
Physiological Responses and Adaptations of the Halophyte Atriplex halimus to Soil Contaminated with Cd, Ni, and NaCl
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Soil
2.2. Experimental Design
2.3. Growth and Physiochemical Analysis
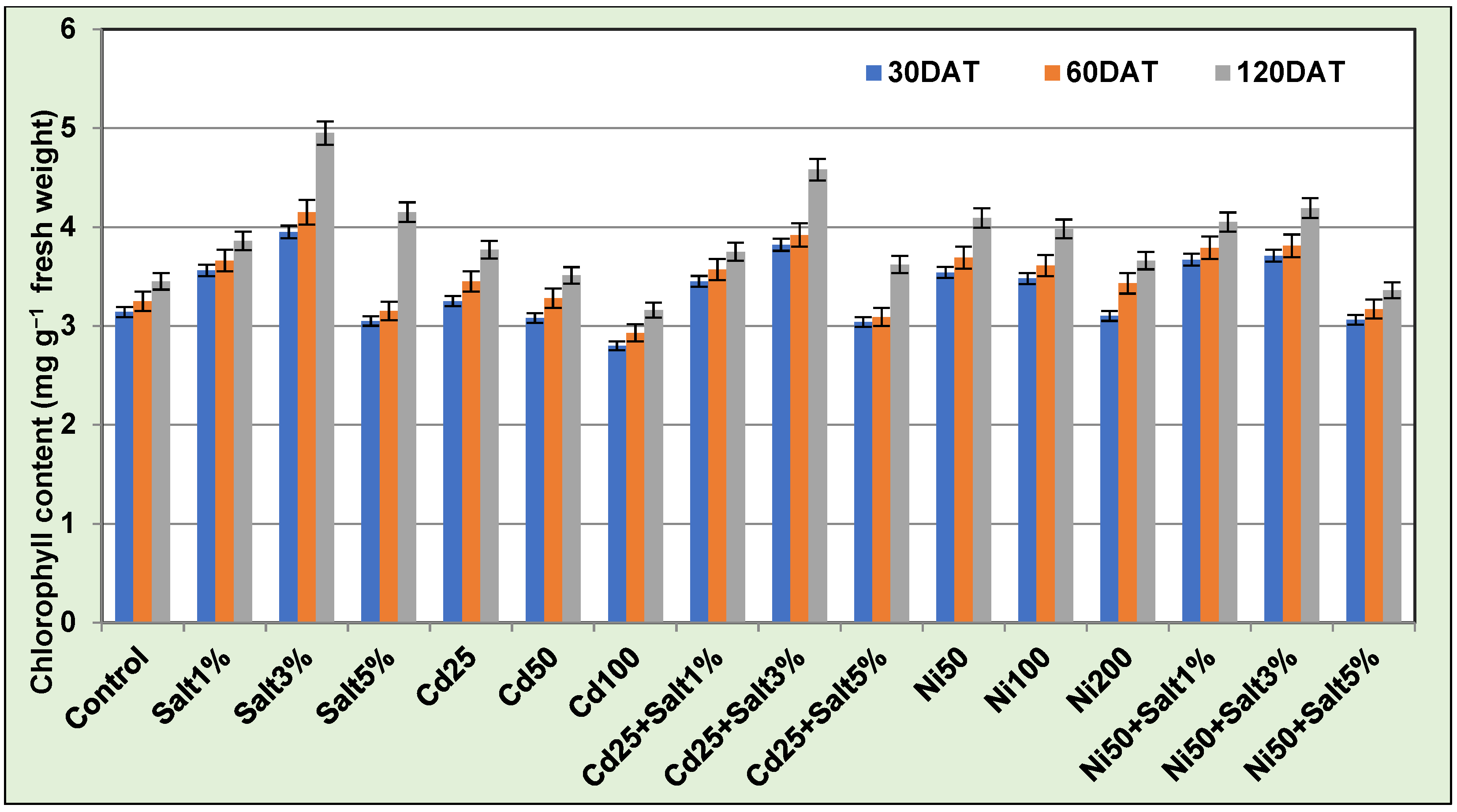
2.3.1. Chlorophyll
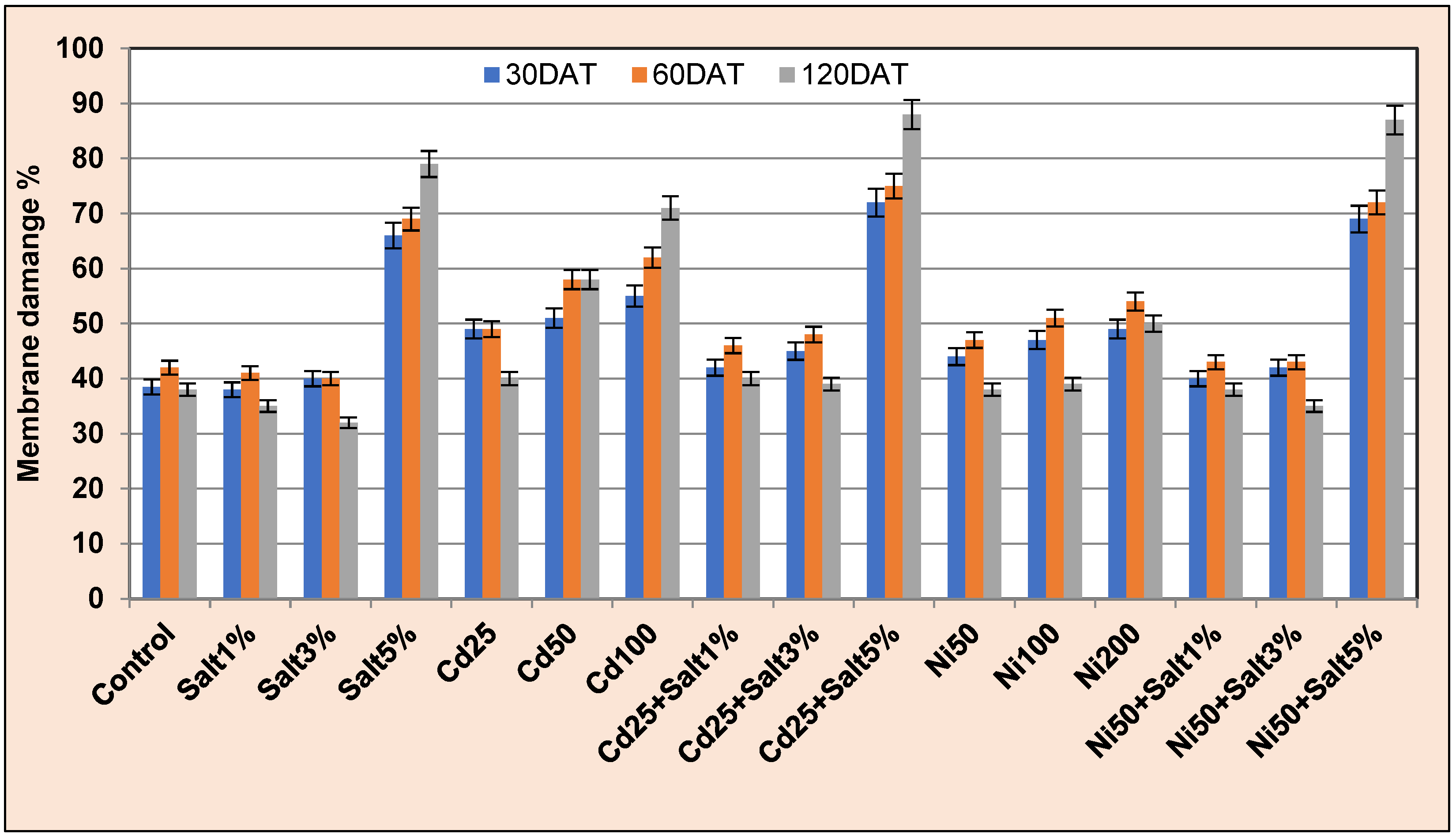
2.3.2. Membrane Damage Rate (MDR)
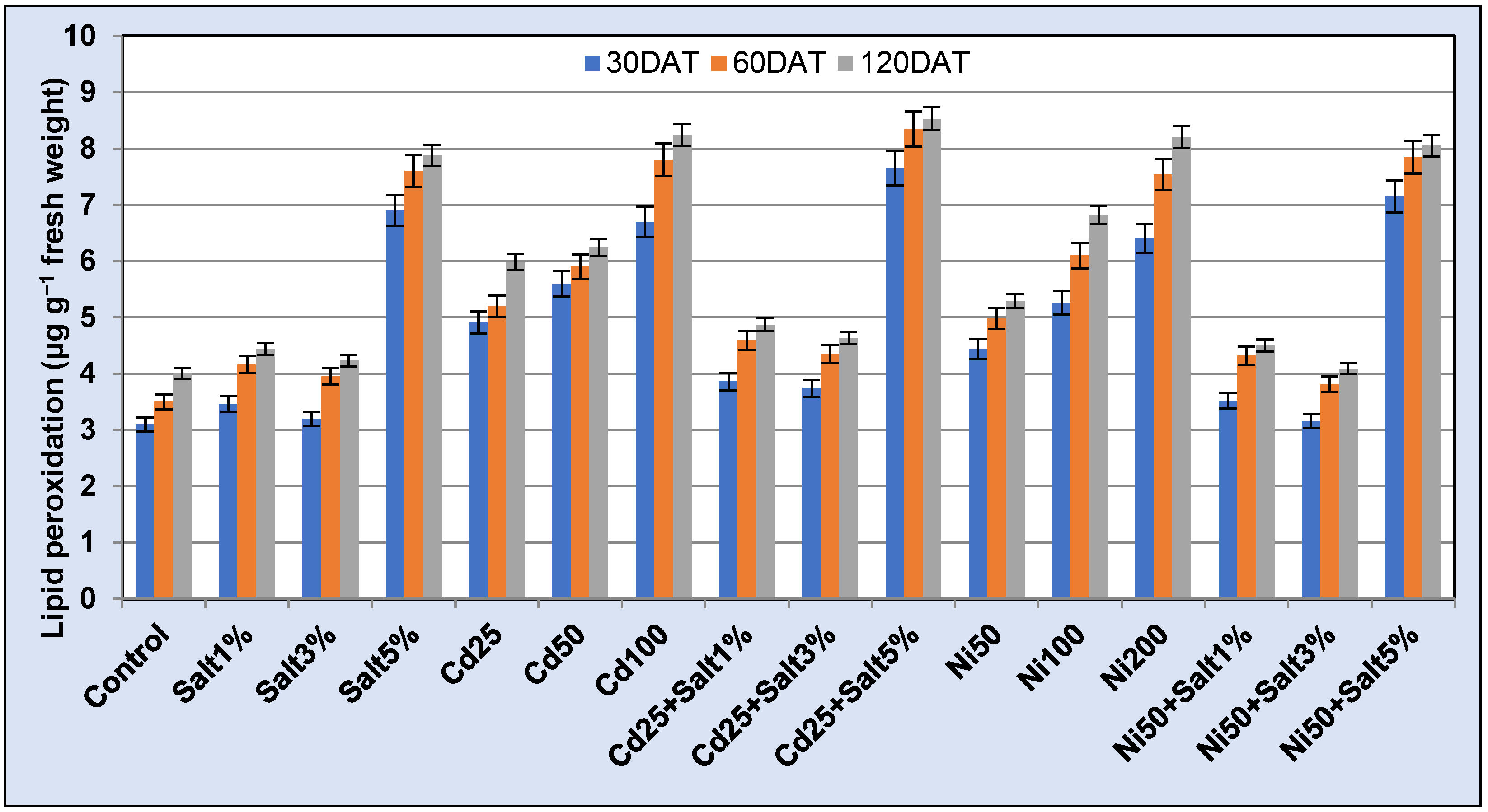
2.3.3. Lipid Peroxidation
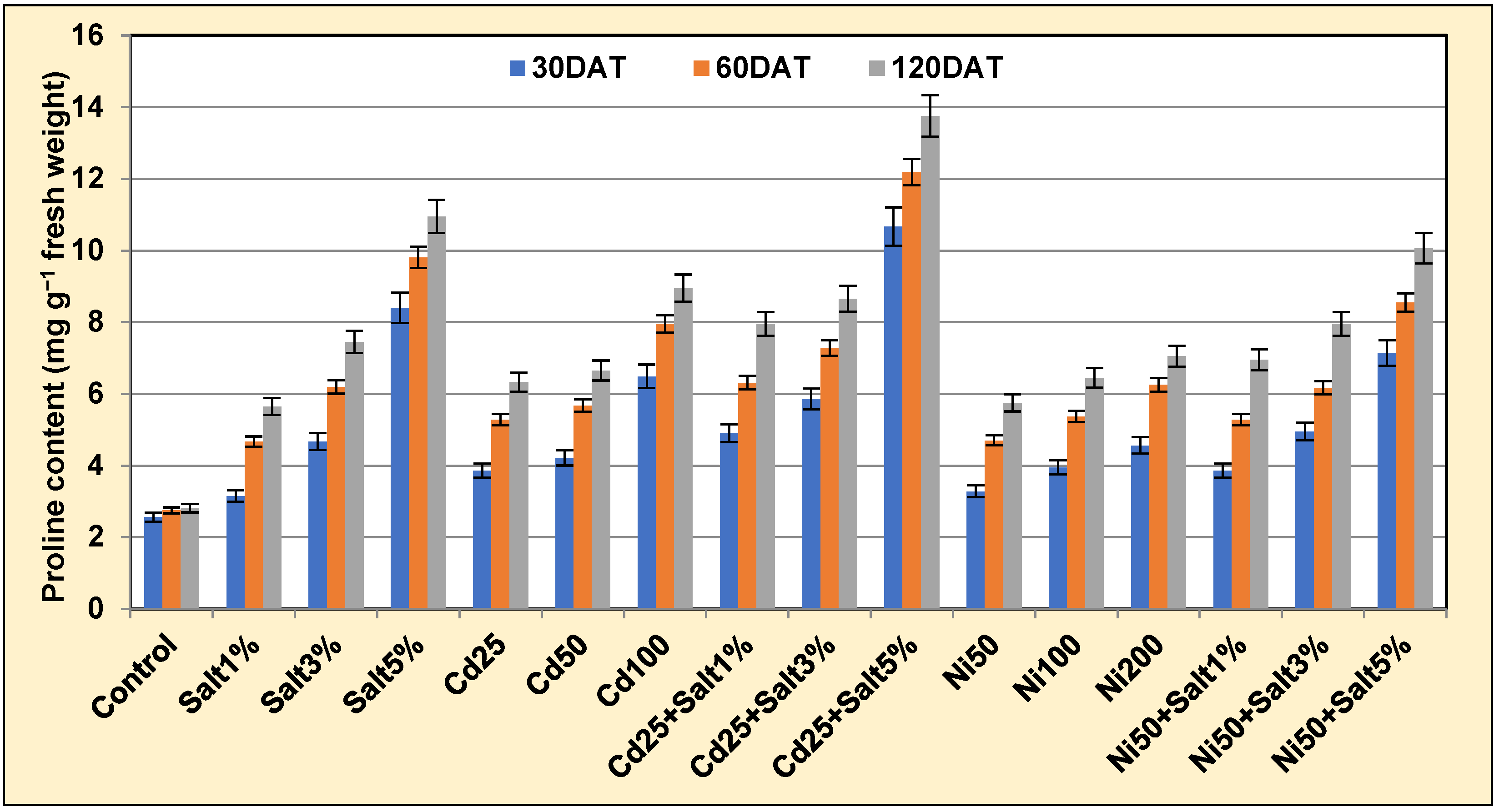
2.3.4. Proline
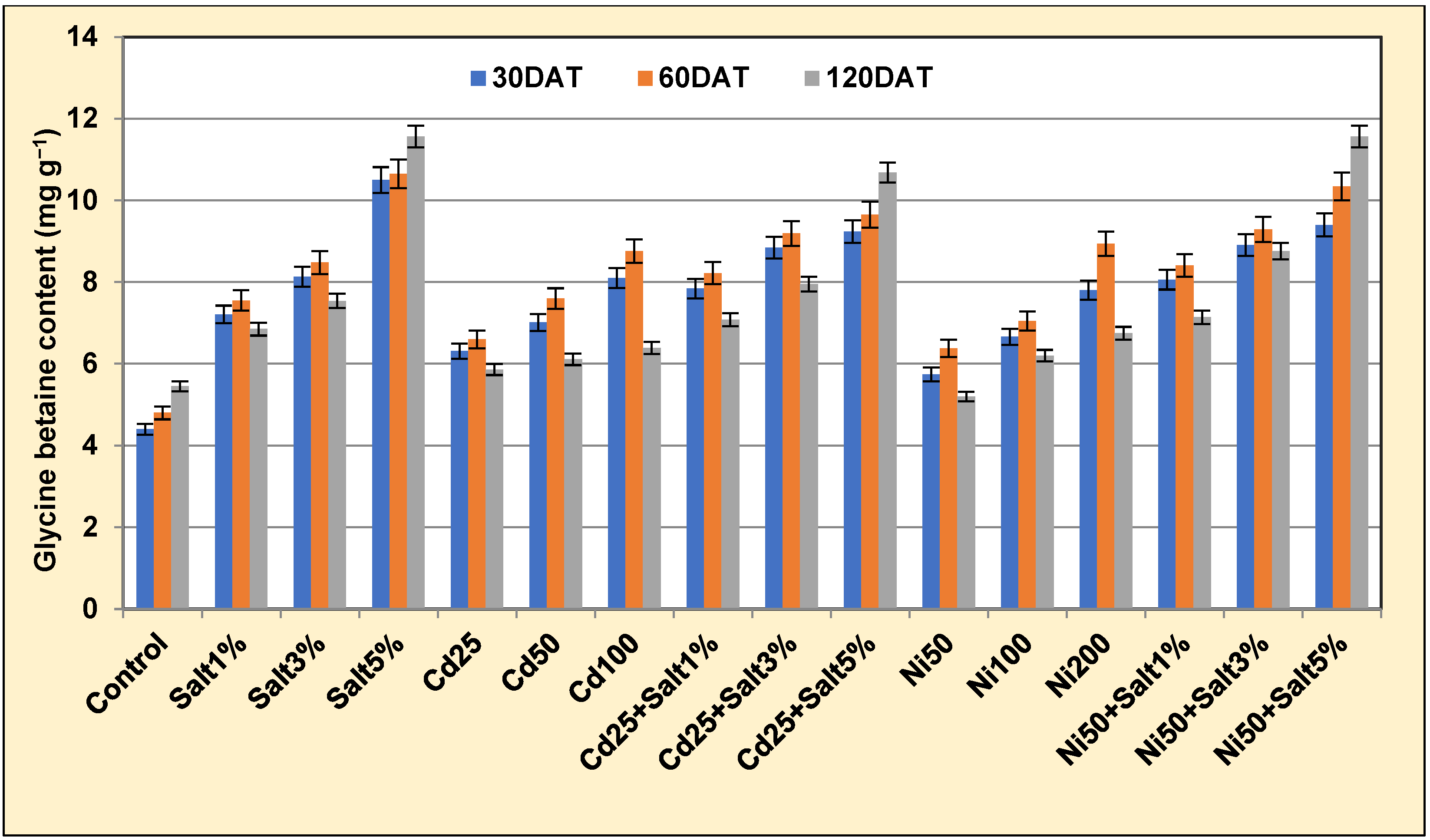
2.3.5. Glycine Betaine
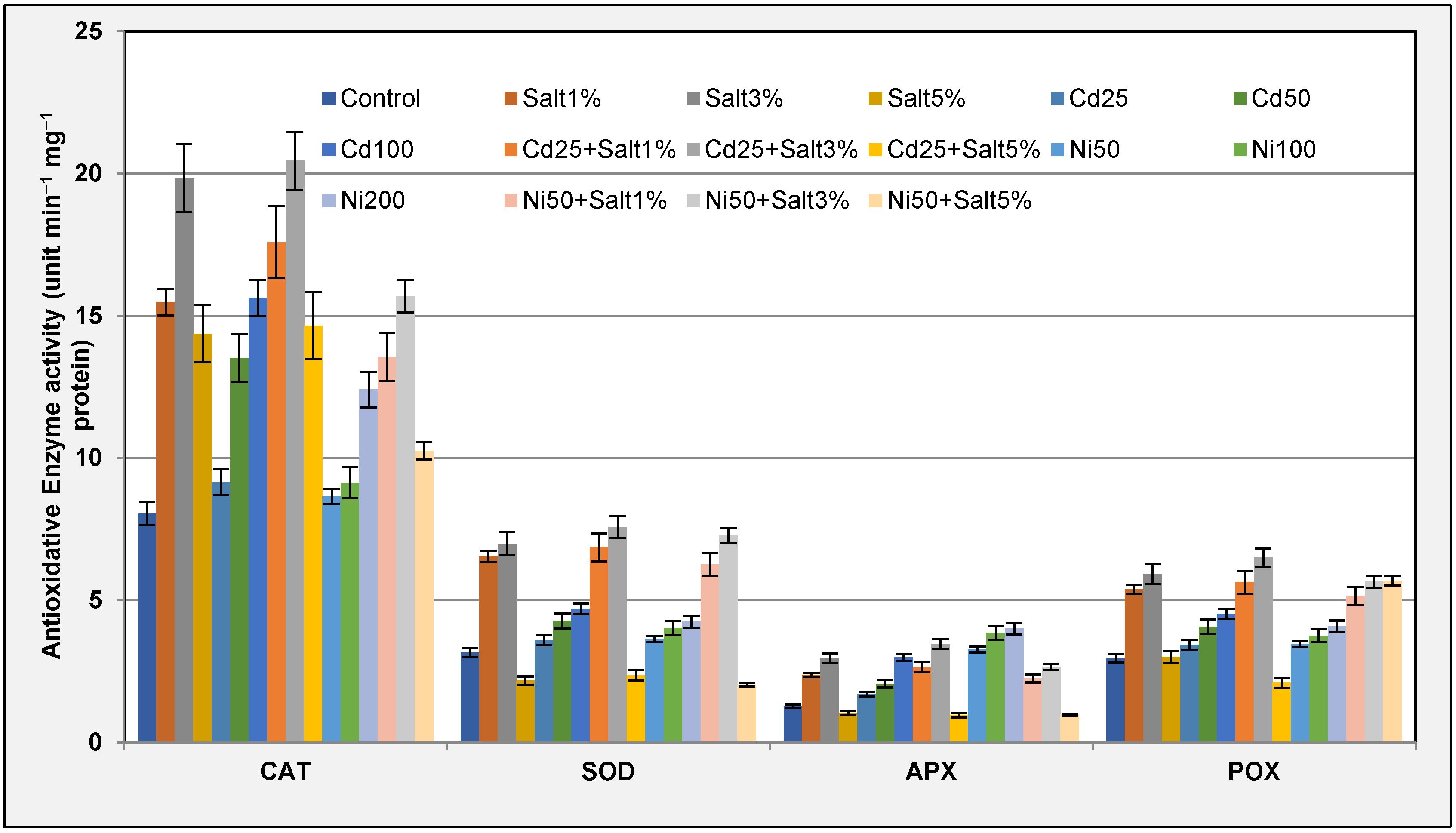
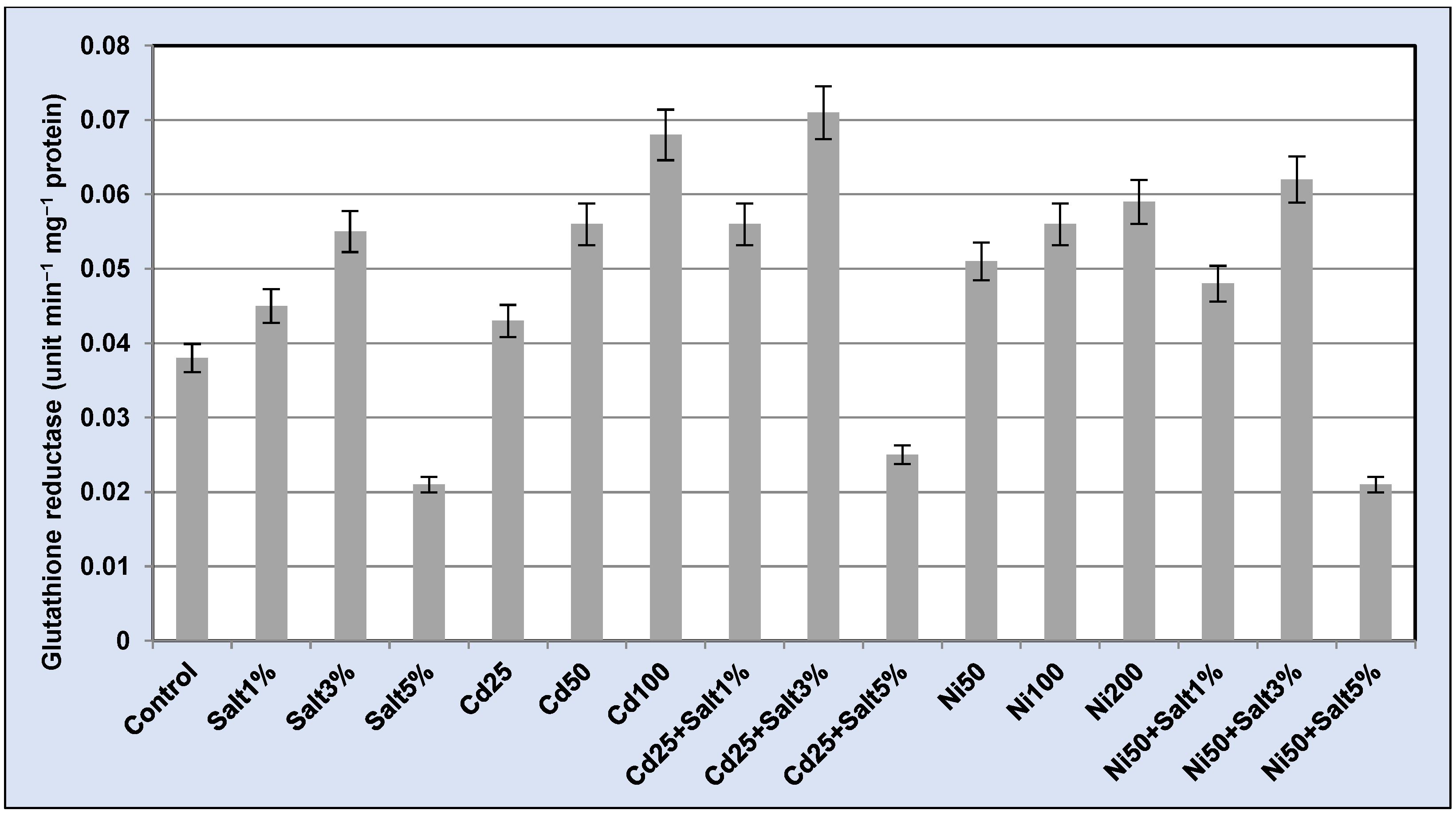
2.3.6. Antioxidative Enzyme Assay
2.3.7. Sodium Adsorption Ratio (SAR), Cd and Ni Analysis
2.4. Statistical Analysis
3. Results
3.1. PTE Accumulation and Phytoremedial Potential of A. halimus
3.2. Potential of A. halimus for Restoration of Saline Soil
3.3. Growth and Physiochemical Analysis
3.3.1. Dry Biomass and Root/Shoot Ratio
3.3.2. Chlorophyll Content
3.3.3. Membrane Damage Rate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Chlorophyll | Membrane Damage Rate | Lipid Peroxidation | Proline | Glycine Betaine | |
|---|---|---|---|---|---|---|
| Sources of Variations | ||||||
| Cd | Treatments | ** | ** | ** | ** | ** |
| Day after treatment | ** | ** | ** | ** | ** | |
| Treatment × DAT | Ns | ** | Ns | ** | ** | |
| Ni | Treatments | ** | ** | ** | ** | ** |
| Day after treatment | ** | ** | ** | ** | ** | |
| Treatment × DAT | Ns | ** | Ns | ** | ** | |
3.3.4. Lipid Peroxidation
3.3.5. Proline and Glycine Betaine Content
3.3.6. Antioxidative Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sihlahla, M.; Mouri, H.; Nomngongo, P.N. Assessment of bioavailability and mobility of major and trace elements in agricultural soils collected in Port St Johns, Eastern Cape, South Africa using single extraction procedures and pseudo-total digestion. J. Environ. Health Sci. Eng. 2020, 18, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Favas, P.J.C.; Pratas, J.; Varun, M.; Paul, M.S. Harnessing Pisum sativum–Glomus mosseae symbiosis for phytoremediation of soil contaminated with lead, cadmium, and arsenic. Int. J. Phytoremediat. 