
‘Pine Decline or pine declines?’ Analysis and Interpretation of Bog-Pines from Wem Moss, Shropshire, UK

Abstract
:1. Introduction
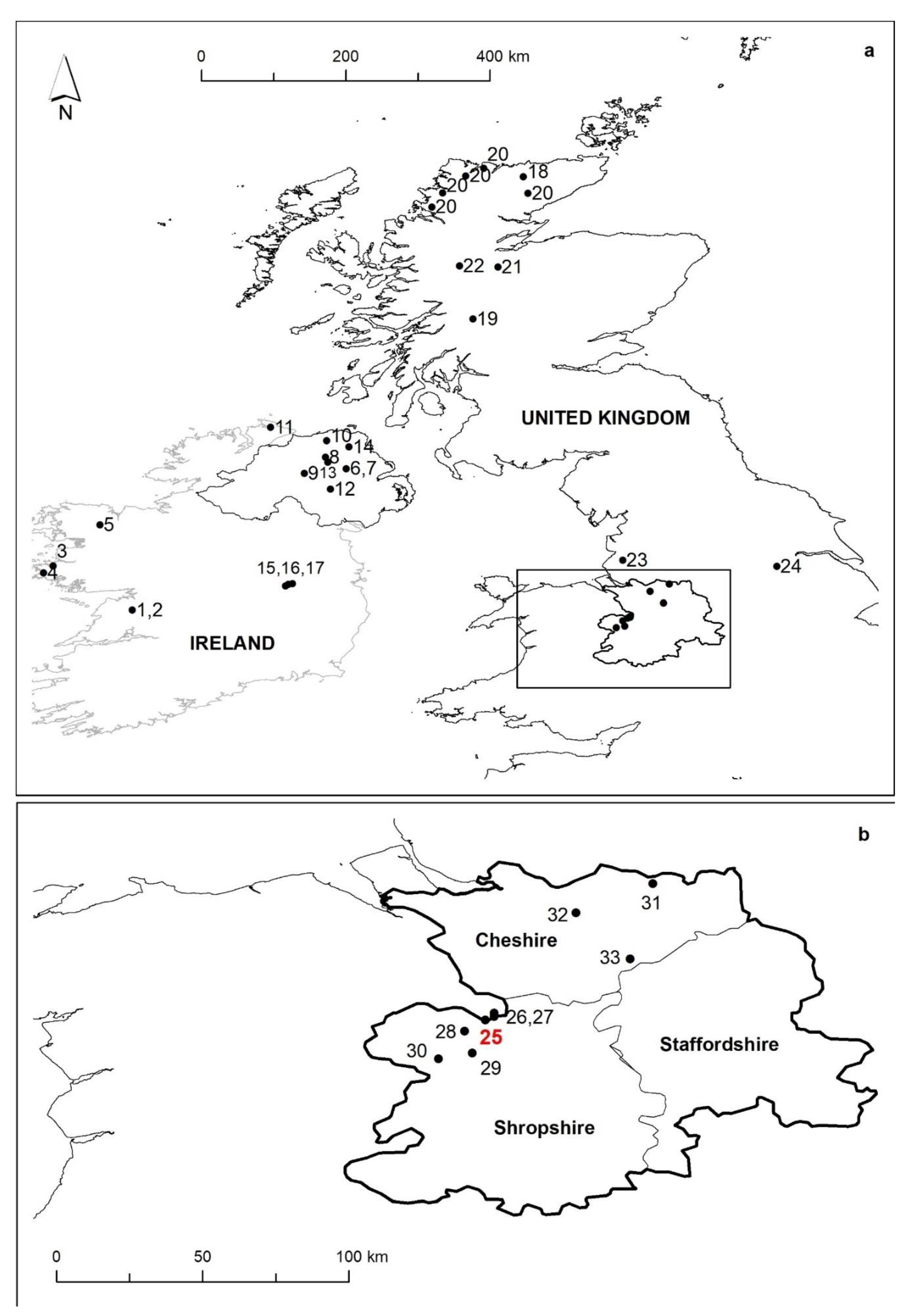
2. Study Area and Site
3. Materials and Methods
3.1. Dendrochronology
3.2. Radiocarbon Dating
4. Results
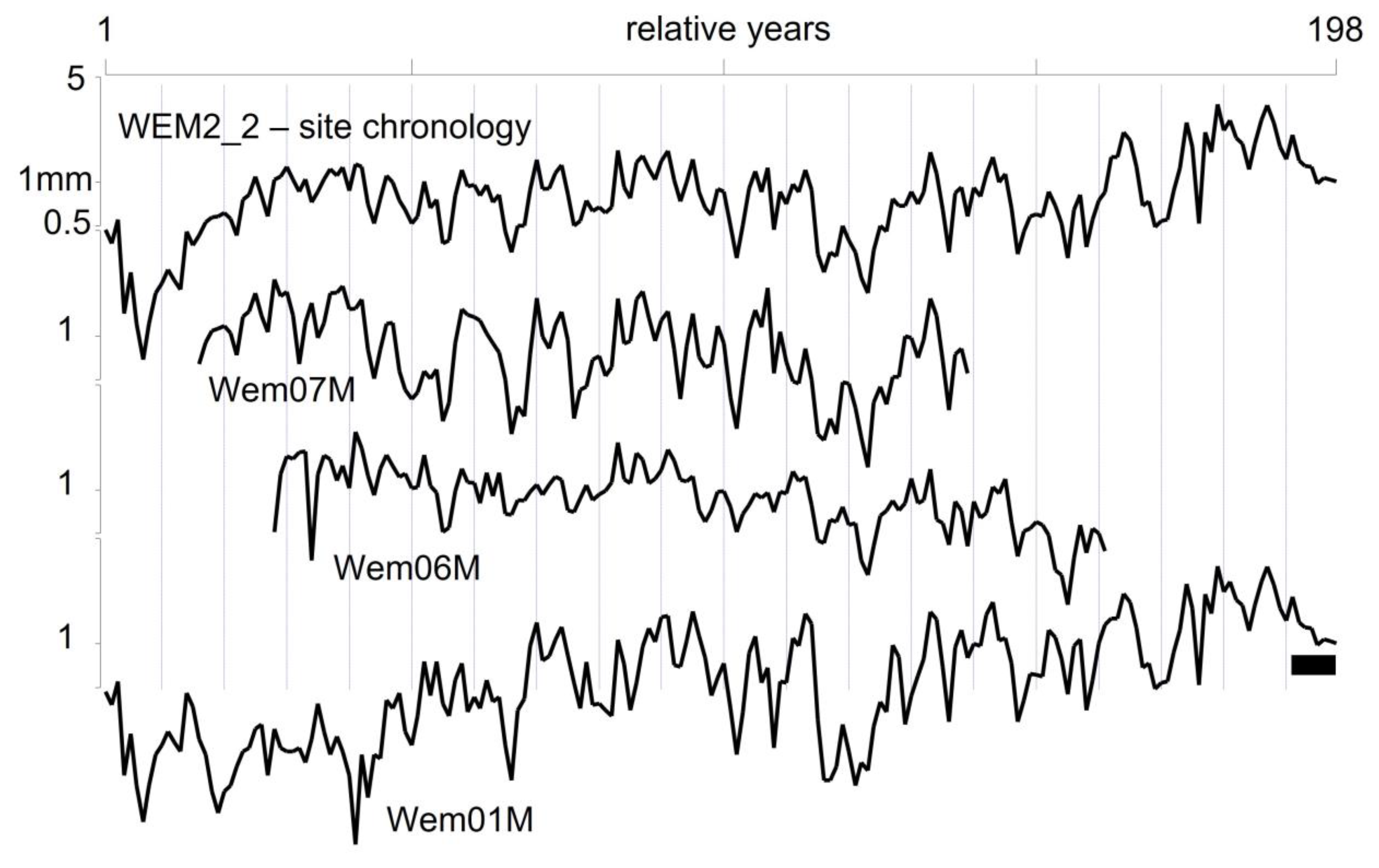
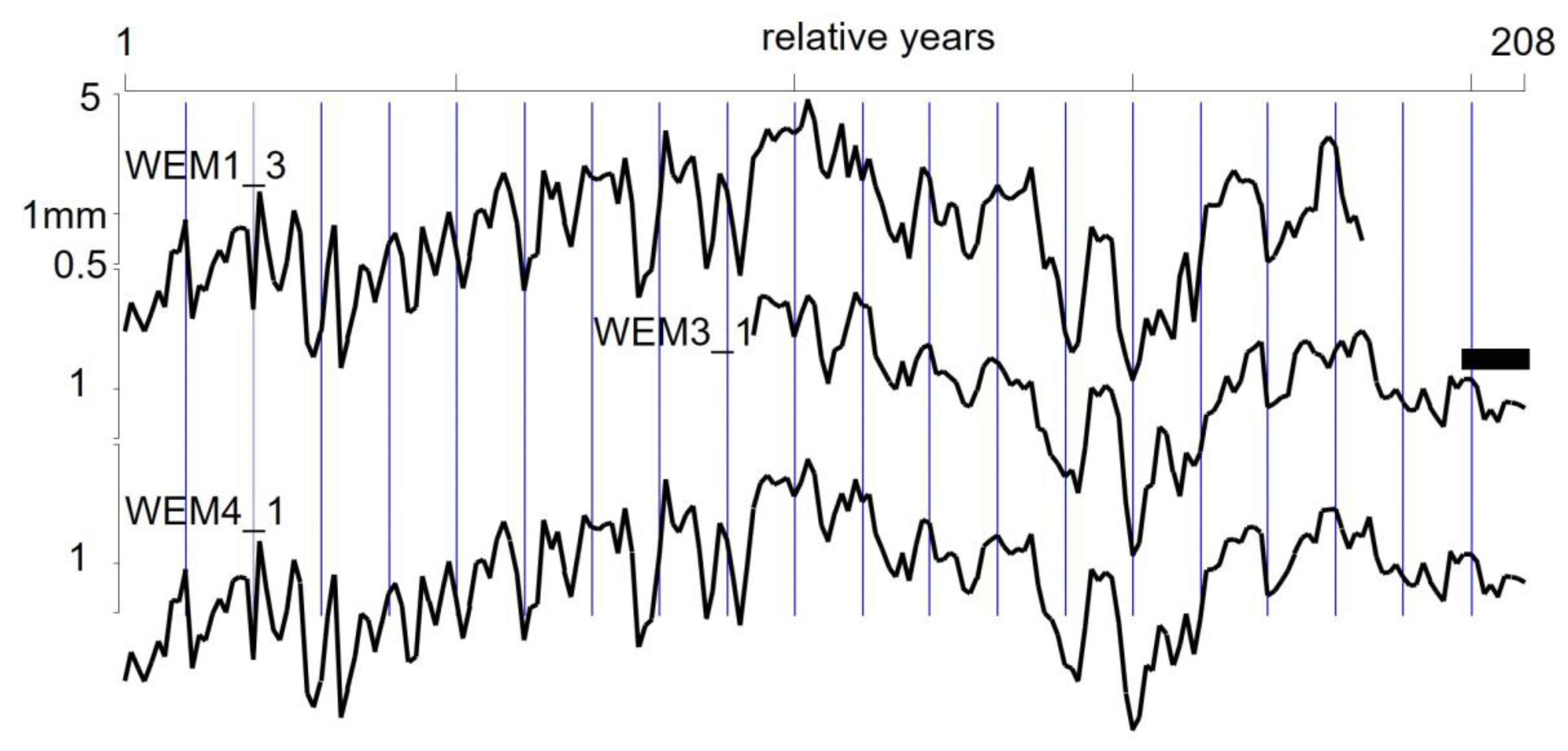
4.1. Dendrochronology
4.2. Radiocarbon Dating
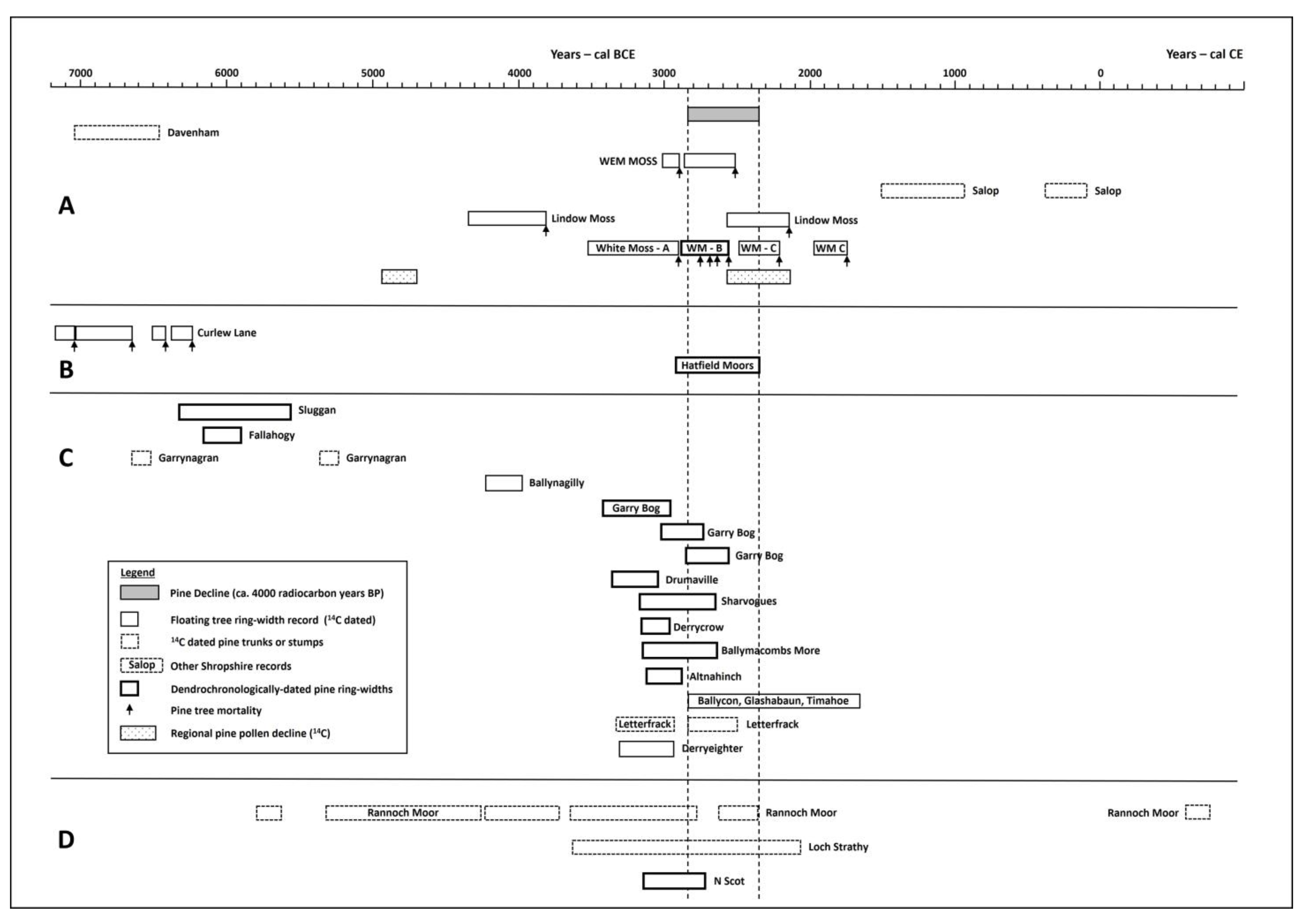
5. Discussion


Wider Context
6. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mukassabi, T.A.; Thomas, P.A.; Coleshaw, T.; Polwart, A. Is Scots Pine a successful invader in a contemporary bog? Int. Proc. Chem. Biol. Env. Eng. 2015, 88, 72–80. [Google Scholar]
- Edvardsson, J.; Stoffel, M.; Corona, C.; Bragazza, L.; Leuschner, H.H.; Charman, D.J.; Helama, S. Subfossil peatland trees as proxies for Holocene palaeohydrology and palaeoclimate. Earth Sci. Revs. 2016, 163, 118–140. [Google Scholar] [CrossRef]
- Smiljanić, M.; Seo, J.-W.; Läänelaid, A.; van der Maaten-Theunissen, M.; Stajić, B.; Wilmking, M. Peatland pines as a proxy for watertable fluctuations: Disentangling tree growth, hydrology and possible human influence. Sci. Total Environ. 2014, 500–501, 52–63. [Google Scholar] [CrossRef]
- Edvardsson, J.; Rimkus, E.; Corona, C.; Šimanauskiene, R.; Kažys, J.; Stoffel, M. Exploring the impact of regional climate and local hydrology on Pinus sylvestris L. growth variability—A comparison between pine populations growing on peat soils and mineral soils in Lithuania. Plant Soil 2015, 392, 345–356. [Google Scholar] [CrossRef]
- Edvardsson, J.; Šimanauskiene, R.; Taminskas, J.; Baužiene, J.; Stoffel, M. Increased tree establishment in Lithuanian peat bogs detected using a combination of field and remotely sensed approaches. Sci. Total Environ. 2015, 505, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Dang, Q.L.; Lieffers, V.J. Assessment of patterns of response of tree ring growth of black spruce following peatland drainage. Can. J. For. Res. 1989, 19, 924–929. [Google Scholar] [CrossRef]
