2010, 80, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Luo, X.; Ren, Y.; Gao, E.; Gao, H. Effects of Yttrium and Phosphorus on Growth and Physiological Characteristics of Microcystis Aeruginosa. J. Rare Earths 2018, 36, 781–788. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetics | ||||||
| Pseudo-First-Order | Pseudo-Second-Order | |||||
| Parameters | qe | k1 | R2 | qe | k2 | R2 |
| Gd(III) | 1.24 | 19.8 | 0.98 | 1.24 | −0.6 | 0.97 |
| Isotherms | ||||||
| Model | Langmuir | Freundlich | ||||
| Parameters | qm | b | R2 | KF | n | R2 |
| Gd(III) | 101 | 0.0081 | 0.08 | 0.09 | 0.84 | 0.98 |
| Sorbent | q, mg/g | Reference |
|---|---|---|
| A. platensis | 101 | Present study |
| Multi-walled carbon nanotubes | 13.21 | [2] |
| Dibenzo crown ether | 1.83 | [7] |
| Dicyclo hexano crown ether | 2.02 | [7] |
| Manganese oxide nanoparticles | 12.6 | [24] |
| Zirconium silico antimonate | 27.2 | [25] |
| Temperature, K | ∆G°, kJ/mol | ∆H°, kJ/mol | ∆S°, J/mol·K |
|---|---|---|---|
| 293 | −11.81 | −14.1 | −7.8 |
| 303 | −11.73 | ||
| 313 | −11.65 | ||
| 323 | −11.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Cyanobacteria Arthospira platensis as an Effective Tool for Gadolinium Removal from Wastewater. Clean Technol. 2023, 5, 638-651. https://doi.org/10.3390/cleantechnol5020032
Yushin N, Zinicovscaia I, Cepoi L, Chiriac T, Rudi L, Grozdov D. Cyanobacteria Arthospira platensis as an Effective Tool for Gadolinium Removal from Wastewater. Clean Technologies. 2023; 5(2):638-651. https://doi.org/10.3390/cleantechnol5020032
Chicago/Turabian StyleYushin, Nikita, Inga Zinicovscaia, Liliana Cepoi, Tatiana Chiriac, Ludmila Rudi, and Dmitrii Grozdov. 2023. "Cyanobacteria Arthospira platensis as an Effective Tool for Gadolinium Removal from Wastewater" Clean Technologies 5, no. 2: 638-651. https://doi.org/10.3390/cleantechnol5020032