
Antioxidants Derived from Natural Products Reduce Radiative Damage in Cultured Retinal Glia to Prevent Oxidative Stress


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Reagents
2.3. Measurement of Cell Survival, Morphology, Proliferation, and GOS
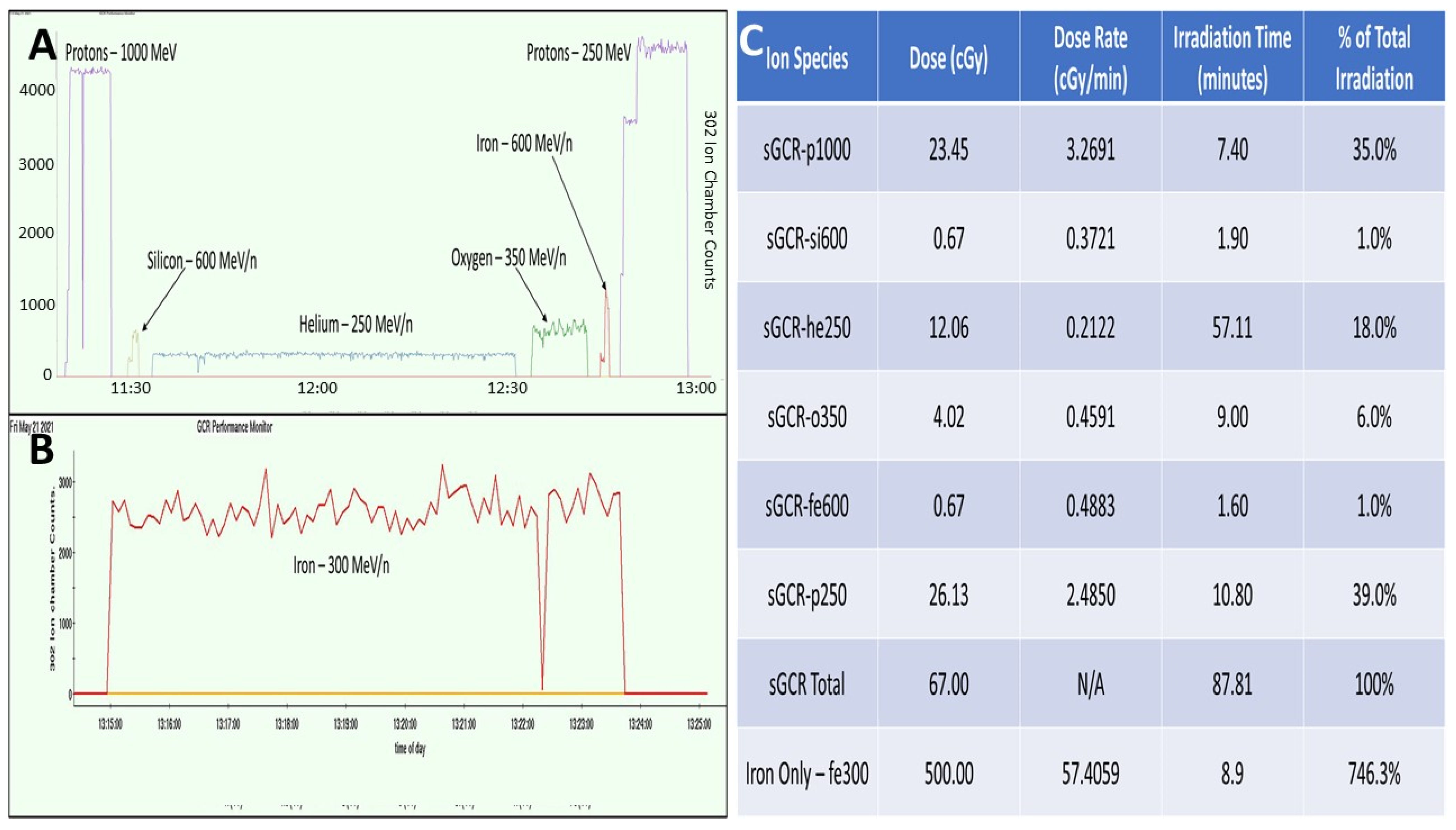
2.4. Galactic Cosmic Rays (GCR)
2.5. Imaging and Analyses
2.6. Statistical Analyses
3. Results
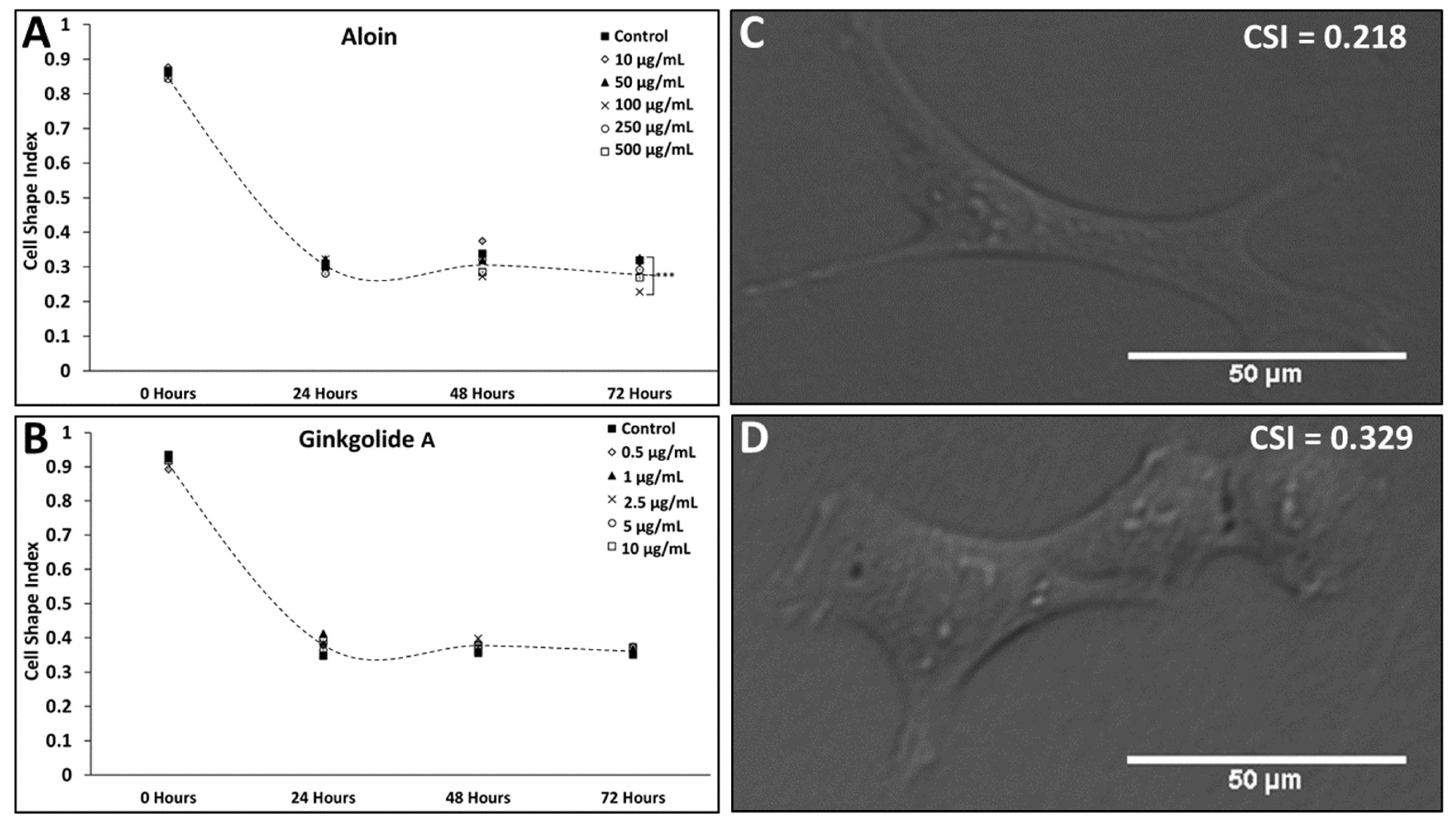
3.1. Changes in Müller Glia Morphology and Proliferation in Antioxidant Solutions
3.2. Changes in Cell Survival within Extracellular Solutions of Antioxidants
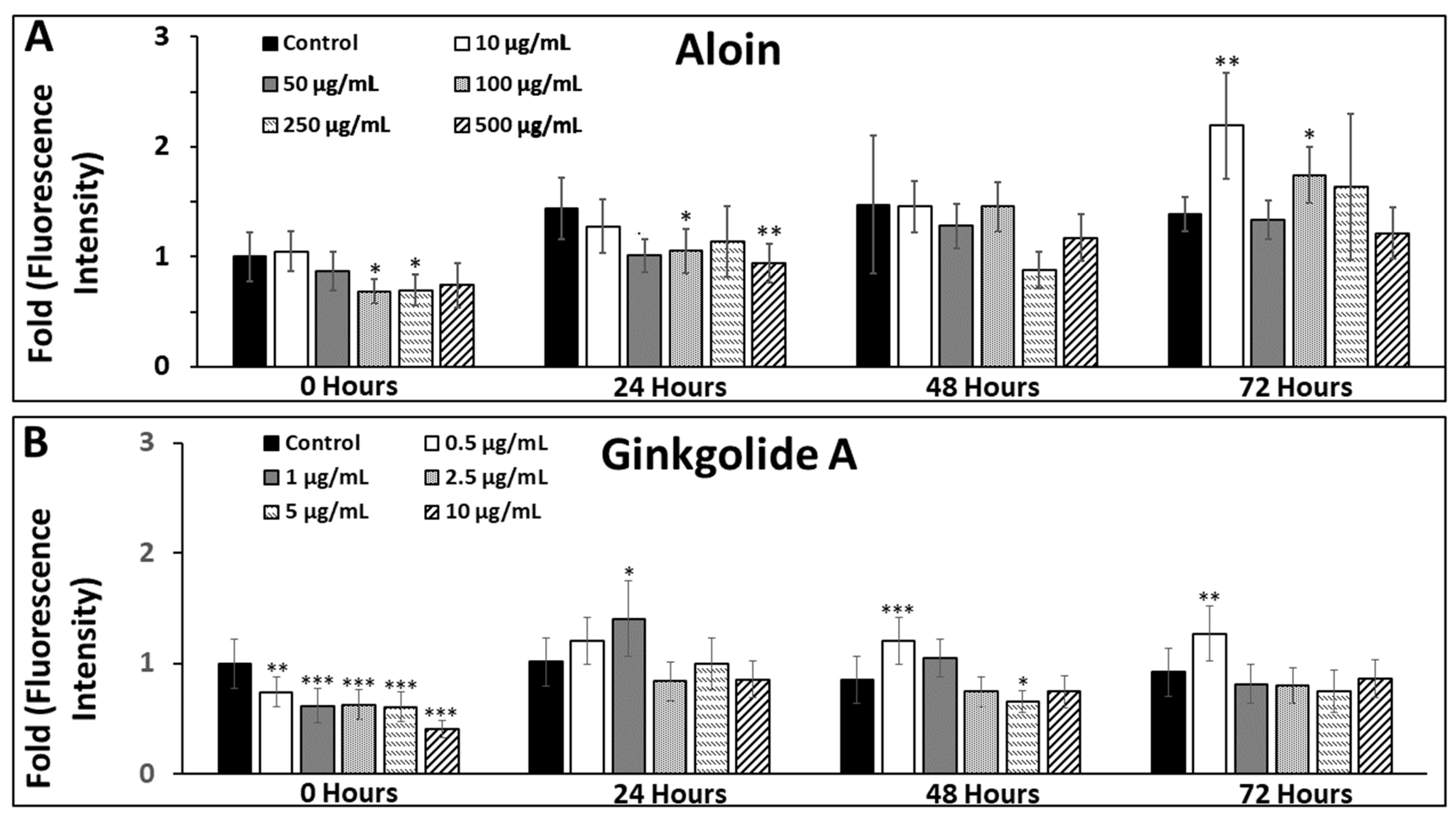
3.3. Changes in Oxidative Stress of Müller Glia within Extracellular Solutions
3.4. Changes in Survival and Oxidative Stress of Müller Glia Post-GCR

4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Martin Paez, Y.; Mudie, L.I.; Subramanian, P.S. Spaceflight Associated Neuro-Ocular Syndrome (SANS): A Systematic Review and Future Directions. Eye Brain 2020, 12, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Boerma, M.; Rodriguez, D.; Campbell-Beachler, M.; Jones, T.; Stanbouly, S.; Sridharan, V.; Nishiyama, N.C.; Wroe, A.; Nelson, G.A. Combined Effects of Low-Dose Proton Radiation and Simulated Microgravity on the Mouse Retina and the Hematopoietic System. Radiat. Res. 2019, 192, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.W.; Boerma, M.; Rodriguez, D.; Campbell-Beachler, M.; Jones, T.; Stanbouly, S.; Sridharan, V.; Wroe, A.; Nelson, G.A. Acute Effect of Low-Dose Space Radiation on Mouse Retina and Retinal Endothelial Cells. Radiat. Res. 2018, 190, 45–52. [Google Scholar] [CrossRef]
- Lett, J.T.; Cox, A.B.; Bergtold, D.S. Cellular and tissue responses to heavy ions: Basic considerations. Radiat. Environ. Biophys. 1986, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Loganovsky, K.N.; Marazziti, D.; Fedirko, P.A.; Kuts, K.V.; Antypchuk, K.Y.; Perchuk, I.V.; Babenko, T.F.; Loganovska, T.K.; Kolosynska, O.O.; Kreinis, G.Y.; et al. Radiation-Induced Cerebro-Ophthalmic Effects in Humans. Life 2020, 10, 41. [Google Scholar] [CrossRef]
- Vazquez, M.E.; Kirk, E. In vitro neurotoxic effects of 1 GeV/n iron particles assessed in retinal explants. Adv. Space Res. 2000, 25, 2041–2049. [Google Scholar] [CrossRef]
