
Species-Specific Responses of Medium and Large Mammals to Fire Regime Attributes in a Fire-Prone Neotropical Savanna

Abstract
:1. Introduction
2. Materials and Methods
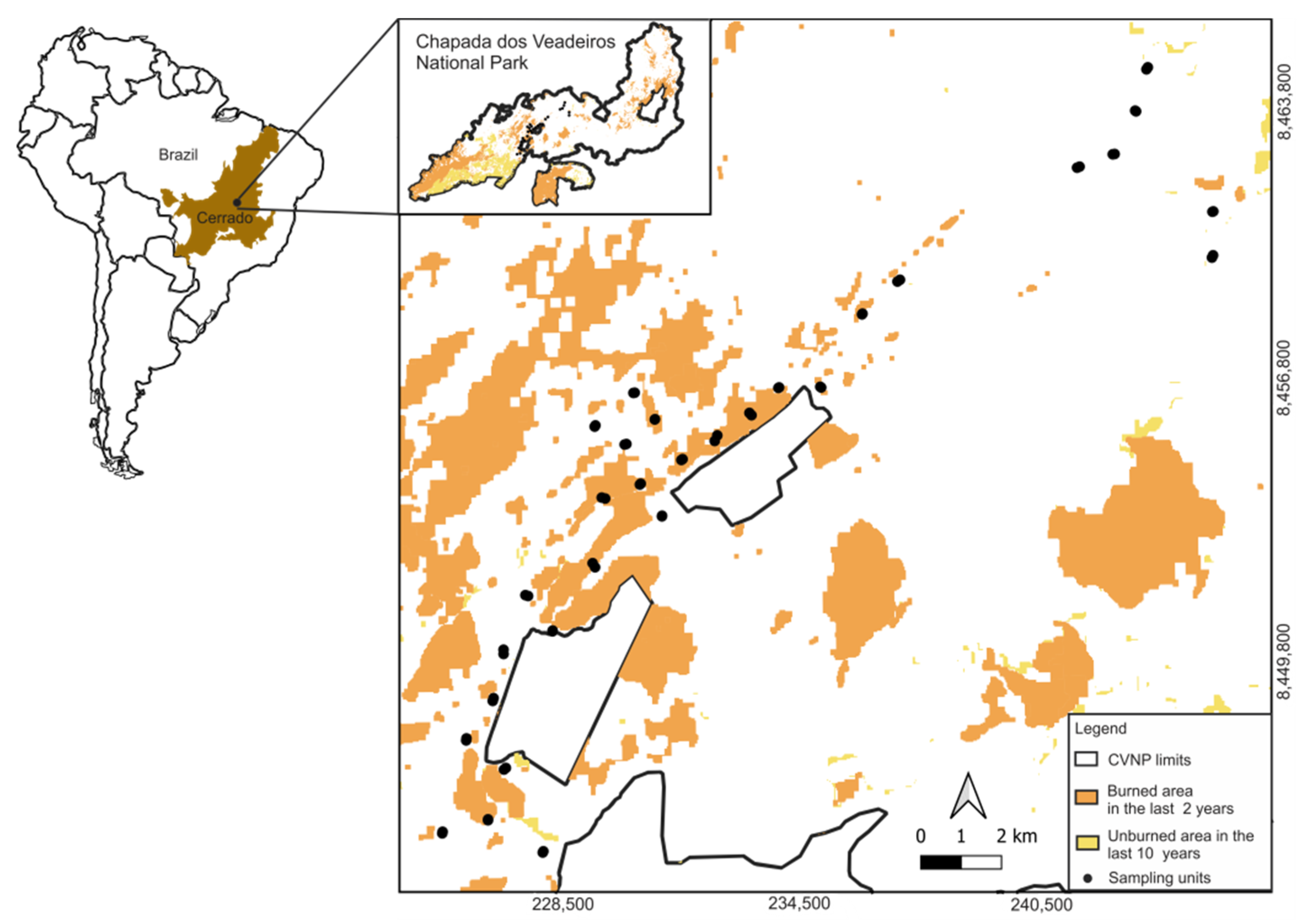
2.1. Study Area and Species
2.2. Sampling of Medium and Large Mammals
2.3. Predictor Variables
2.4. Statistical Analysis
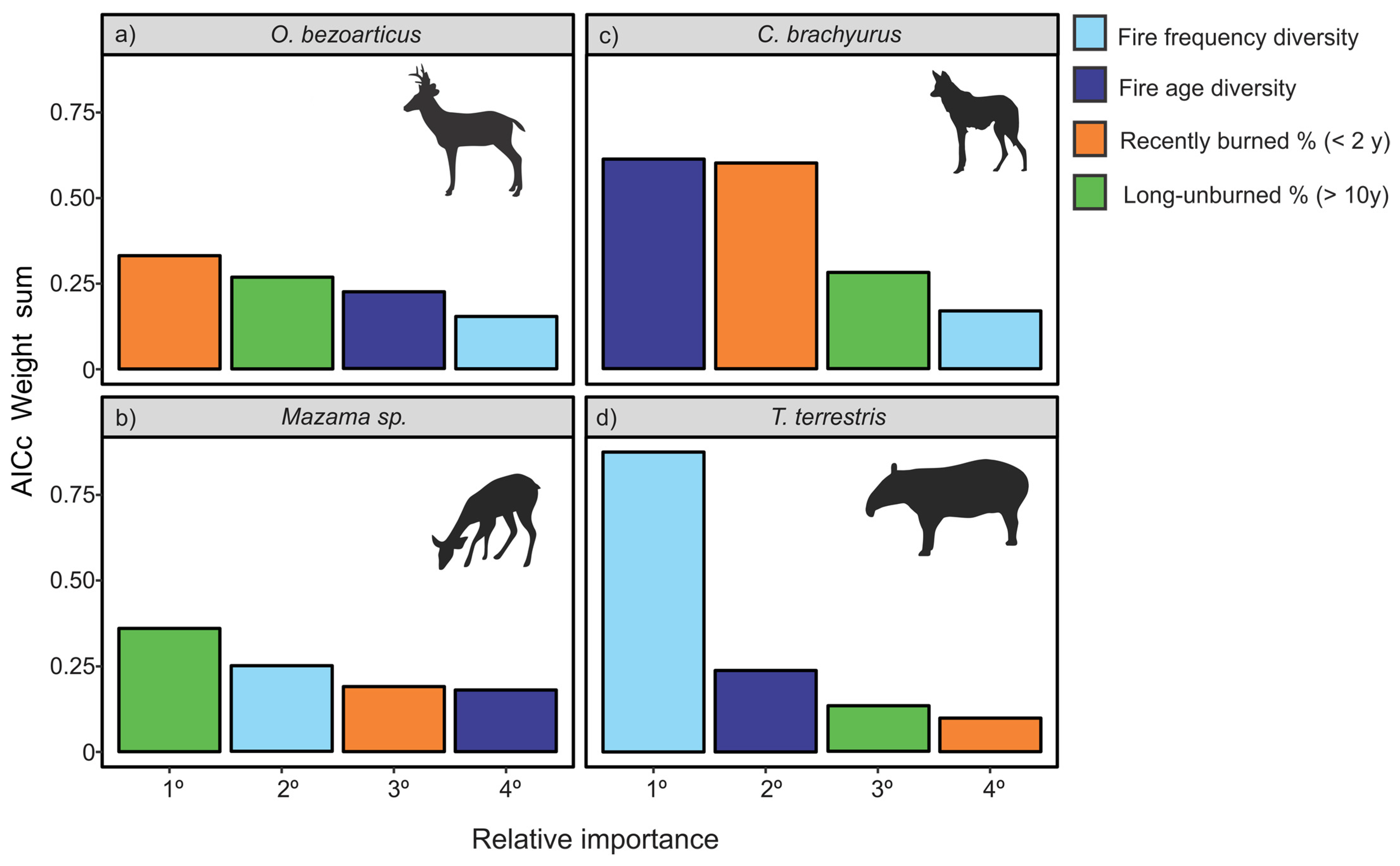
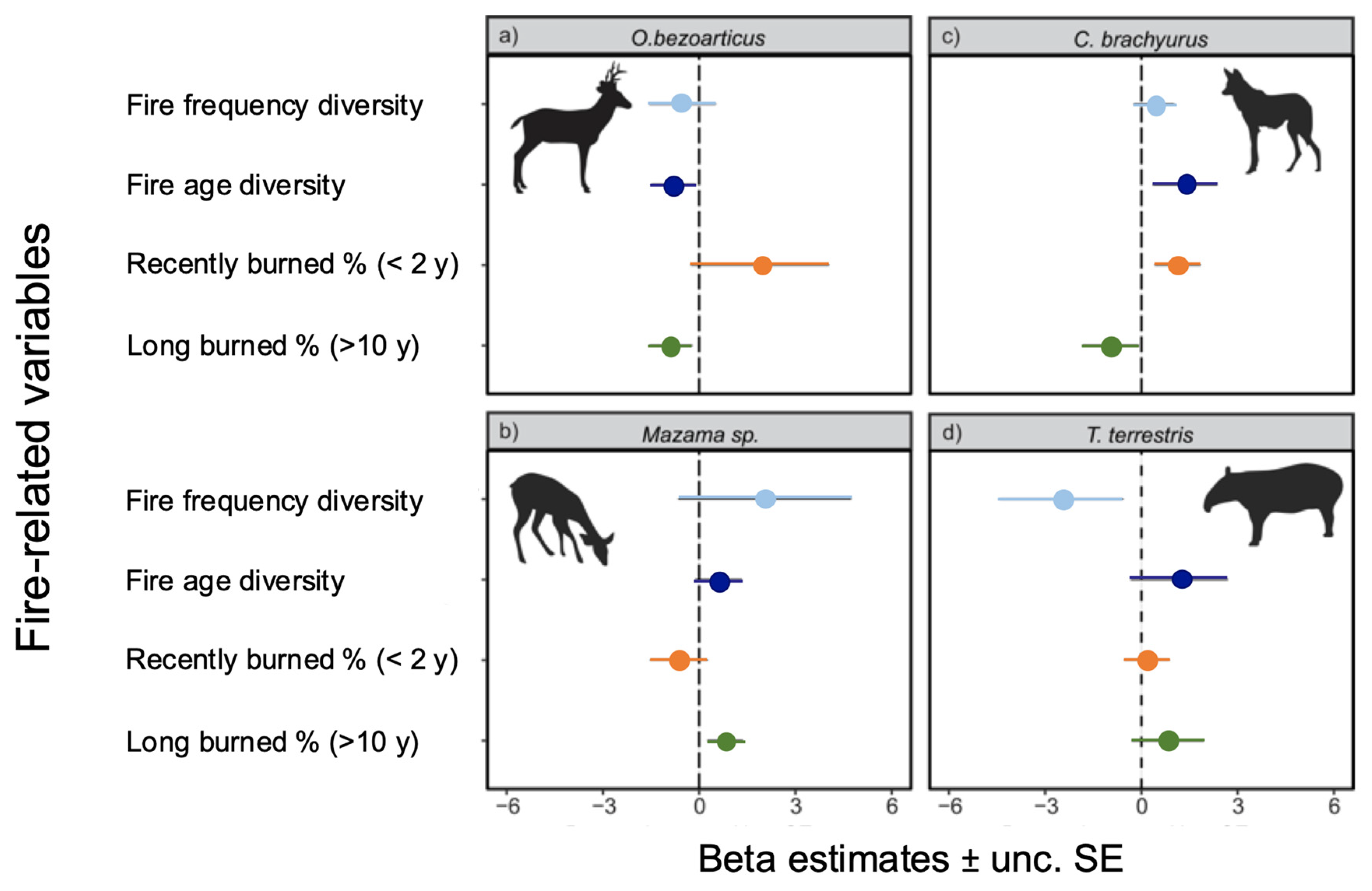
3. Results
4. Discussion
4.1. Fire Regime and Mammal Occupancy
4.2. Spatial Scales
4.3. Implications for Fire Management
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, R.S.; Gentle, L.K.; Mgoola, W.O.; Stone, E.L.; Uzal, A.; Yarnell, R.W. Habitat structure and the presence of large carnivores shape the site use of an understudied small carnivore: Caracal ecology in a miombo woodland. Mamm. Res. 2023, 68, 113–120. [Google Scholar] [CrossRef]
- Morin, S.J.; Bowman, J.; Marrotte, R.R.; Fortin, M.J. Fine-scale habitat selection by sympatric Canada lynx and bobcat. Ecol. Evol. 2020, 10, 9396–9409. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Carbone, C.; Wearn, O.R.; Rowcliffe, J.M.; Espinosa, S.; Lima, M.G.M.; Ahumada, J.A.; Gonçalves, A.L.S.; Trevelin, L.C.; Alvarez-Loayza, P.; et al. Prey availability and temporal partitioning modulate felid coexistence in Neotropical forests. PLoS ONE 2019, 14, e0213671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, L.N.; Davis, C.L.; Farris, Z.J.; Miller, D.A.W.; Tucker, J.M.; Hamel, S.; Farhadinia, M.S.; Steenweg, R.; Di Bitetti, M.S.; Thapa, K. Assessing global patterns in mammalian carnivore occupancy and richness by integrating local camera trap surveys. Glob. Ecol. Biogeogr. 2017, 26, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Regolin, A.L.; Oliveira-Santos, L.G.; Ribeiro, M.C.; Bailey, L.L. Habitat quality, not habitat amount, drives mammalian habitat use in the Brazilian Pantanal. Landsc. Ecol. 2021, 36, 2519–2533. [Google Scholar] [CrossRef]
- Apolloni, N.; Grüebler, M.U.; Arlettaz, R.; Gottschalk, T.K.; Naef-Daenzer, B. Habitat selection and range use of little owls in relation to habitat patterns at three spatial scales. Anim. Conserv. 2018, 21, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Viejou, R.; Avgar, T.; Brown, G.S.; Patterson, B.R.; Reid, D.E.B.; Rodgers, A.R.; Shuter, J.; Thompson, I.D.; Fryxell, J.M. Woodland caribou habitat selection patterns in relation to predation risk and forage abundance depend on reproductive state. Ecol. Evol. 2018, 8, 5863–5872. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; White, P.S. Natural disturbance and patch dynamics: An introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 3–13. [Google Scholar]
- Sergio, F.; Blas, J.; Hiraldo, F. Animal responses to natural disturbance and climate extremes: A review. Glob. Planet. Chang. 2018, 161, 28–40. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef]
- Camargo, A.C.L.; Barrio, R.O.L.; Camargo, N.F.D.; Mendonça, A.F.; Ribeiro, J.F.; Rodrigues, C.M.F.; Vieira, E.M. Fire affects the occurrence of small mammals at distinct spatial scales in a neotropical savanna. Eur. J. Wildl. Res. 2018, 64, 63. [Google Scholar] [CrossRef]
- Barlow, J.; Peres, C.A. Effects of single and recurrent wildfires on fruit production and large vertebrate abundance in a central Amazonian forest. Biodivers Conserv. 2006, 15, 985–1012. [Google Scholar] [CrossRef]