2021, 23, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Lutts, S.; Lefevre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef] [PubMed]
- Grigore, M.N.; Ivanescu, L.; Toma, C. Halophytes: An Integrative Anatomical Study; Springer: New York, NY, USA, 2014. [Google Scholar]
- Manousaki, E.; Kalogerakis, N. Phytoextraction of Pb and Cd by the Mediterranean Saltbush (Atriplex halimus L.): Metal uptake in relation to salinity. Environ. Sci. Pollut. Res. 2009, 16, 844–854. [Google Scholar] [CrossRef]
- Han, R.M.; Lefevre, I.; Albacete, A.; Perez-Alfocea, F.; Barba-Espín, G.; Díaz-Vivancos, P.; Quinet, M.; Ruan, C.J.; Hernandez, J.H.; Cantero-Navarro, E.; et al. Antioxidant enzyme activities and hormonal status in response to Cd stress in the wetland halophyte Kosteletzkya virginica under saline conditions. Physiol. Plant. 2012, 147, 352–368. [Google Scholar] [CrossRef]
- Gill, S.S.; Khan, A.N.; Tuteja, N. Cadmium at high dose perturbs growth, photosynthesis and nitrogen metabolism while at low dose it up regulates sulfur assimilation and antioxidant machinery in garden cress (Lepidium sativum L.). Plant Sci. 2012, 182, 112–120. [Google Scholar] [CrossRef]
- Huang, C.; Zeng, G.; Huang, D.; Lai, C.; Xu, P.; Zhang, C.; Cheng, M.; Wan, J.; Hu, L.; Zhang, Y. Effect of Phanerochaete chrysosporium inoculation on bacterial community and metal stabilization in lead-contaminated agricultural waste composting. Bioresour. Technol. 2017, 243, 294–303. [Google Scholar] [CrossRef]
- Mesnoua, M.; Mateos-Naranjo, E.; Barcia-Piedras, J.M.; Pérez-Romero, J.A.; Lotmani, B.; Redondo-Gómez, S. Physiological and biochemical mechanisms preventing Cd-toxicity in the hyperaccumulator Atriplex halimus L. Plant Physiol. Biochem. 2016, 106, 30–38. [Google Scholar] [CrossRef]