- Schulthess, J. Der Einfluss von Entwasserung auf Bewaldung eines Hochmoores: Eine Studie zur Rezenten Bewaldungsentwicklung am Etang de la Gruere (JU); Unpublished Diplomarbeit; Geographischer Institut der Universitat Zurich-Irchel: Zurich, Switzerland, 1990. [Google Scholar]
- MacDonald, S.E.; Yin, F. Factors influencing size inequality in peatland black spruce and tamarack: Evidence from a post-drainage release growth. J. Ecol. 1999, 87, 404–412. [Google Scholar] [CrossRef]
- Freléchoux, F.; Buttler, A.; Schweingruber, F.H.; Gobat, J.-M. Stand structure, invasion, and growth dynamics of bog pine (Pinus uncinata var. rotundata) in relation to peat cutting and drainage in the Jura Mountains, Switzerland. Can. J. For. Res. 2000, 30, 1114–1126. [Google Scholar] [CrossRef]
- McNally, A.; Doyle, G.J. A study of subfossil pine layers in a raised bog complex in the Irish Midlands: 1. Palaeowoodland extent and dynamics. Proc. Roy. Irish Acad. 1984, 6–7, 57–70. [Google Scholar]
- Tallis, J.H. Tree remains in southern Pennine peats. Nature 1975, 256, 482–484. [Google Scholar] [CrossRef]
- Tallis, J.H. The uplands: Human influence on the plant cover. In Ecology & Landscape Development: A History of the Mersey Basin; Greenwood, E.R., Ed.; Liverpool University Press: Liverpool, UK, 1999; pp. 109–121. [Google Scholar]
- Chambers, F.M.; Grant, M.E.; Lageard, J.G.A.; Roberts, L.J.; Thomas, P.A. The Palaeoenvironmental Record. In Fenn’s and Whixall Mosses; Berry, A.Q., Daniels, J.F., Allmark, W., Eds.; Clwyd County Council: Mold, UK, 1996; pp. 27–40. [Google Scholar]
- Feehan, J.; O’Donovan, G. The Bogs of Ireland: An Introduction to the Natural, Cultural and Industrial Heritage of Irish Peatlands; The Environmental Institute, University College Dublin: Dublin, Ireland, 1996; pp. 1–530. [Google Scholar]
- Eckstein, J.; Leuschner, H.H.; Giesecke, T.; Shumilovskikh, L.; Bauerochse, A. Dendroecological investigations at Venner Moor (northwest Germany) document climate-driven woodland dynamics and mire development in the period 2450–2050 BC. Holocene 2010, 20, 231–244. [Google Scholar] [CrossRef]
- Lageard, J.G.A.; Robinson, E.A. An investigation of subfossil Scots pine (Pinus sylvestris) from Curlew Lane, Lancashire. North West Geogr. 2022, 22(2), 1–13. [Google Scholar]