- Williams, G.R.; Lett, J.T. Damage to the photoreceptor cells of the rabbit retina from 56Fe ions: Effect of age at exposure, 1. Adv. Space Res. 1996, 18, 55–58. [Google Scholar] [CrossRef]
- Krebs, W.; Krebs, I.; Worgul, B.V. Effect of accelerated iron ions on the retina. Radiat. Res. 1990, 123, 213–219. [Google Scholar] [CrossRef]
- Nelson, A.C.; Hayes, T.L.; Tobias, C.A.; Yang, T.C. Some indications of structural damage in retina by heavy ion radiation. Scan. Electron. Microsc. 1981, 4, 79–85. [Google Scholar]
- Mohler, S.R. Galactic radiation exposure during commercial flights: Is there a risk? CMAJ 2003, 168, 1157–1158. [Google Scholar]
- Lerner, D.J.; Gorog, J.M., Jr. How “Rad” Is a Trip to Space? A Brief Discussion of Radiation Exposure in Suborbital Space Tourism. J. Am. Coll. Radiol. 2021, 18, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Kiser, P.D.; Golczak, M.; Maeda, A.; Palczewski, K. Key enzymes of the retinoid (visual) cycle in vertebrate retina. Biochim. Biophys. Acta 2012, 1821, 137–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveillard, T.; Sahel, J.A. Metabolic and redox signaling in the retina. Cell Mol. Life Sci. 2017, 74, 3649–3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, M.; Milliner, C.; Bell, B.A.; Bonilha, V.L. Oxidative stress in the retina and retinal pigment epithelium (RPE): Role of aging, and DJ-1. Redox Biol. 2020, 37, 101623. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Jiang, S.; Musayeva, A.; Gericke, A. Oxidative Stress and Vascular Dysfunction in the Retina: Therapeutic Strategies. Antioxidants 2020, 9, 761. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Bellezza, I. Oxidative Stress in Age-Related Macular Degeneration: Nrf2 as Therapeutic Target. Front. Pharmacol. 2018, 9, 1280. [Google Scholar] [CrossRef]
- Calderon, G.D.; Juarez, O.H.; Hernandez, G.E.; Punzo, S.M.; de la Cruz, Z.D. Oxidative stress and diabetic retinopathy: Development and treatment. Eye 2017, 31, 1122–1130. [Google Scholar] [CrossRef]
- Murakami, Y.; Nakabeppu, Y.; Sonoda, K.H. Oxidative Stress and Microglial Response in Retinitis Pigmentosa. Int. J. Mol. Sci. 2020, 21, 7170. [Google Scholar] [CrossRef]