- Kahli, H.; Sbartai, H.; Cohen-Bouhacina, T.; Bourguignon, J. Characterization of cadmium accumulation and phytoextraction in three species of the genus Atriplex (canescens, halimus and nummularia) in the presence or absence of salt. Plant Physiol. Biochem. 2021, 166, 902–911. [Google Scholar] [CrossRef]
- Chen, C.; Huang, D.; Liu, J. Functions and toxicity of nickel in plants: Recent advances and future prospects. Clean 2009, 37, 304–313. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Abdelly, C. Nickel, cadmium and lead phytotoxicity and potential of halophytic plants in heavy metal extraction. S. Afr. J. Bot. 2017, 111, 99. [Google Scholar] [CrossRef]
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation mechanism of salt excluders under saline conditions and its applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef]
- Negrin, V.L.; Spetter, C.V.; Guinder, V.A.; Perillo, G.M.E.; Marcovecchio, J.E. The role of Sarcocornia perennis and tidal flooding on sediment biogeochemistry in a South American wetland. Mar. Biol. Res. 2013, 9, 703–715. [Google Scholar] [CrossRef]
- Cong, M.; Zhao, J.; Lu, J.; Ren, Z.; Wu, H. Homologous cloning, characterization and expression of a new halophyte phytochelatin synthase gene in Suaeda salsa. J. Oceanol. Limnol. 2016, 34, 1034. [Google Scholar] [CrossRef]
- Ghnaya, T.; Slama, I.; Messedi, D.; Grignon, C.; Abdelly, C. Cd-induced growth reduction in the halophyte Sesuvium portulacastrum is significantly improved by NaCl. J. Plant Res. 2007, 120, 309–316. [Google Scholar] [CrossRef]
- Eid, M.A. Halophytic plants for phytoremediation of heavy metals contaminated soil. J. Am. Sci. 2011, 7, 377–382. [Google Scholar]
- Chaturvedi, A.K.; Mishra, A.; Tiwari, V.; Jha, B. Cloning and transcript analysis of type 2 metallothionein gene (SbMT-2) from extreme halophyte Salicornia brachiata and its heterologous expression in E. coli. Gene 2012, 499, 280–287. [Google Scholar] [CrossRef]