- Aldhouse-Green, M. Bog Bodies Uncovered: Solving Europe’s Ancient Mystery; Thames & Hudson: London, UK, 2015; pp. 1–223. [Google Scholar]
- Turner-Walker, G.; Peacock, E.E. Preliminary results of bone diagenesis in Scandinavian bogs. Palaeogeog. Palaeoclim. Palaeoecol. 2008, 266, 151–159. [Google Scholar] [CrossRef]
- Tallis, J.H.; Switsur, V.R. Forest and moorland in the south Pennine uplands in the mid-Flandrian period: I. Macrofossil evidence of the former forest cover. J. Ecol. 1983, 71, 585–600. [Google Scholar] [CrossRef]
- Godwin, H. History of the British Flora: A Factual Basis for Phytogeography, 2nd ed.; Cambridge University Press: Cambridge, UK, 1975; pp. 1–541. [Google Scholar]
- Munaut, A.V. Recherches dendrochronologiques sur Pinus silvestris, 1. Étude de 45 pins sylvestres récents originaires de Belgique. Agricultura 1966, 14, 193–232. [Google Scholar]
- Munaut, A.V. Recherches dendrochronologiques sur Pinus silvestris, 2. Première applications des methodes dendrochronologique a l’étude de pins sylvestres subfossiles (Terneuzen, Pay-Bas). Agricultura 1966, 14, 361–389. [Google Scholar]
- Godwin, H. Terneuzen and buried forests of the East Anglian fenland. New Phytol. 1968, 67, 733–738. [Google Scholar] [CrossRef]
- Munaut, A.V.; Casparie, W.A. Étude dendrochronologique des Pinus sylvestris L. subfossiles provenant de la tourbière D’Emmen (Drenthe, Pays-Bas). Rev. Palaeo. Palynol. 1971, 11, 201–226. [Google Scholar] [CrossRef]
- Lageard, J.G.A.; Chambers, F.M.; Thomas, P.A. Climatic significance of the marginalisation of Scots pine (Pinus sylvestris L.) circa 2500 BC at White Moss, south Cheshire, UK. Holocene 1999, 9, 321–332. [Google Scholar] [CrossRef]
- Lageard, J.G.A.; Thomas, P.A.; Chambers, F.M. Using fire scars and growth release in subfossil Scots pine to reconstruct prehistoric fires. Palaeogeog. Palaeoclim. Palaeoecol. 2000, 164, 87–99. [Google Scholar] [CrossRef]
- Leuschner, H.H.; Sass-Klaassen, U.; Jansma, E.; Baillie, M.G.L.; Spurk, M. Subfossil European bog oaks: Population dynamics and long-term growth depressions as indicators of changes in the Holocene hydro-regime and climate. Holocene 2002, 12, 695–706. [Google Scholar] [CrossRef]