- Chauhan, B.; Kumar, G.; Kalam, N.; Ansari, S.H. Current concepts and prospects of herbal nutraceutical: A review. J. Adv. Pharm. Technol. Res. 2013, 4, 4–8. [Google Scholar] [PubMed]
- Helal, N.A.; Eassa, H.A.; Amer, A.M.; Eltokhy, M.A.; Edafiogho, I.; Nounou, M.I. Nutraceuticals’ Novel Formulations: The Good, the Bad, the Unknown and Patents Involved. Recent Pat. Drug Deliv. Formul. 2019, 13, 105–156. [Google Scholar] [CrossRef] [PubMed]
- Lordan, R. Dietary supplements and nutraceuticals market growth during the coronavirus pandemic—Implications for consumers and regulatory oversight. Pharma Nutr. 2021, 18, 100282. [Google Scholar] [CrossRef] [PubMed]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar] [PubMed]
- Lobo, G.P.; Biswal, M.R.; Kondkar, A.A. Editorial: Molecular Mechanisms of Retinal Cell Degeneration and Regeneration. Front. Cell Dev. Biol. 2021, 9, 667028. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Deyev, A.I.; Yermakova, V.N.; Brikman, I.V.; Bours, J. Lipid peroxidation and cataracts: N-acetylcarnosine as a therapeutic tool to manage age-related cataracts in human and in canine eyes. Drugs R D 2004, 5, 125–139. [Google Scholar] [CrossRef]
- Rohowetz, L.J.; Kraus, J.G.; Koulen, P. Reactive Oxygen Species-Mediated Damage of Retinal Neurons: Drug Development Targets for Therapies of Chronic Neurodegeneration of the Retina. Int. J. Mol. Sci. 2018, 19, 3362. [Google Scholar] [CrossRef] [Green Version]
- Sanhueza Salas, L.F.; Garcia-Venzor, A.; Beltramone, N.; Capurro, C.; Toiber, D.; Silberman, D.M. Metabolic Imbalance Effect on Retinal Muller Glial Cells Reprogramming Capacity: Involvement of Histone Deacetylase SIRT6. Front. Genet. 2021, 12, 769723. [Google Scholar] [CrossRef]
- Wang, J.; O’Sullivan, M.L.; Mukherjee, D.; Punal, V.M.; Farsiu, S.; Kay, J.N. Anatomy and spatial organization of Muller glia in mouse retina. J. Comp. Neurol. 2017, 525, 1759–1777. [Google Scholar] [CrossRef]
- Kumar, A.; Pandey, R.K.; Miller, L.J.; Singh, P.K.; Kanwar, M. Muller glia in retinal innate immunity: A perspective on their roles in endophthalmitis. Crit. Rev. Immunol. 2013, 33, 119–135. [Google Scholar] [CrossRef]