- Poulos, H.M.; Freiburger, M.R.; Barton, A.M.; Taylor, A.H. Mixed-severity wildfire as a driver of vegetation change in an Arizona Madrean Sky Island System, USA. Fire 2021, 4, 78. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Kolden, C.A.; Abatzoglou, J.T.; Johnston, F.H.; van der Werf, G.R.; Flannigan, M. Vegetation fires in the Anthropocene. Nat. Rev. Earth Environ. 2020, 1, 500–515. [Google Scholar] [CrossRef]
- Iglesias, V.; Balch, J.K.; Travis, W.R. U.S. fires became larger, more frequent, and more widespread in the 2000s. Sci. Adv. 2022, 8, eabc0020. [Google Scholar] [CrossRef] [PubMed]
- Fidelis, A.; Alvarado, S.T.; Barradas, A.C.S.; Pivello, V.R. The year 2017: Megafires and management in the Cerrado. Fire 2018, 1, 49. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.L.; Nogueira, J.; de Souza, R.A.; Falleiro, R.M.; Schmidt, I.B.; Libonati, R. Prescribed burning reduces large, high-intensity wildfires and emissions in the Brazilian savanna. Fire 2021, 4, 56. [Google Scholar] [CrossRef]
- Kelly, L.T.; Brotons, L.; McCarthy, M.A. Putting pyrodiversity to work for animal conservation. Conserv. Biol. 2016, 31, 952–955. [Google Scholar] [CrossRef]
- Taylor, R.S.; Watson, S.J.; Bennett, A.F.; Clarke, M.F. Which fire management strategies benefit biodiversity? A landscape-perspective case study using birds in mallee ecosystems of south-eastern Australia. Biol. Conserv. 2013, 159, 248–256. [Google Scholar] [CrossRef]
- Brockett, B.; Biggs, H.; van WILGEN, B.W. A patch mosaic burning system for conservation areas in southern African savannas. Int. J. Wildland Fire 2001, 10, 169–183. [Google Scholar] [CrossRef]
- Parr, C.L.; Andersen, A.N. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv. Biol. 2006, 20, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.J.; Taylor, R.S.; Spence-Bailey, L.; Nimmo, D.G.; Kenny, S.; Kelly, L.T.; Haslem, A.; Griffioen, P.; Callister, K.; Brown, L. The Mallee fire and biodiversity project. Proc. R. Soc. Vic. 2012, 124, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.E.; Sapsis, D.B. Fires as Agents of Biodiversity: Pyrodiversity Promotes Biodiversity. In Proceedings of the Symposium on Biodiversity of Northwestern California, Santa Rosa, CA, USA, 28–30 October 1991. [Google Scholar]
- Bird, R.B.; Bird, D.W.; Fernandez, L.E.; Taylor, N.; Taylor, W.; Nimmo, D. Aboriginal burning promotes fine-scale pyrodiversity and native predators in Australia’s Western Desert. Biol. Conserv. 2018, 219, 110–118. [Google Scholar] [CrossRef]
- Radford, I.J.; Woolley, L.-A.; Corey, B.; Vigilante, T.; Hatherley, E.; Fairman, R.; Carnes, K.; Start, A.N.; Wunambal Gaambera Aboriginal, C. Prescribed burning benefits threatened mammals in northern Australia. Biodivers. Conserv. 2020, 29, 2985–3007. [Google Scholar] [CrossRef]
- Jorge, M.H.; Conner, L.M.; Garrison, E.P.; Cherry, M.J. Avian species richness in a frequently burned ecosystem: A link between pyrodiversity and biodiversity. Landsc. Ecol. 2022, 37, 983–996. [Google Scholar] [CrossRef]
- Jones, G.M.; Tingley, M.W. Pyrodiversity and biodiversity: A history, synthesis, and outlook. Divers. Distrib. 2022, 28, 386–403. [Google Scholar] [CrossRef]
- Nimmo, D.G.; Kelly, L.T.; Spence-Bailey, L.M.; Watson, S.J.; Taylor, R.S.; Clarke, M.F.; Bennett, A.F. Fire mosaics and reptile conservation in a fire-prone region. Conserv. Biol. 2013, 27, 345–353. [Google Scholar] [CrossRef]
- Jorge, M.H.; Garrison, E.P.; Conner, L.M.; Cherry, M.J. Fire and land cover drive predator abundances in a pyric landscape. For. Ecol. Manag. 2020, 461, 117939. [Google Scholar] [CrossRef]