- Lefevre, I.; Marchal, G.; Meerts, P.; Corréal, E.; Lutts, S. Chloride salinity reduces cadmium accumulation by the Mediterranean halophyte species Atriplex halimus L. Environ. Exp. Bot. 2009, 65, 142–152. [Google Scholar] [CrossRef]
- Kadukova, J.; Manousaki, E.; Papadantonakis, N.; Kalogerakis, N. Phytoextraction and phytoexcretion of Cd by the leaves of Tamarix smyrnensis growing on contaminated non-saline and saline soils. Environ. Res. 2008, 106, 326–332. [Google Scholar]
- Black, R.F. The leaf anatomy of Australian members of the genus Atriplex I. Atriplex vesicaria Heward and A. nummularia Lindl. Aust. J. Bot. 1954, 2, 269–286. [Google Scholar] [CrossRef]
- Sousa, A.I.; Caçador, I.; Lillebø, A.I.; Pardal, M.A. Heavy metal accumulation in Halimione portulacoides: Intra- and extra-cellular metal binding sites. Chemosphere 2008, 70, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes, Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Reclassification of Brevibacterium halotolerans DSM8802 as Bacillus halotolerans comb. nov. based on microbial and biochemical characterization and multiple gene sequence, rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water-soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Aebi, H. Catalase In Vitro, Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Dhindsa, R.; Plumb-Dhindsa, P.; Thorpe, T. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Smith, O.B.; Osafo, E.L.K.; Adegbola, A.A. Studies on the feeding value of agro-industrial by products: Strategies for improving the utilisation of cocoa-pod-based diets by ruminants. Anim. Feed Sci. Technol. 1988, 20, 189–201. [Google Scholar] [CrossRef]