- Hippert, C.; Graca, A.B.; Barber, A.C.; West, E.L.; Smith, A.J.; Ali, R.R.; Pearson, R.A. Muller glia activation in response to inherited retinal degeneration is highly varied and disease-specific. PLoS ONE 2015, 10, e0120415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graca, A.B.; Hippert, C.; Pearson, R.A. Muller Glia Reactivity and Development of Gliosis in Response to Pathological Conditions. Adv. Exp. Med. Biol. 2018, 1074, 303–308. [Google Scholar] [PubMed]
- Bringmann, A.; Wiedemann, P. Muller glial cells in retinal disease. Ophthalmologica 2012, 227, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Ackerman, K.M.; O’Hayer, P.; Bailey, T.J.; Gorsuch, R.A.; Hyde, D.R. Tumor necrosis factor-alpha is produced by dying retinal neurons and is required for Muller glia proliferation during zebrafish retinal regeneration. J. Neurosci. 2013, 33, 6524–6539. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.; Mysona, B.; Dun, Y.; Gnana-Prakasam, J.P.; Pabla, N.; Li, W.; Dong, Z.; Ganapathy, V.; Smith, S.B. Expression, subcellular localization, and regulation of sigma receptor in retinal muller cells. Invest. Ophthalmol. Vis. Sci. 2006, 47, 5576–5582. [Google Scholar] [CrossRef]
- Bringmann, A.; Grosche, A.; Pannicke, T.; Reichenbach, A. GABA and Glutamate Uptake and Metabolism in Retinal Glial (Muller) Cells. Front. Endocrinol. 2013, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Kay, A.M.; Simpson, C.L.; Stewart, J.A., Jr. The Role of AGE/RAGE Signaling in Diabetes-Mediated Vascular Calcification. J. Diabetes Res. 2016, 2016, 6809703. [Google Scholar] [CrossRef] [Green Version]
- Dorrell, M.I.; Aguilar, E.; Jacobson, R.; Yanes, O.; Gariano, R.; Heckenlively, J.; Banin, E.; Ramirez, G.A.; Gasmi, M.; Bird, A.; et al. Antioxidant or neurotrophic factor treatment preserves function in a mouse model of neovascularization-associated oxidative stress. J. Clin. Investig. 2009, 119, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Jung, E.; Kim, J. Aloin Inhibits Muller Cells Swelling in a Rat Model of Thioacetamide-Induced Hepatic Retinopathy. Molecules 2018, 23, 2806. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Jin, Z.; Xu, Z.; Yang, H.; Li, L.; Li, G.; Li, F.; Gu, S.; Zong, S.; Zhou, J.; et al. Antioxidant effects of ginkgolides and bilobalide against cerebral ischemia injury by activating the Akt/Nrf2 pathway in vitro and in vivo. Cell Stress Chaperones 2019, 24, 441–452. [Google Scholar] [CrossRef]