- Docherty, T.D.S.; Hethcoat, M.G.; MacTavish, L.M.; MacTavish, D.; Dell, S.; Stephens, P.A.; Willis, S.G. Burning savanna for avian species richness and functional diversity. Ecol. Appl. 2020, 30, e02091. [Google Scholar] [CrossRef]
- Taylor, R.S.; Watson, S.J.; Nimmo, D.G.; Kelly, L.T.; Bennett, A.F.; Clarke, M.F. Landscape-scale effects of fire on bird assemblages: Does pyrodiversity beget biodiversity? Divers. Distrib. 2012, 18, 519–529. [Google Scholar] [CrossRef]
- Chia, E.K.; Bassett, M.; Leonard, S.W.J.; Holland, G.J.; Ritchie, E.G.; Clarke, M.F.; Bennett, A.F. Effects of the fire regime on mammal occurrence after wildfire: Site effects vs landscape context in fire-prone forests. For. Ecol. Manag. 2016, 363, 130–139. [Google Scholar] [CrossRef]
- Cherry, M.J.; Chandler, R.B.; Garrison, E.P.; Crawford, D.A.; Kelly, B.D.; Shindle, D.B.; Godsea, K.G.; Miller, K.V.; Conner, L.M. Wildfire affects space use and movement of white-tailed deer in a tropical pyric landscape. For. Ecol. Manag. 2018, 409, 161–169. [Google Scholar] [CrossRef]
- Lewis, J.S.; LeSueur, L.; Oakleaf, J.; Rubin, E.S. Mixed-severity wildfire shapes habitat use of large herbivores and carnivores. For. Ecol. Manag. 2022, 506, 119933. [Google Scholar] [CrossRef]
- Geary, W.L.; Doherty, T.S.; Nimmo, D.G.; Tulloch, A.I.T.; Ritchie, E.G. Predator responses to fire: A global systematic review and meta-analysis. J. Anim. Ecol. 2020, 89, 955–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eby, S.; Mosser, A.; Swanson, A.; Packer, C.; Ritchie, M. The impact of burning on lion Panthera leo habitat choice in an African savanna. Curr. Zool. 2013, 59, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Doherty, T.S.; Geary, W.L.; Jolly, C.J.; Macdonald, K.J.; Miritis, V.; Watchorn, D.J.; Cherry, M.J.; Conner, L.M.; González, T.M.; Legge, S.M. Fire as a driver and mediator of predator–prey interactions. Biol. Rev 2022, 97, 1539–1558. [Google Scholar] [CrossRef]
- Cherry, M.J.; Warren, R.J.; Conner, L.M. Fire-mediated foraging tradeoffs in white-tailed deer. Ecosphere 2017, 8, e01784. [Google Scholar] [CrossRef]
- Geary, W.L.; Ritchie, E.G.; Lawton, J.A.; Healey, T.R.; Nimmo, D.G. Incorporating disturbance into trophic ecology: Fire history shapes mesopredator suppression by an apex predator. J. Appl. Ecol. 2018, 55, 1594–1603. [Google Scholar] [CrossRef]
- Eby, S.L.; Anderson, T.M.; Mayemba, E.P.; Ritchie, M.E. The effect of fire on habitat selection of mammalian herbivores: The role of body size and vegetation characteristics. J. Anim. Ecol. 2014, 83, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Nieman, W.A.; van Wilgen, B.W.; Radloff, F.G.T.; Leslie, A.J. A review of the responses of medium-to large-sized African mammals to fire. Afr. J. Range Forage Sci. 2022, 39, 249–263. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Burns, C.E.; Tambling, C.J.; Amendola, E.; Buis, G.M.; Govender, N.; Nelson, V.; Thompson, D.I.; Zinn, A.D.; Smith, M.D. Habitat selection by large herbivores in a southern African savanna: The relative roles of bottom-up and top-down forces. Ecosphere 2013, 4, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Reid, A.M.; Murphy, B.P.; Vigilante, T.; Corporation, W.G.A.; Bowman, D.M. Distribution and abundance of large herbivores in a northern Australian tropical savanna: A multi-scale approach. Austral Ecol. 2020, 45, 529–547. [Google Scholar] [CrossRef]