- Lageard, J.G.A.; Ryan, P. Microscopic fungi as subfossil woodland indicators. Holocene 2013, 23, 990–1001. [Google Scholar] [CrossRef]
- Walker, D. Direction and rate of change in some British post glacial hydroseres. In Studies in the Vegetation History of the British Isles; Walker, D., West, R.G., Eds.; Cambridge University Press: Cambridge, UK, 1970; pp. 117–139. [Google Scholar]
- Moore, P.D. The Origin of blanket mire, revisited. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman & Hall: London, UK, 1993; pp. 217–224. [Google Scholar]
- Atkinson, D.; Smart, R.A.; Fairhurst, J.; Oldfield, P.; Lageard, J.G.A. A history of woodland in the Mersey Basin. In History of the Mersey Basin: Ecology and Landscape Development; Greenwood, E., Ed.; University of Liverpool Press: Liverpool, UK, 1999; pp. 91–107. [Google Scholar]
- Sass-Klaassen, U.; Hanraets, E. Woodlands of the past—The excavation of wetland woods at Zwolle-Stadshagen (the Netherlands): Growth pattern and population dynamics of oak and ash. Neth. J. Geosci. 2006, 85, 61–71. [Google Scholar] [CrossRef]
- Copini, P.; den Ouden, J.; Robert, E.M.R.; Tardif, J.C.; Loesberg, W.A.; Goudzwaard, L.; Sass-Klaassen, U. Flood-ring formation and root development in response to experimental flooding of young Quercus robur trees. Front. Plant Sci. 2016, 7, 775. [Google Scholar] [CrossRef] [PubMed]
- Eckstein, J.; Leuschner, H.H.; Giesecke, T.; Bauerochse, A.; Sass-Klaassen, U. Subfossil bog-pine horizons document climate and ecosystem changes during the Mid-Holocene. Dendrochronologia 2009, 27, 129–146. [Google Scholar] [CrossRef]
- Margielewski, W.; Krąpiec, M.; Kupryjanowicz, M.; Fiłoc, M.; Buczek, K.; Stachowicz-Rybka, R.; Obidowicz, A.; Pociecha, A.; Szychowska-Krąpiec, E.; Sala, D.; et al. Bog pine dendrochronology related to peat stratigraphy: Palaeoenvironmental changes reflected in peatland deposits since the Late Glacial (case study of the Imszar raised bog, Northeastern Poland). Quat. Int. 2022, 613, 61–80. [Google Scholar] [CrossRef]