- Walchuk, C.; Suh, M. Nutrition and the aging retina: A comprehensive review of the relationship between nutrients and their role in age-related macular degeneration and retina disease prevention. Adv. Food Nutr. Res. 2020, 93, 293–332. [Google Scholar] [PubMed]
- Labkovich, M.; Jacobs, E.B.; Bhargava, S.; Pasquale, L.R.; Ritch, R. Ginkgo Biloba Extract in Ophthalmic and Systemic Disease, With a Focus on Normal-Tension Glaucoma. Asia Pac. J. Ophthalmol. 2020, 9, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Mo, X.F.; Jiang, X.H.; Rong, X.F.; Miao, H.M. Ginkgolide B promotes axonal growth of retina ganglion cells by anti-apoptosis in vitro. Sheng Li Xue Bao 2012, 64, 417–424. [Google Scholar] [PubMed]
- Sarthy, V.P.; Brodjian, S.J.; Dutt, K.; Kennedy, B.N.; French, R.P.; Crabb, J.W. Establishment and characterization of a retinal Muller cell line. Invest. Ophthalmol. Vis. Sci. 1998, 39, 212–216. [Google Scholar] [PubMed]
- Pena, J.S.; Vazquez, M. VEGF Upregulates EGFR Expression to Stimulate Chemotactic Behaviors in the rMC-1 Model of Muller Glia. Brain Sci. 2020, 10, 330. [Google Scholar] [CrossRef]
- Trueblood, K.E.; Mohr, S.; Dubyak, G.R. Purinergic regulation of high-glucose-induced caspase-1 activation in the rat retinal Muller cell line rMC-1. Am. J. Physiol. Cell Physiol. 2011, 301, C1213–C1223. [Google Scholar] [CrossRef]
- Xue, W.; Du, P.; Lin, S.; Dudley, V.J.; Hernandez, M.R.; Sarthy, V.P. Gene expression changes in retinal Muller (glial) cells exposed to elevated pressure. Curr. Eye Res. 2011, 36, 754–767. [Google Scholar] [CrossRef]
- Cliver, R.N.; Ayers, B.; Brady, A.; Firestein, B.L.; Vazquez, M. Cerebrospinal fluid replacement solutions promote neuroglia migratory behaviors and spinal explant outgrowth in microfluidic culture. J. Tissue Eng. Regen. Med. 2021, 15, 176–188. [Google Scholar] [CrossRef]
- Pena, J.; Dulger, N.; Singh, T.; Zhou, J.; Majeska, R.; Redenti, S.; Vazquez, M. Controlled microenvironments to evaluate chemotactic properties of cultured Muller glia. Exp. Eye Res. 2018, 173, 129–137. [Google Scholar] [CrossRef]
- Grosche, A.; Hauser, A.; Lepper, M.F.; Mayo, R.; von Toerne, C.; Merl-Pham, J.; Hauck, S.M. The Proteome of Native Adult Muller Glial Cells from Murine Retina. Mol. Cell Proteom. 2016, 15, 462–480. [Google Scholar] [CrossRef] [Green Version]