- Rabhi, M.; Hafsi, C.; Lakhdar, A.; Hajji, S.; Barhoumi, Z.; Ham-Rouni, M.H.; Abdelly, C.; Smaoui, A. Evaluation of the capacity of three halophytes to desalinize their rhizosphere as grown on saline soils under non-leaching conditions. Afr. J. Ecol. 2009, 47, 463–468. [Google Scholar] [CrossRef]
- Vickerman, D.B.; Shannon, M.C.; Banuelos, G.S.; Grieve, C.M.; Trumble, J.T. Evaluation of Atriplex lines for selenium accumulation, salt tolerance and suitability for a key agricultural insect pest. Environ. Pollut. 2002, 120, 463–473. [Google Scholar] [CrossRef]
- Kachout, S.S.; Mansoura, A.B.; Mechergui, R.; Leclerc, J.C.; Rejeb, M.N.; Ouerghi, Z. Accumulation of Cu, Pb, Ni and Zn in the halophyte plant Atriplex grown on polluted soil. J. Sci. Food Agric. 2012, 92, 336–342. [Google Scholar] [CrossRef]
- Tapia, Y.; Diaz, O.; Pizarro, C.; Segura, R.; Vines, M.; Zúñiga, G.; Moreno-Jiménez, E. Atriplex atacamensis and Atriplex halimus resist As contamination in Pre-Andean soils [northern Chile]. Sci. Total Environ. 2013, 450–451, 188–196. [Google Scholar] [CrossRef]
- Ramos, J.; Naya, L.; Gay, M.; Abian, J.; Becana, M. Functional characterization of an unusual phytochelatin synthase, LjPCS3, of Lotus japonicas. Plant Physiol. 2002, 148, 536–545. [Google Scholar] [CrossRef]
- Amer, N.; Al Chami, Z.; Al Bitar, L.; Mondelli, D.; Dumontet, S. Evaluation of Atriplex halimus, Medicago lupulina and Portulaca oleracea for phytoremediation of Ni, Pb, and Zn. Int. J. Phytor. 2013, 15, 498–512. [Google Scholar] [CrossRef]
- Acosta, J.A.; Abbaspour, A.; Martínez, G.R.; Martínez-Martínez, S.; Zornoza, R.; Gabarrón, M.; Faz, A. Phytoremediation of mine tailings with Atriplex halimus and organic /inorganic amendments: A five-year field case study. Chemosphere 2018, 204, 71–78. [Google Scholar] [CrossRef]
- Bhatia, N.P.; Walsh, K.B.; Orlic, I.; Siegele, R.; Ashwath, N.; Baker, A.J.M. Studies on spatial distribution of nickel in leaves and stems of the metal hyperaccumulator Stackhousia tryonii Bailey using micro-PIXE and EDXS techniques. Funct. Plant Biol. 2004, 31, 1061–1074. [Google Scholar] [CrossRef]