- Bennett, K.D. The post-glacial history of Pinus sylvestris in the British Isles. Quat. Sci. Revs. 1984, 3, 133–155. [Google Scholar] [CrossRef]
- Lamb, H.H. Trees and climatic history in Scotland. Quart. J. Roy. Met. Soc. 1964, 90, 382–394. [Google Scholar] [CrossRef]
- Pennington, W.; Haworth, E.Y.; Bonny, A.P.; Lishman, J.P. Lake sediments in northern Scotland. Phil. Trans. Roy. Soc. Lond. Ser. B 1972, 264, 191–294. [Google Scholar]
- Birks, H.H. Studies in the vegetational history of Scotland. IV. Pine stumps in Scottish blanket peats. Phil. Trans. Roy. Soc. Lond. Ser. B 1975, 270, 181–226. [Google Scholar]
- Birks, H.J.B. The Flandrian forest history of Scotland: A preliminary synthesis. In British Quaternary Studies: Recent Advances; Shotton, F.W., Ed.; Clarendon Press: Oxford, UK, 1977; pp. 119–135. [Google Scholar]
- Blackford, J.J.; Edwards, K.J.; Dugmore, A.J.; Cook, G.T.; Buckland, P.C. Icelandic volcanic ash and the mid-Holocene Scots pine (Pinus sylvestris) pollen decline in northern Scotland. Holocene 1992, 2, 260–265. [Google Scholar] [CrossRef]
- Hall, V.A.; Pilcher, J.R.; McCormac, F.G. Icelandic volcanic ash and the mid-Holocene Scots pine (Pinus sylvestris) decline in the north of Ireland: No correlation. Holocene 1994, 4, 79–83. [Google Scholar] [CrossRef]
- Hall, V.A.; McVicker, S.J.; Pilcher, J.R. Tephra-Linked Landscape History around 2310 BC of Some Sites in Counties Antrim and Down. Biol. & Env.: Proc. Roy. Irish Acad. 1994, 94B, 245–253. [Google Scholar]
- Edwards, K.J.; Dugmore, A.J.; Buckland, P.C.; Blackford, J.J.; Cook, G.T. Hekla-4 ash, the pine decline in Northern Ireland and the effective use of tephra isochrones: A comment on Hall, Pilcher and McCormac. Holocene 1996, 6, 495–496. [Google Scholar] [CrossRef]
- Hall, V.A.; Pilcher, J.R.; McCormac, F.G. Hekla-4 ash, the pine decline in Northern Ireland and the effective use of tephra isochrones: A reply to Edwards, Dugmore, Buckland, Blackford and Cook. Holocene 1996, 6, 496–497. [Google Scholar] [CrossRef]
- Edwards, K.J.; Dugmore, A.J.; Blackford, J.J. Vegetational response to tephra deposition and land-use change in Iceland: A modern analogue and multiple working hypothesis approach to tephropalynology. Polar Rec. 2004, 40, 113–120. [Google Scholar] [CrossRef]