- Norbury, J.W.; Schimmerling, W.; Slaba, T.C.; Azzam, E.I.; Badavi, F.F.; Baiocco, G.; Benton, E.; Bindi, V.; Blakely, E.A.; Blattnig, S.R.; et al. Galactic cosmic ray simulation at the NASA Space Radiation Laboratory. Life Sci. Space Res. 2016, 8, 38–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichenbach, A.; Bringmann, A. Glia of the human retina. Glia 2020, 68, 768–796. [Google Scholar] [CrossRef] [PubMed]
- Philips, T.; Rothstein, J.D. Oligodendroglia: Metabolic supporters of neurons. J. Clin. Investig. 2017, 127, 3271–3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, D. Muller glial cell reprogramming and retina regeneration. Nat. Rev. Neurosci. 2014, 15, 431–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, G.; Volpi, L.; Pasquali, L.; Petrozzi, L.; Siciliano, G. Astrocyte-neuron interactions in neurological disorders. J. Biol. Phys. 2009, 35, 317–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hladky, S.B.; Barrand, M.A. Mechanisms of fluid movement into, through and out of the brain: Evaluation of the evidence. Fluids Barriers CNS 2014, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardourel, M.; Felgerolle, C.; Paris, A.; Acar, N.; Othman, K.R.B.; Ueda, N.; Rossignol, R.; Bazinet, A.; Hebert, B.; Briault, S.; et al. Dietary Supplement Enriched in Antioxidants and Omega-3 Promotes Glutamine Synthesis in Muller Cells: A Key Process against Oxidative Stress in Retina. Nutrients 2021, 13, 3216. [Google Scholar] [CrossRef]
- Toft-Kehler, A.K.; Skytt, D.M.; Kolko, M. A Perspective on the Muller Cell-Neuron Metabolic Partnership in the Inner Retina. Mol. Neurobiol. 2018, 55, 5353–5361. [Google Scholar] [CrossRef]
- Reichenbach, A.; Bringmann, A. New functions of Muller cells. Glia 2013, 61, 651–678. [Google Scholar] [CrossRef]
- Kuse, Y.; Ogawa, K.; Tsuruma, K.; Shimazawa, M.; Hara, H. Damage of photoreceptor-derived cells in culture induced by light emitting diode-derived blue light. Sci. Rep. 2014, 4, 5223. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.J.; Kaja, S.; Naumchuk, Y.; Kunjukunju, N.; Koulen, P. Antioxidant drug therapy approaches for neuroprotection in chronic diseases of the retina. Int. J. Mol. Sci. 2014, 15, 1865–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastlake, K.; Luis, J.; Limb, G.A. Potential of Muller Glia for Retina Neuroprotection. Curr. Eye Res. 2020, 45, 339–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groeger, G.; Doonan, F.; Cotter, T.G.; Donovan, M. Reactive oxygen species regulate prosurvival ERK1/2 signaling and bFGF expression in gliosis within the retina. Invest. Ophthalmol. Vis. Sci. 2012, 53, 6645–6654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassagne, F.; Huang, X.; Lyles, J.T.; Quave, C.L. Validation of a 16th Century Traditional Chinese Medicine Use of Ginkgo biloba as a Topical Antimicrobial. Front. Microbiol. 2019, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Chassagne, F.; Samarakoon, T.; Porras, G.; Lyles, J.T.; Dettweiler, M.; Marquez, L.; Salam, A.M.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. A Systematic Review of Plants with Antibacterial Activities: A Taxonomic and Phylogenetic Perspective. Front. Pharmacol. 2020, 11, 586548. [Google Scholar] [CrossRef]
- Bosch-Morell, F.; Villagrasa, V.; Ortega, T.; Acero, N.; Munoz-Mingarro, D.; Gonzalez-Rosende, M.E.; Castillo, E.; Sanahuja, M.A.; Soriano, P.; Martinez-Solis, I. Medicinal plants and natural products as neuroprotective agents in age-related macular degeneration. Neural Regen. Res. 2020, 15, 2207–2216. [Google Scholar]
- Mojaverrostami, S.; Bojnordi, M.N.; Ghasemi-Kasman, M.; Ebrahimzadeh, M.A.; Hamidabadi, H.G. A Review of Herbal Therapy in Multiple Sclerosis. Adv. Pharm. Bull. 2018, 8, 575–590. [Google Scholar] [CrossRef]
- Tewari, D.; Samoila, O.; Gocan, D.; Mocan, A.; Moldovan, C.; Devkota, H.P.; Atanasov, A.G.; Zengin, G.; Echeverria, J.; Vodnar, D.; et al. Medicinal Plants and Natural Products Used in Cataract Management. Front. Pharmacol. 2019, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Y.; Chen, C.Z.; Su, Y.; Li, L.; Yi, Z.H.; Qi, H.; Weng, M.; Xing, Y.Q. Effect of EGb761 on light-damaged retinal pigment epithelial cells. Int. J. Ophthalmol. 2014, 7, 8–13. [Google Scholar]
- St-Pierre, J.; Drori, S.; Uldry, M.; Silvaggi, J.M.; Rhee, J.; Jager, S.; Handschin, C.; Zheng, K.; Lin, J.; Yang, W.; et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell 2006, 127, 397–408. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aloin | Control | 10 ug/mL | 50 ug/mL | 100 ug/mL | 250 ug/mL | 500 ug/mL |