- Yusuf, M.; Qazi, F.; Hayat, S.; Ahmad, A. Nickel: An overview of uptake, essentiality and toxicity in plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17. [Google Scholar] [CrossRef]
- USDA. Heavy Metal Soil Contamination; Soil Quality-Urban Technical Note No.3; U.S. Department of Agriculture: Washington, DC, USA, 2003.
- Lutts, S.; Lefèvre, I.; Delpéré, C.; Kivits, S.; Dechamps, C.; Robledo, A.; Correal, E. Heavy metal accumulation by the halophyte species mediterranean saltbush. J. Environ. Qual. 2004, 33, 1271–1279. [Google Scholar] [CrossRef]
- Kachout, S.; Leclerc, J.C.; Mansoura, A.B.; Rejeb, M.N.; Ouerghi, Z. Effects of heavy metals on growth and bioaccumulation of the annual halophytes Atriplex hortensis and A. rosea. J. Appl. Sci. Res. 2009, 5, 746–756. [Google Scholar]
- Mazharia, M.; Homaeed, M. Annual halophyte Chenopodium botrys can phytoextract cadmium from contaminated soils. J. Basic Appl. Sci. Res. 2012, 2, 1415–1422. [Google Scholar]
- Zaier, H.; Ghnaya, T.; Lakhdar, A.; Baioui, R.; Ghabriche, R.; Mnasri, M.; Sghaier, S.; Lutts, S.; Abdelly, C. Comparative study of Pb phytoextraction potential in Sesuvium portulacastrum and Brassica juncea: Tolerance and accumulation. J. Hazard. Mater. 2010, 183, 609–615. [Google Scholar] [CrossRef]
- Milic, Y.; He, H.; Li, X.; Zhong, T.; Hui, L.; Li, S. Enhancement of Cd phytoextraction by two Amaranthus species with endophytic Rahnella sp. JN27. Chemosphere 2012, 103, 99–104. [Google Scholar]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savoure, A. Diversity, distribution, and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Gonnelli, C.; Galardi, F.; Gabbrielli, R. Nickel and copper tolerance in three Tuscan populations of Silene paradoxa. Plant Physiol. 2001, 113, 507–514. [Google Scholar] [CrossRef]