- Delaney, L.; Di Stefano, J.; Sitters, H. Mammal responses to spatial pattern in fire history depend on landscape context. Landsc. Ecol. 2021, 36, 897–914. [Google Scholar] [CrossRef]
- Prowse, T.A.A.; Collard, S.J.; Blackwood, A.; O’Connor, P.J.; Delean, S.; Barnes, M.; Cassey, P.; Possingham, H.P. Prescribed burning impacts avian diversity and disadvantages woodland-specialist birds unless long-unburnt habitat is retained. Biol. Conserv. 2017, 215, 268–276. [Google Scholar] [CrossRef]
- Takach, B.V.; Jolly, C.J.; Dixon, K.M.; Penton, C.E.; Doherty, T.S.; Banks, S.C. Long-unburnt habitat is critical for the conservation of threatened vertebrates across Australia. Landsc. Ecol. 2022, 37, 1469–1482. [Google Scholar] [CrossRef]
- Stillman, A.N.; Siegel, R.B.; Wilkerson, R.L.; Johnson, M.; Tingley, M.W. Age-dependent habitat relationships of a burned forest specialist emphasise the role of pyrodiversity in fire management. J. Appl. Ecol. 2019, 56, 880–890. [Google Scholar] [CrossRef]
- Radford, I.J.; Corey, B.; Carnes, K.; Shedley, E.; McCaw, L.; Woolley, L.-A. Landscape-scale effects of fire, cats, and feral livestock on threatened savanna mammals: Unburnt habitat matters more than pyrodiversity. Front. Ecol. Evol. 2021, 9, 739817. [Google Scholar] [CrossRef]
- Beale, C.M.; Courtney Mustaphi, C.J.; Morrison, T.A.; Archibald, S.; Anderson, T.M.; Dobson, A.P.; Donaldson, J.E.; Hempson, G.P.; Probert, J.; Parr, C.L. Pyrodiversity interacts with rainfall to increase bird and mammal richness in African savannas. Ecol. Lett. 2018, 21, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Blanchard, G.; Ibanez, T.; Munoz, F.; Bruy, D.; Hely, C.; Munzinger, J.; Birnbaum, P. Drivers of tree community assembly during tropical forest post-fire succession in anthropogenic savannas. Perspect. Plant Ecol. Evol. Syst. 2021, 52, 125630. [Google Scholar] [CrossRef]
- Zylinski, S.; Swan, M.; Sitters, H. Interrelationships between fire, habitat, and mammals in a fragmented heathy woodland. For. Ecol. Manag. 2022, 522, 120464. [Google Scholar] [CrossRef]
- Nimmo, D.G.; Avitabile, S.; Banks, S.C.; Bliege Bird, R.; Callister, K.; Clarke, M.F.; Dickman, C.R.; Doherty, T.S.; Driscoll, D.A.; Greenville, A.C. Animal movements in fire-prone landscapes. Biol. Rev. 2019, 94, 981–998. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, M.R.D.; Marcuzzo, F.F.N.; Barros, J.R. Classificação climática de Köppen-Geiger para o estado de Goiás e o Distrito Federal. ACTA Geográfica 2014, 8, 40–55. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Walter, B.M.T. Fitofisionomias do bioma cerrado. In Cerrado: Ambiente e Flora; Sano, S.M., Almeida, S.P., Eds.; Embrapa CPAC: Planaltina, Brazil, 1998; pp. 89–166. [Google Scholar]
- Lewis, K.; Barros, F.V.; Cure, M.B.; Davies, C.A.; Furtado, M.N.; Hill, T.C.; Hirota, M.; Martins, D.L.; Mazzochini, G.G.; Mitchard, E.T.A. Mapping native and non-native vegetation in the Brazilian Cerrado using freely available satellite products. Sci. Rep. 2022, 12, 1588. [Google Scholar] [CrossRef]
- Miranda, H.S.; Sato, M.N.; Neto, W.N.; Aires, F.S. Fires in the cerrado, the Brazilian savanna. In Tropical Fire Ecology: Climate Change, Land Use, and Ecosystem Dynamics; Cochrane, M.A., Ed.; Springer: Berlin, Germany, 2009; pp. 427–450. [Google Scholar]
- MapBiomas, MapBiomas Project-Collection 6 of the Annual Series of Land Use and Land Cover Maps of Brazil. Available online: https://mapbiomas.org/ (accessed on 15 October 2021).