| 72 h | 97.0 ± 5.6% | 94.4 ± 3.7% | 93.4 ± 4.5% | 90.5 ± 3.5% * | 73.5 ± 13.0% *** | 63.3 ± 10.0% *** |
| Ginkgolide A | Control | 0.5 µg/mL | 1 µg/mL | 2.5 µg/mL | 5 µg/mL | 10 µg/mL |
| 72 h | 99.3 ± 0.2% | 99.5 ± 0.5% | 98.7 ± 0.3% * | 96.3 ± 1.6% * | 95.8 ± 3.7% | 93.8 ± 2.7% * |
| Pre-Treatment Survival | Post-Treatment Survival | (Pre − Post) Survival Difference | ||
|---|---|---|---|---|
| 30 min | ||||
| Aloin Control | 92.9% | Aloin Control | 92.9% | 0.0% |
| Aloin 50 µg/mL | 92.2% | Aloin 50 µg/mL | 92.1% | 0.1% |
| Aloin 100 µg/mL | 94.6% | Aloin 100 µg/mL | 93.3% | 1.3% |
| Ginkgolide A Control | 95.1% | Ginkgolide A Control | 94.1% | 1.0% |
| Ginkgolide A 1 µg/mL | 95.5% | Ginkgolide A 1 µg/mL | 94.7% | 0.8% |
| Ginkgolide A 2.5 µg/mL | 93.2% | Ginkgolide A 2.5 µg/mL | 95.1% | − 1.9% |
| 6 h | ||||
| Aloin Control | 92.7% | Aloin Control | 91.1% | 1.6% |
| Aloin 50 µg/mL | 91.3% | Aloin 50 µg/mL | 95.8% | − 4.5% |
| Aloin 100 µg/mL | 94.7% | Aloin 100 µg/mL | 90.3% | 4.4% |
| Ginkgolide A Control | 91.3% | Ginkgolide A Control | 94.2% | − 2.9% |
| Ginkgolide A 1 µg/mL | 91.9% | Ginkgolide A 1 µg/mL | 94.8% | − 2.9% |
| Ginkgolide A 2.5 µg/mL | 85.2% | Ginkgolide A 2.5 µg/mL | 95.7% | − 10.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cliver, R.N.; Castro, N.; Russomano, T.; Lardieri, G.; Quarrie, L.; van der Merwe, H.; Vazquez, M. Antioxidants Derived from Natural Products Reduce Radiative Damage in Cultured Retinal Glia to Prevent Oxidative Stress. Neuroglia 2022, 3, 84-98. https://doi.org/10.3390/neuroglia3030006
Cliver RN, Castro N, Russomano T, Lardieri G, Quarrie L, van der Merwe H, Vazquez M. Antioxidants Derived from Natural Products Reduce Radiative Damage in Cultured Retinal Glia to Prevent Oxidative Stress. Neuroglia. 2022; 3(3):84-98. https://doi.org/10.3390/neuroglia3030006
Chicago/Turabian StyleCliver, Richard N., Natalia Castro, Thais Russomano, Gaetano Lardieri, Lindsay Quarrie, Helena van der Merwe, and Maribel Vazquez. 2022. "Antioxidants Derived from Natural Products Reduce Radiative Damage in Cultured Retinal Glia to Prevent Oxidative Stress" Neuroglia 3, no. 3: 84-98. https://doi.org/10.3390/neuroglia3030006