- Yang, M.F.; Song, J.; Wang, B.S. Organ-Specific Responses of Vacuolar H+-ATPase in the Shoots and Roots of C3 Halophyte Suaeda salsa to NaCl. J. Integr. Plant Biol. 2010, 52, 308–314. [Google Scholar] [CrossRef]
- Daud, M.K.; Quiling, H.; Lei, M.; Ali, B.; Zhu, S.J. Ultrastructural, metabolic and proteomic changes in leaves of upland cotton in response to cadmium stress. Chemosphere 2015, 120, 309–320. [Google Scholar] [CrossRef]
- Santos, D.; Duarte, B.; Caçador, I. Unveiling Zn hyperaccumulation in Juncus acutus: Implications on the electronic energy fluxes and on oxidative stress with emphasis on non-functional Zn-chlorophylls. J. Photochem. Photobiol. 2014, B140, 228–239. [Google Scholar] [CrossRef]
- Ishtiyaq, S.; Kumar, H.; Clement, O.; Varun, M.; Paul, M.S. Role of Secondary Metabolites in Salt and Heavy Metal Stress Mitigation by Halophytic Plants: An Overview. In Handbook of Bioremediation; Mirza, H., Prasad, M.N.V., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2020; pp. 307–321. [Google Scholar]
- El-Bakatoushi, R.; Alframawy, A.M.; Tammam, A.; Youssef, D.; El-Sadek, L. Molecular and Physiological Mechanisms of Heavy Metal Tolerance in Atriplex halimus. Int. J. Phytor. 2015, 17, 789–800. [Google Scholar] [CrossRef]







| Treatments | Cd/Ni Content (mg kg−1) | Growth Parameters | ||||
|---|---|---|---|---|---|---|
| Soil | Root | Shoot | Dry Biomass (in g) | Root/Shoot Ratio | ||
| T0 | Control | nd | Nd | nd | 37.5 ± 1.7 | 0.25 |
| T1 | Salt1% | nd | Nd | nd | 51.2 ± 0.6 | 0.16 |
| T2 | Salt3% | nd | Nd | nd | 55.6 ± 1.0 | 0.16 |
| T3 | Salt5% | nd | Nd | nd | 32.4 ± 1.1 | 0.26 |
| Cd content (mg kg−1) | ||||||
| T4 | Cd25 | 23.6 ± 0.3 a | 6.41 ± 0.27 a | 2.61± 0.13 a | 38.6 ± 1.1 | 0.19 |
| T5 | Cd50 | 46.8 ± 0.7 b | 10.5 ± 0.33 b | 4.1 ± 0.18 b | 37.4 ± 0.6 | 0.19 |
| T6 | Cd100 | 96.5 ± 0.7 c | 16.7 ± 0.25 c | 8.0 ± 0.22 c | 36.8 ± 1.0 | 0.20 |
| T7 | Cd25 + Salt1% | 22.5 ± 0.3 d | 6.89 ± 0.24 d | 4.75 ± 0.23 d | 42.5 ± 0.3 | 0.15 |