- Schmidt, I.B.; Moura, L.C.; Ferreira, M.C.; Eloy, L.; Sampaio, A.B.; Dias, P.A.; Berlinck, C.N. Fire management in the Brazilian savanna: First steps and the way forward. J. Appl. Ecol. 2018, 55, 2094–2101. [Google Scholar] [CrossRef] [Green Version]
- Pivello, V.R.; Vieira, I.; Christianini, A.V.; Ribeiro, D.B.; da Silva Menezes, L.; Berlinck, C.N.; Melo, F.P.L.; Marengo, J.A.; Tornquist, C.G.; Tomas, W.M. Understanding Brazil’s catastrophic fires: Causes, consequences and policy needed to prevent future tragedies. Perspect. Ecol. Conserv. 2021, 19, 233–255. [Google Scholar] [CrossRef]
- González, S.; Cosse, M.; Góss Braga, F.; Vila, A.R.; Merino, M.L.; Dellafiore, C.; Cartes, J.L.; Maffei, L.; Gimenez Dixon, M. Pampas deer Ozotoceros bezoarticus (Linnaeus 1758). In Neotropical Cervidology: Biology and Medicine of Latin American Deer; Duarte, J.M.B., González, S., Eds.; Funep/IUCN: Jaboticabal, Brazil, 2010; pp. 119–132. [Google Scholar]
- Vila, A.R.; Beade, M.S.; Barrios Lamunière, D. Home range and habitat selection of pampas deer. J. Zool. 2008, 276, 95–102. [Google Scholar] [CrossRef]
- Rivero, K.; Rumiz, D.I.; Taber, A.B. Differential habitat use by two sympatric brocket deer species (Mazama americana and M. gouazoubira) in a seasonal Chiquitano forest of Bolivia. Mammalia 2005, 69, 169–183. [Google Scholar] [CrossRef]
- Black-Décima, P.; Rossi, R.V.; Vogliotti, A.; Cartes, J.L.; Maffei, L.; Duarte, J.M.B.; González, S.; Juliá, J.P. Brown brocket deer Mazama gouazoubira (Fischer 1814). In Neotropical Cervidology: Biology and medicine of Latin American deer; Duarte, J.M.B., González, S., Eds.; Funep/IUCN: Jaboticabal, Brazil, 2010; pp. 190–201. [Google Scholar]
- Di Bitetti, M.S.; Iezzi, M.E.; Cruz, P.; Varela, D.; De Angelo, C. Effects of cattle on habitat use and diel activity of large native herbivores in a South American rangeland. J. Nat. Conserv. 2020, 58, 125900. [Google Scholar] [CrossRef]
- Varela, D.M.; Trovati, R.G.; Guzmán, K.R.; Rossi, R.V.; Duarte, J.M.B.; Duarte, J. Red brocket deer Mazama americana (Erxleben 1777). In Neotropical Cervidology, Biology and Medicine of Latin American Deer; Duarte, J.M.B., González, S., Eds.; Funep/IUCN: Jaboticabal, Brazil, 2010; pp. 151–159. [Google Scholar]
- Padilla, M.; Dowler, R.C. Tapirus terrestris. Mamm. Species 1994, 481, 1–8. [Google Scholar] [CrossRef]
- Medici, E.P.; Mezzini, S.; Fleming, C.H.; Calabrese, J.M.; Noonan, M.J. Movement ecology of vulnerable lowland tapirs between areas of varying human disturbance. Mov. Ecol. 2022, 10, 14. [Google Scholar] [CrossRef]
- Wilman, H.; Belmaker, J.; Simpson, J.; de la Rosa, C.; Rivadeneira, M.M.; Jetz, W. EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar] [CrossRef] [Green Version]
- Giordano, C.; Lyra-Jorge, M.C.; Miotto, R.A.; Pivello, V.R. Food habits of three carnivores in a mosaic landscape of São Paulo state, Brazil. Eur. J. Wildl. Res. 2018, 64, 15. [Google Scholar] [CrossRef]
- Jácomo, A.T.A.; Silveira, L.; Diniz-Filho, J.A.F. Niche separation between the maned wolf (Chrysocyon brachyurus), the crab-eating fox (Dusicyon thous) and the hoary fox (Dusicyon vetulus) in central Brazil. J. Zool. 2004, 262, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Prada, M. Effects of fire on the abundance of large mammalian herbivores in Mato Grosso, Brazil. Mammalia 2001, 65, 55–62. [Google Scholar] [CrossRef]
- Nunes, B.d.A. Impactos de um Grande Incêndio na Comunidade de Mamíferos de Médio e Grande Porte em Uma Área de Cerrado no Brasil Central. Master’s Dissertation, University of Brasilia, Brasilia, Brazil, 2022. [Google Scholar]
- Meek, P.D.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’brien, T.; O’connell, A.; Sanderson, J.; Swann, D.E.; Tobler, M. Recommended guiding principles for reporting on camera trapping research. Biodivers. Conserv. 2014, 23, 2321–2343. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Brennan, J.M.; Bender, D.J.; Contreras, T.A.; Fahrig, L. Focal patch landscape studies for wildlife management: Optimizing sampling effort. In Integrating Landscape Ecology into Natural Resource Management; Liu, J., Taylor, W.W., Eds.; Cambridge University Press: Cambridge, UK, 2002; p. 68. [Google Scholar]
- QGIS. QGIS Geographic Information System. Available online: http://www.qgis.org (accessed on 15 October 2021).
- Pilon, N.A.L.; Cava, M.G.B.; Hoffmann, W.A.; Abreu, R.C.R.; Fidelis, A.; Durigan, G. The diversity of post-fire regeneration strategies in the cerrado ground layer. J. Ecol. 2021, 109, 154–166. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Huete, A.R. Assessing the seasonal dynamics of the Brazilian Cerrado vegetation through the use of spectral vegetation indices. Int. J. Remote Sens. 2004, 25, 1837–1860. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Royle, J.A. Designing occupancy studies: General advice and allocating survey effort. J. Appl. Ecol. 2005, 42, 1105–1114. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Andrew Royle, J.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Mackenzie, D.I. Modeling the probability of resource use: The effect of, and dealing with, detecting a species imperfectly. J. Wildl. Manag. 2006, 70, 367–374. [Google Scholar] [CrossRef]
- Rovero, F.; Martin, E.; Rosa, M.; Ahumada, J.A.; Spitale, D. Estimating species richness and modelling habitat preferences of tropical forest mammals from camera trap data. PLoS ONE 2014, 9, e103300. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Reis, M.B.; Nichols, J.D.; Chiarello, A.G.; Ribeiro, M.C.; Setz, E.Z. Landscape use and co-occurrence patterns of Neotropical spotted cats. PLoS ONE 2017, 12, e0168441. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, J.D.; Arkle, R.S.; Rachlow, J.L.; Vierling, K.T.; Pilliod, D.S.; Wiest, M.M. Occupancy and abundance of predator and prey: Implications of the fire-cheatgrass cycle in sagebrush ecosystems. Ecosphere 2016, 7, e01307. [Google Scholar] [CrossRef] [Green Version]
- Gould, M.J.; Gould, W.R.; Cain, J.W., III; Roemer, G.W. Validating the performance of occupancy models for estimating habitat use and predicting the distribution of highly-mobile species: A case study using the American black bear. Biol. Conserv. 2019, 234, 28–36. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. A Practical Information-Theoretic Approach: Model Selection and Multimodel Inference, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- McHugh, D.; Goldingay, R.L.; Letnic, M. Occupancy and co-occurrence patterns of endemic mammals and introduced predators across a broad geographical gradient in eastern Australia. Biodivers. Conserv. 2022, 31, 989–1021. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2004, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 10 January 2021).