- Payne, R.J.; Edwards, K.J.; Blackford, J.J. Volcanic impacts on the Holocene vegetation history of Britain and Ireland? A review and meta-analysis of the pollen evidence. Veget. Hist. Archaeobot. 2013, 22, 153–164. [Google Scholar] [CrossRef]
- Leah, M.D.; Wells, C.E.; Appleby, C.; Huckerby, E. The Wetlands of Cheshire. North-West Wetlands Survey 4; Lancaster University Archaeology Unit: Lancaster, UK, 1997; pp. 1–246. [Google Scholar]
- Leah, M.D.; Wells, C.E.; Stamper, P.; Huckerby, E.; Welch, C. North-West Wetlands Survey 5: The Wetlands of Shropshire and Staffordshire; Lancaster University Archaeological Unit: Kendall, UK, 1998; pp. 1–262. [Google Scholar]
- Natural England. NCA Profile: 61 Shropshire, Cheshire and Staffordshire Plain (NE556); Natural England: Peterborough, UK, 2014; ISBN 978-1-78367-116-8. Available online: http://publications.naturalengland.org.uk/publication/6076647514046464?category=587130 (accessed on 11 October 2022).
- Sinker, C.A. The north Shropshire meres and mosses: A background for ecologists. Field Stud. 1962, 1, 101–138. [Google Scholar]
- Turner, J. The Tilia decline, an anthropogenic interpretation. New Phytol. 1962, 61, 328–341. [Google Scholar] [CrossRef]
- Turner, J. The Anthropogenic Factor in Vegetational History. I. Tregaron and Whixall Mosses. New Phytol. 1964, 63, 73–90. [Google Scholar] [CrossRef]
- Tallis, J.H. The terrestrialisation of lake basins in North Cheshire, with special reference to the development of Schwingmoor structures. J. Ecol. 1973, 61, 537–567. [Google Scholar] [CrossRef]
- Green, B.H.; Pearson, M.C. Ecology of Wybunbury Moss II Post-Glacial history and formation of the Cheshire mere and mire landscape. J. Ecol. 1977, 65, 793–814. [Google Scholar] [CrossRef]
- Hayek, T. The Meres & Mosses of the Marches Landscape Partnership Scheme Final Report: Beyond the Scheme; The Meres & Mosses Landscape Partnership Scheme, Shropshire Wildlife Trust: Shrewsbury, UK, 2018; pp. 1–30. Available online: https://themeresandmosses.co.uk/wp-content/uploads/2019/08/MM_Report_FINALemailversion.pdf (accessed on 11 October 2022).