| T8 | Cd25 + Salt3% | 21.8 ± 0.5 d | 7.41 ± 0.22 e | 6.97 ± 0.22 e | 49.8 ± 0.7 | 0.16 |
| T9 | Cd25 + Salt5% | 22.9 ± 0.6 a | 6.02 ± 0.14 a | 3.85 ± 0.17 b | 29.9 ± 2.6 | 0.26 |
| Ni content (mg kg−1) | ||||||
| T10 | Ni50 | 48.6 ± 0.4 a | 1.72 ± 0.18 a | 5.56 ± 0.33 a | 37.7 ± 0.8 | 0.18 |
| T11 | Ni100 | 98.2 ± 0.1 b | 2.52 ± 0.27 b | 7.14 ± 0.27 b | 36.4 ± 0.5 | 0.18 |
| T12 | Ni200 | 198.4 ± 0.5 c | 4.5 ± 0.22 c | 10.6 ± 0.22 c | 36.1 ± 0.5 | 0.18 |
| T13 | Ni50 + Salt1% | 46.8 ± 0.7 d | 1.92 ± 0.17 a | 7.12 ± 0.33 b | 53.5 ± 1.3 | 0.15 |
| T14 | Ni50 + Salt3% | 46.1 ± 0.8 d | 3.45 ± 0.22 d | 9.42 ± 0.24 d | 56.2 ± 1.6 | 0.13 |
| T15 | Ni50 + Salt5% | 49.2 ± 0.5 a | 1.24 ± 0.19 e | 2.42 ± 0.21 e | 30.2 ± 1.3 | 0.29 |
| Parameters | Biomass | CAT | SOD | APX | POX | GR |
|---|---|---|---|---|---|---|
| Sources of Variations | ||||||
| Cd | ** | ** | ns | ns | ns | ns |
| NaCl | ** | ** | ** | ** | ** | ns |
| Cd × NaCl | ** | ** | ns | ns | ns | ns |
| Ni | * | ** | ns | * | * | ns |
| NaCl | ** | ** | ** | ** | ** | ns |
| Ni × NaCl | * | ** | ns | ** | ** | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishtiyaq, S.; Kumar, H.; D’Souza, R.J.; Varun, M.; Favas, P.J.C.; Paul, M.S. Physiological Responses and Adaptations of the Halophyte Atriplex halimus to Soil Contaminated with Cd, Ni, and NaCl. Soil Syst. 2023, 7, 46. https://doi.org/10.3390/soilsystems7020046
Ishtiyaq S, Kumar H, D’Souza RJ, Varun M, Favas PJC, Paul MS. Physiological Responses and Adaptations of the Halophyte Atriplex halimus to Soil Contaminated with Cd, Ni, and NaCl. Soil Systems. 2023; 7(2):46. https://doi.org/10.3390/soilsystems7020046
Chicago/Turabian StyleIshtiyaq, Shumailah, Harsh Kumar, Rohan J. D’Souza, Mayank Varun, Paulo J. C. Favas, and Manoj S. Paul. 2023. "Physiological Responses and Adaptations of the Halophyte Atriplex halimus to Soil Contaminated with Cd, Ni, and NaCl" Soil Systems 7, no. 2: 46. https://doi.org/10.3390/soilsystems7020046