- Fiske, I.; Chandler, R. unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Venter, J.A.; Nabe-Nielsen, J.; Prins, H.H.; Slotow, R. Forage patch use by grazing herbivores in a South African grazing ecosystem. Acta Theriol. 2014, 59, 457–466. [Google Scholar] [CrossRef]
- Ganz, T.R.; DeVivo, M.T.; Kertson, B.N.; Roussin, T.; Satterfield, L.; Wirsing, A.J.; Prugh, L.R. Interactive effects of wildfires, season and predator activity shape mule deer movements. J. Anim. Ecol. 2022, 91, 2273–2288. [Google Scholar] [CrossRef] [PubMed]
- Reis, N.R.; Peracchi, A.L.; Pedro, W.A.; Lima, I.P. Mamíferos do Brasil. In Mamíferos do Brasil; Universidade Estadual de Londrina: Londrina, Brazil, 2006; p. 437. [Google Scholar]
- Azevedo, F.C.C. Food habits and livestock depredation of sympatric jaguars and pumas in the Iguacu National Park area, south Brazil. Biotropica 2008, 40, 494–500. [Google Scholar] [CrossRef]
- Foster, R.J.; Harmsen, B.J.; Valdes, B.; Pomilla, C.; Doncaster, C. Food habits of sympatric jaguars and pumas across a gradient of human disturbance. J. Zool. 2010, 280, 309–318. [Google Scholar] [CrossRef]
- Cosse, M.; González, S.; Gimenez-Dixon, M. Feeding ecology of Ozotoceros bezoarticus: Conservation implicationsin Uruguay. Iheringia. Série Zool. 2009, 99, 158–164. [Google Scholar] [CrossRef]
- Desbiez, A.L.J.; Santos, S.A.; Alvarez, J.M.; Tomas, W.M. Forage use in domestic cattle (Bos indicus), capybara (Hydrochoerus hydrochaeris) and pampas deer (Ozotoceros bezoarticus) in a seasonal Neotropical wetland. Mamm. Biol. 2011, 76, 351–357. [Google Scholar] [CrossRef]
- Prado, H.M. Feeding ecology of five Neotropical ungulates: A critical review. Oecol. Aust. 2013, 17, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, L.; Somers, M.J.; Dalerum, F. Effects of prescribed burning and mechanical bush clearing on ungulate space use in an African savannah. Restor. Ecol. 2013, 21, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Klop, E.; Goethem, J. Savanna fires govern community structure of ungulates in Bénoué National Park, Cameroon. J. Trop. Ecol. 2008, 24, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Kelly, L.T.; Nimmo, D.G.; Spence-Bailey, L.M.; Taylor, R.S.; Watson, S.J.; Clarke, M.F.; Bennett, A.F. Managing fire mosaics for small mammal conservation: A landscape perspective. J. Appl. Ecol. 2012, 49, 412–421. [Google Scholar] [CrossRef]
- Steel, Z.L.; Campos, B.; Frick, W.F.; Burnett, R.; Safford, H.D. The effects of wildfire severity and pyrodiversity on bat occupancy and diversity in fire-suppressed forests. Sci. Rep. 2019, 9, 16300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobler, M.W.; Carrillo-Percastegui, S.E.; Powell, G. Habitat use, activity patterns and use of mineral licks by five species of ungulate in south-eastern Peru. J. Trop. Ecol. 2009, 25, 261–270. [Google Scholar] [CrossRef]
- Ferreguetti, Á.C.; Tomás, W.M.; Bergallo, H.G. Density, occupancy, and detectability of lowland tapirs, Tapirus terrestris, in Vale Natural Reserve, southeastern Brazil. J. Mammal. 2017, 98, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Vélez, J.; Espelta, J.M.; Rivera, O.; Armenteras, D. Effects of seasonality and habitat on the browsing and frugivory preferences of Tapirus terrestris in north-western Amazonia. J. Trop. Ecol. 2017, 33, 395–406. [Google Scholar] [CrossRef]
- Villalobos, M.P.; Bagno, M. Frugivorous mammals and the buriti palm, Mauritia flexuosa (Arecaceae), in Central Brazil. Brenesia 2011, 75–76, 109–111. [Google Scholar]
- Kuhlmann, M.; Ribeiro, J.F. Fruits and frugivores of the Brazilian Cerrado: Ecological and phylogenetic considerations. Acta Bot Bras. 2016, 30, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Le Stradic, S.; Hernandez, P.; Fernandes, G.W.; Buisson, E. Regeneration after fire in campo rupestre: Short- and long-term vegetation dynamics. Flora 2018, 238, 191–200. [Google Scholar] [CrossRef]
- Gomes, L.; Maracahipes, L.; Marimon, B.S.; Reis, S.M.; Elias, F.; Maracahipes-Santos, L.; Marimon-Junior, B.H.; Lenza, E. Post-fire recovery of savanna vegetation from rocky outcrops. Flora Morphol. Distrib. Funct. Ecol. 2014, 209, 201–208. [Google Scholar] [CrossRef]
- Lino, S.; Sillero, N.; Torres, J.; Santos, X.; Álvares, F. The role of fire on wolf distribution and breeding-site selection: Insights from a generalist carnivore occurring in a fire-prone landscape. Landsc Urban Plan. 2019, 183, 111–121. [Google Scholar] [CrossRef]
- Green, D.S.; Roloff, G.J.; Heath, B.R.; Holekamp, K.E. Temporal dynamics of the reponses by African mammals to prescribed fire. J. Wildl. Manag. 2015, 79, 235–242. [Google Scholar] [CrossRef]
- Silva, B.R.; Root-Bernstein, M. History of canids in Chile and impacts on prey adaptations. Ecol. Evol. 2021, 11, 9892–9903. [Google Scholar] [CrossRef]
- Queirolo, D.; Motta-Junior, J.C. Prey availability and diet of maned wolf in Serra da Canastra National Park, southeastern Brazil. Acta Theriol. 2007, 52, 391–402. [Google Scholar] [CrossRef]
- Rodrigues, F.H.; Hass, A.; Lacerda, A.C.; Grando, R.L.; Bagno, M.A.; Bezerra, A.M.; Silva, W.R. Feeding habits of the maned wolf (Chrysocyon brachyurus) in the Brazilian Cerrado. Mastozool. Neotrop. 2007, 14, 37–51. [Google Scholar]