- Kneen, S.; Lageard, J.G.A. COMM2: Vegetation History from Cole Mere and Clarepool Moss, Shropshire; Report prepared for Shropshire Wildlife Trust & The Meres and Mosses Landscape Partnership Scheme, 2015; pp. 1–36. [Google Scholar] [CrossRef]
- McGeever, A.H.; Mitchell, F.J. Redefining the natural range of Scots Pine (Pinus sylvestris L.): A newly discovered microrefugium in western Ireland. J. Biogeog. 2016, 43, 2199–2208. [Google Scholar] [CrossRef]
- Roche, J.R.; Mitchell, F.J.G.; Waldren, S.; Stefanini, B.S. Palaeoecological evidence for survival of Scots Pine through the Late Holocene in Western Ireland: Implications for ecological management. Forests 2018, 9, 350. [Google Scholar] [CrossRef]
- O’Connell, M.; Jennings, E.; Molloy, K. Holocene vegetation dynamics, landscape change and human impact in western Ireland as revealed by multidisciplinary, palaeoecological investigations of peat deposits and bog-pine in lowland Connemara. Geographies 2021, 1, 251–291. [Google Scholar] [CrossRef]
- Pilcher, J.R.; Baillie, M.G.L.; Brown, D.M.; McCormac, F.G.; Macsweeney, P.B.; McLawrence, A.S. Dendrochronology of sub fossil pine in the north of Ireland. J. Ecol. 1995, 83, 665–672. [Google Scholar] [CrossRef]
- Torbenson, M.C.A.; Plunkett, G.; Brown, D.M.; Pilcher, J.R.; Leuschner, H.H. Asynchrony in key Holocene chronologies: Evidence from Irish bog pines. Geology 2015, 43, 799–802. [Google Scholar] [CrossRef]
- Plunkett, G.; Brown, D.M.; Swindles, G.T. Siccitas magna ultra modum: Examining the occurrence and societal impact of droughts in Prehistoric Ireland. Proc. Roy. Ir. Acad. Sect. C 2020, 120C, 83–104. [Google Scholar] [CrossRef]
- Smith, A.G.; Pilcher, J.R. Radiocarbon dates and vegetational history of the British Isles. New Phytol. 1973, 72, 903–914. [Google Scholar] [CrossRef]
- Pilcher, J.R.; Smith, A.G. A Neolithic and Bronze Age settlement in County Tyrone, Northern Ireland. Phil. Trans. Roy. Soc. London 1979, B1013, 346–369. [Google Scholar]
- Gear, A.J.; Huntley, B. Rapid changes in the range limits of Scots pine 4000 years ago. Science 1991, 251, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Bridge, M.C.; Haggart, B.A.; Lowe, J.J. The history and palaeoclimatic significance of subfossil remains of Pinus sylvestris in blanket peats from Scotland. J. Ecol. 1990, 78, 77–99. [Google Scholar] [CrossRef]
- Moir, A.K.; Leroy, S.A.G.; Brown, D.; Collins, P.E.F. Dendrochronological evidence for a lower water-table on peatland around 3200–3000 BC from subfossil pine in northern Scotland. Holocene 2010, 20, 931–942. [Google Scholar] [CrossRef]
- Froyd, C.A. Fossil stomata reveal early pine presence in Scotland: Implications for postglacial colonization analyses. Ecology 2005, 86, 579–586. [Google Scholar] [CrossRef]
- Boswijk, G.; Whitehouse, N.J. Pinus and Prostomis: A dendrochronological and palaeoentomological study of a mid-Holocene woodland in eastern England. Holocene 2002, 12, 585–596. [Google Scholar] [CrossRef]
- Hardy, E.H. Studies of the Post-Glacial History of the British Vegetation, V. The Shropshire and Flint Maelor Mosses. New Phytol. 1939, 38, 364–396. [Google Scholar] [CrossRef]
- Turner, J. A contribution to the history of forest clearance. Proc. Roy. Soc. Lond. B 1965, 161, 343–354. [Google Scholar]
- Haslam, C.J. Late Holocene Peat Stratigraphy and Climatic Change: A Macrofossil Investigation from the Raised Mires of North Western Europe. Unpublished. Ph.D. Thesis, University of Southampton, Southampton, UK, 1987. [Google Scholar]
- Lageard, J.G.A.; Chambers, F.M.; Grant, M. Study of Vegetation History at Fenn’s and Whixall Mosses Based on a Study of Pine Remains and Pollen in Peat Strata; Interim Report for English Nature and The Countryside Council for Wales, 1994; pp. 1–20. [Google Scholar]
- Grant, M.E. The Dating and Significance of Pinus sylvestris L. Macrofossil Remains from Whixall Moss, Shropshire: Palaeoecological and Modern Comparative Analyses. Unpublished. Ph.D. Thesis, Keele University, Keele, UK, 1995. [Google Scholar]
- Lageard, J.G.A.; Chambers, F.M. The Palaeoecological significance of a new, subfossil-oak chronology (Quercus sp.) from Morris’ Bridge, Shropshire, UK. Dendrochronologia 1994, 11, 25–33. [Google Scholar]
- Beales, P.W. The Late Devensian and Flandrian of Crose Mere, Shropshire. New Phytol. 1980, 85, 133–161. [Google Scholar] [CrossRef]
- Barber, K.E.; Twigger, S.N. Late Quaternary palaeoecology of the Severn Basin. In Palaeohydrology in Practice; Greggory, K.J., Lewin, J., Thornes, J.B., Eds.; Wiley & Sons: Chichester, UK, 1987; pp. 219–252. [Google Scholar]
- Sassoon, D.; Fletcher, W.J.; Hotchkiss, A.; Owen, F.; Feng, L. Scots pine (Pinus sylvestris) dynamics in the Welsh Marches during the mid- to late-Holocene. Holocene 2021, 31, 1033–1046. [Google Scholar] [CrossRef]
- Lageard, J.G.A. Dendrochronological analysis and dating of subfossil Pinus sylvestris L. at Lindow Moss, Cheshire. Bull. Brit. Ecol. Soc. 1998, 29, 31–32. [Google Scholar]
- Howard-Davies, C.; Buxton, K. I: Excavations at Church Moss, Davenham 1995–6: A post-glacial environmental sequence in Mid-Cheshire. J. Chester Arch. Soc. 1999, 75, 1–17. [Google Scholar]
- Natural England. Natural England Shropshire’s National Nature Reserves (Corporate Report); Natural England: Peterborugh, UK, 2008. Available online: https://www.gov.uk/government/publications/shropshires-national-nature-reserves/shropshires-national-nature-reserves#wem-moss (accessed on 11 October 2022).