- Briani, D.C.; Palma, A.R.T.; Vieira, E.M.; Henriques, R.P.B. Post-fire succession of small mammals in the Cerrado of central Brazil. Biodivers. Conserv. 2004, 13, 1023–1037. [Google Scholar] [CrossRef]
- Leahy, L.; Legge, S.M.; Tuft, K.; McGregor, H.W.; Barmuta, L.A.; Jones, M.E.; Johnson, C.N. Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas. Wildl. Res. 2016, 42, 705–716. [Google Scholar] [CrossRef]
- Tingley, M.W.; Ruiz-Gutiérrez, V.; Wilkerson, R.L.; Howell, C.A.; Siegel, R.B. Pyrodiversity promotes avian diversity over the decade following forest fire. Proc. R. Soc. B 2016, 283, 20161703. [Google Scholar] [CrossRef] [Green Version]
- Pastro, L.A.; Dickman, C.R.; Letnic, M. Burning for biodiversity or burning biodiversity? Prescribed burn vs. wildfire impacts on plants, lizards, and mammals. Ecol. Appl. 2011, 21, 3238–3253. [Google Scholar] [CrossRef]
- Burgess, E.E.; Maron, M. Does the response of bird assemblages to fire mosaic properties vary among spatial scales and foraging guilds? Landsc. Ecol. 2016, 31, 687–699. [Google Scholar] [CrossRef]
- Kelly, L.T.; Bennett, A.F.; Clarke, M.F.; McCarthy, M.A. Optimal fire histories for biodiversity conservation. Conserv. Biol. 2015, 29, 473–481. [Google Scholar] [CrossRef]
- Jones, G.M.; Kramer, H.A.; Whitmore, S.A.; Berigan, W.J.; Tempel, D.J.; Wood, C.M.; Hobart, B.K.; Erker, T.; Atuo, F.A.; Pietrunti, N.F. Habitat selection by spotted owls after a megafire reflects their adaptation to historical frequent-fire regimes. Landsc. Ecol. 2020, 35, 1199–1213. [Google Scholar] [CrossRef]
- York, A.; Lewis, T. Understanding the effects of fire on invertebrates in Australian temperate and sub-tropical forests: The value of long-term experiments. Aust. Zool. 2018, 39, 633–645. [Google Scholar] [CrossRef]
- Stillman, A.N.; Lorenz, T.J.; Fischer, P.C.; Siegel, R.B.; Wilkerson, R.L.; Johnson, M.; Tingley, M.W. Juvenile survival of a burned forest specialist in response to variation in fire characteristics. J. Anim. Ecol. 2021, 90, 1317–1327. [Google Scholar] [CrossRef]
- Jácomo, A.T.A.; Kashivakura, C.K.; Ferro, C.; Furtado, M.M.; Astete, S.P.; Tôrres, N.M.; Sollmann, R.; Silveira, L. Home range and spatial organization of maned wolves in the Brazilian grasslands. J. Mammal. 2009, 90, 150–157. [Google Scholar] [CrossRef]
- Coelho, C.M.; De Melo, L.F.B.; Sábato, M.A.L.; Vaz Magni, E.M.; Hirsch, A.; Young, R.J. Habitat use by wild maned wolves (Chrysocyon brachyurus) in a transition zone environment. J. Mammal. 2008, 89, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Juarez, K.M.; Marinho-Filho, J. Diet, habitat use, and home ranges of sympatric Canids in central Brazil. J. Mammal. 2002, 83, 925–933. [Google Scholar] [CrossRef]
- Emmons, L.H. The Maned Wolves of Noel Kempff Mercado National Park; Smithsonian Institution: Washington, DC, USA, 2012. [Google Scholar]
- Ferreira, P.M.A.; Ely, C.V.; Beal-Neves, M. Different post-fire stages encompass different plant community compositions in fire-prone grasslands from Southern Brazil. Flora 2021, 285, 151937. [Google Scholar] [CrossRef]
- Hale, S.; Mendoza, L.; Yeatman, T.; Cooke, R.; Doherty, T.; Nimmo, D.; White, J.G. Evidence that post-fire recovery of small mammals occurs primarily via in situ survival. Divers. Distrib. 2022, 28, 404–416. [Google Scholar] [CrossRef]
- McGregor, H.W.; Legge, S.; Jones, M.E.; Johnson, C.N. Landscape management of fire and grazing regimes alters the fine-scale habitat utilisation by feral cats. PLoS ONE 2014, 9, e109097. [Google Scholar] [CrossRef] [Green Version]
- Mayor, S.J.; Schneider, D.C.; Schaefer, J.A.; Mahoney, S.P. Habitat selection at multiple scales. Ecoscience 2009, 16, 238–247. [Google Scholar] [CrossRef]
- Holland, J.D.; Yang, S. Multi-scale studies and the ecological neighborhood. Curr. Landsc. Ecol. Rep. 2016, 1, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Jackson, H.B.; Fahrig, L. What size is a biologically relevant landscape? Landsc. Ecol. 2012, 27, 929–941. [Google Scholar] [CrossRef]
- Bowman, J.; Jaeger, J.A.; Fahrig, L. Dispersal distance of mammals is proportional to home range size. Ecology 2002, 83, 2049–2055. [Google Scholar] [CrossRef]
- Wilgen, B.W.V.; Forsyth, G.G. Towards adaptive fire management for biodiversity conservation: Experience in South African National Parks. Koedoe Afr. Prot. Area Conserv Sci 2011, 53, a982. [Google Scholar] [CrossRef]
- Andersen, A.N. Faunal responses to fire in Australian tropical savannas: Insights from field experiments and their lessons for conservation management. Divers. Distrib. 2021, 27, 828–843. [Google Scholar] [CrossRef]
- Farnsworth, L.M.; Nimmo, D.G.; Kelly, L.T.; Bennett, A.F.; Clarke, M.F. Does pyrodiversity beget alpha, beta or gamma diversity? A case study using reptiles from semi-arid Australia. Divers. Distrib. 2014, 20, 663–673. [Google Scholar] [CrossRef]
- Doumas, S.L.; Koprowski, J.L. Effect of heterogeneity in burn severity on Mexican fox squirrels following the return of fire. Int. J. Wildland Fire 2012, 22, 405–413. [Google Scholar] [CrossRef]
- Furnas, B.J.; Goldstein, B.R.; Figura, P.J. Intermediate fire severity diversity promotes richness of forest carnivores in California. Divers. Distrib. 2022, 28, 493–505. [Google Scholar] [CrossRef]
- Jones, G.M.; Ayars, J.; Parks, S.A.; Chmura, H.E.; Cushman, S.A.; Sanderlin, J.S. Pyrodiversity in a warming world: Research challenges and opportunities. Curr. Landsc. Ecol. Rep. 2022, 1–19. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Type | Multiple Spatial Scales | Description |
|---|---|---|---|
| NDVI | Continuous | Yes | Normalized Difference Vegetation Index indicating the vegetation cover. |
| season | Categorical | No | Temporal variable representing the two seasons according to mensal precipitation: wet season and dry season. |
| method | Categorical | No | Variable representing the methods used during the sampling period: photo and video. |
| div-freq | Continuous | Yes | Fire frequency diversity estimated using the exponential of Shannon entropy. Div-freq values represent the number of effective patches with different fire frequencies needed to obtain the diversity index value. |
| div-age | Continuous | Yes | Fire age diversity estimated using the exponential of Shannon’s diversity index. Div-age values represent the number of effective patches with different fire ages needed to obtain the diversity index value. |
| burned < 2 | Continuous | Yes | Proportion of area burned in the two years before the mammal sampling. Proportion was calculated relative to the total area of the buffer around the camera. |
| unburned > 10 | Continuous | Yes | Proportion of area unburned in the previous 10 years before the mammal sampling. Proportion was calculated relative to the total area of the buffer around the camera. |
| Models | K | AICc | ΔAICc | AICcWt | Cum.Wt | 22LL |
|---|---|---|---|---|---|---|
| O. bezoarticus | ||||||
| ψ(.), p(ndvi) | 3 | 800.75 | 0 | 0.22 | 0.22 | −396.91 |
| ψ(unburned > 10), p(ndvi) | 4 | 801.62 | 0.87 | 0.14 | 0.36 | −396.01 |
| ψ(burned < 2), p(ndvi) | 4 | 801.74 | 1.00 | 0.13 | 0.49 | −396.07 |
| ψ(div-age), p(ndvi) | 4 | 802.27 | 1.52 | 0.1 | 0.59 | −396.33 |
| ψ(div-age + burned < 2), p(ndvi) | 5 | 802.58 | 1.83 | 0.09 | 0.68 | −395.04 |
| ψ(div-freq), p(ndvi) | 4 | 803.38 | 2.63 | 0.06 | 0.74 | −396.89 |
| ψ(div-freq + burned < 2), p(ndvi) | 5 | 803.43 | 2.68 | 0.06 | 0.8 | −395.46 |
| ψ(burned < 2 + unburned > 10), p(ndvi) | 5 | 803.5 | 2.75 | 0.05 | 0.85 | −395.5 |
| ψ(div-freq + unburned > 10), p(ndvi) | 5 | 804.3 | 3.55 | 0.04 | 0.89 | −395.9 |
| ψ(div-age + unburned > 10), p(ndvi) | 5 | 804.42 | 3.67 | 0.03 | 0.92 | −395.96 |
| Mazama sp. | ||||||
| ψ(.), p(ndvi) | 3 | 206.76 | 0 | 0.21 | 0.21 | −99.92 |
| ψ(unburned > 10), p(ndvi) | 4 | 207.03 | 0.27 | 0.18 | 0.39 | −98.72 |
| ψ(div-freq), p(ndvi) | 4 | 208.07 | 1.31 | 0.11 | 0.5 | −99.24 |
| ψ(div-last), p(ndvi) | 4 | 208.23 | 1.47 | 0.1 | 0.59 | −99.32 |
| ψ(div-freq + unburned > 10), p(ndvi) | 5 | 208.47 | 1.71 | 0.09 | 0.68 | −97.98 |
| ψ(burned < 2), p(ndvi) | 4 | 208.85 | 2.09 | 0.07 | 0.76 | −99.62 |
| ψ(div-freq + burned < 2), p(ndvi) | 5 | 209.58 | 2.81 | 0.05 | 0.81 | −98.54 |
| ψ(div-last + unburned > 10), p(ndvi) | 5 | 209.68 | 2.92 | 0.05 | 0.85 | −98.59 |
| ψ(burned < 2 + unburned > 10), p(ndvi) | 5 | 209.9 | 3.13 | 0.04 | 0.9 | −98.7 |
| ψ(div-last + burned < 2), p(ndvi) | 5 | 210.49 | 3.73 | 0.03 | 0.93 | −99 |
| C. brachyurus | ||||||
| ψ(burned < 2), p(ndvi) | 4 | 429.38 | 0 | 0.21 | 0.21 | −209.89 |
| ψ(div-last + burned < 2), p(ndvi) | 5 | 429.68 | 0.3 | 0.18 | 0.4 | −208.59 |
| ψ(div-last + unburned > 10), p(ndvi) | 5 | 430.24 | 0.86 | 0.14 | 0.54 | −208.87 |
| ψ(div-last + burned < 2 + unburned > 10), p(ndvi) | 6 | 430.94 | 1.56 | 0.1 | 0.63 | −207.64 |
| ψ(div-freq + burned < 2), p(ndvi) | 5 | 431.27 | 1.89 | 0.08 | 0.72 | −209.39 |
| ψ(div-last), p(ndvi) | 4 | 431.47 | 2.09 | 0.08 | 0.79 | −210.93 |
| ψ(burned < 2 + unburned > 10), p(ndvi) | 5 | 432.25 | 2.87 | 0.05 | 0.84 | −209.87 |
| ψ(div-freq + div-last + burned < 2), p(ndvi) | 6 | 432.78 | 3.4 | 0.04 | 0.88 | −208.56 |
| ψ(.), p(ndvi) | 3 | 432.97 | 3.59 | 0.04 | 0.92 | −213.02 |
| T. terrestris | ||||||
| ψ(div-freq), p(ndvi) | 4 | 222.77 | 0 | 0.41 | 0.41 | −106.59 |
| ψ(div-freq + div-last), p(ndvi) | 5 | 223.84 | 1.06 | 0.24 | 0.64 | −105.67 |
| ψ(div-freq + unburned > 10), p(ndvi) | 5 | 225 | 2.22 | 0.13 | 0.78 | −106.25 |
| ψ(div-freq + burned < 2), p(ndvi) | 5 | 225.61 | 2.84 | 0.1 | 0.88 | −106.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, C.V.; Lourenço, Á.; Vieira, E.M. Species-Specific Responses of Medium and Large Mammals to Fire Regime Attributes in a Fire-Prone Neotropical Savanna. Fire 2023, 6, 110. https://doi.org/10.3390/fire6030110
Souza CV, Lourenço Á, Vieira EM. Species-Specific Responses of Medium and Large Mammals to Fire Regime Attributes in a Fire-Prone Neotropical Savanna. Fire. 2023; 6(3):110. https://doi.org/10.3390/fire6030110
Chicago/Turabian StyleSouza, Clarice Vieira, Águeda Lourenço, and Emerson Monteiro Vieira. 2023. "Species-Specific Responses of Medium and Large Mammals to Fire Regime Attributes in a Fire-Prone Neotropical Savanna" Fire 6, no. 3: 110. https://doi.org/10.3390/fire6030110