- Tyers, I. Dendro for Windows Program Guide, 2nd ed.; Report 500; Archaeological Research and Consultancy at the University of Sheffield (ARCUS): Sheffield, UK, 1999. [Google Scholar]
- Turner, R.C.; Penney, S. Three bog bodies from Whixall Moss, Shropshire. Shrop. Hist. Arch. 1996, 71, 1–9. [Google Scholar]
- Reimer, P.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Bronk Ramsey, C. OxCal Version 4.4. Radiocarbon Calibration Program Utilising IntCal20. 2022. Available online: https://c14.arch.ox.ac.uk/oxcal.html (accessed on 12 October 2022).
- Lageard, J.G.A. Vegetational History and Palaeoforest Reconstruction at White Moss, South Cheshire, UK. Unpublished. Ph.D. Thesis, Keele University, Keele, UK, 1992. [Google Scholar]
- Twigger, S.N. Late Holocene Palaeoecology and Environmental Archaeology of Six Lowland Lakes and Bogs in North Shropshire. Unpublished. Ph.D. Thesis, University of Southampton, Southampton, UK, 1988. [Google Scholar]
- Erdtman, G. Handbook of Palynology, Morphology, Taxonomy, Ecology: An Introduction to the Study of Pollen Grains and Spores; Munsgaard: Copenhagen, Denmark, 1969; pp. 1–486. [Google Scholar]
- Moore, P.D.; Webb, J.A. An Illustrated Guide to Pollen Analysis; Hodder and Stoughton: London, UK, 1978; pp. 1–133. [Google Scholar]
- Bennett, K.D. Post-glacial dynamics of pine (Pinus sylvestris) and pinewoods in Scotland. In Our Pinewood Heritage; Aldhous, J.R., Ed.; Forestry Commission, The Royal Society for the Protection of Birds, Scottish Natural Heritage: Sandy, UK, 1995; pp. 23–39. [Google Scholar]
- Fossitt, J.A. Late-glacial and Holocene vegetation history of western Donegal, Ireland. Biol. Env. Proc. Roy. Ir. Acad. 1994, 94B, 1–31. [Google Scholar]
- Smout, T.C. (Ed.) People and the Woods in Scotland: A History; University of Edinburgh Press: Edinburgh, UK, 2003; pp. 1–244. [Google Scholar]
- Tipping, R.; Ashmore, P.; Davies, A.L.; Haggart, A.; Moir, A.; Newton, A.; Sands, R.; Skinner, T.; Tisdall, E. Prehistoric Pinus woodland dynamics in an upland landscape in northern Scotland: The roles of climate change and human impact. Veg. Hist. Archaeobot. 2008, 17, 251–267. [Google Scholar] [CrossRef]
- Manning, A.D.; Kesteven, J.; Stein, J.; Lunn, A.; Xu, T.; Rayner, B. Could native Scots pines (Pinus sylvestris) still persist in northern England and southern Scotland? Plant Ecol. Div. 2010, 3, 187–201. [Google Scholar] [CrossRef]
- O’Connell, M.; Molloy, K. Aran Islands, western Ireland: Farming history and environmental change reconstructed from field surveys, historical sources, and pollen analyses. J. Nor. Atlantic 2019, 38, 1–27. [Google Scholar] [CrossRef]
- O’Connell, M. Bog-deal in Co. Clare, with particular reference to bog-pine and its significance. J. Shannon Arch. Hist. Soc. 2022, 46, 97–105. [Google Scholar]
- Day, S.P. Woodland origin and ‘ancient woodland indicators’: A case-study from Sidlings Copse, Oxfordshire, UK. The Holocene 1993, 3, 45–53. [Google Scholar] [CrossRef]
- Salmela, M.J.; Cavers, S.; Cottrell, J.E.; Iason, G.R.; Ennos, R.A. Seasonal patterns of photochemical capacity and spring phenology reveal genetic differentiation among native Scots pine (Pinus sylvestris L.) populations in Scotland. For. Ecol. Manage. 2011, 262, 1020–1029. [Google Scholar] [CrossRef]
- Donnelly, K.; Cavers, S.; Cottrell, J.E.; Ennos, R.A. Genetic variation for needle traits in Scots pine (Pinus sylvestris L.). Tree Genet. Genomes 2016, 12, 40. [Google Scholar] [CrossRef]
- Donnelly, K.; Cavers, S.; Cottrell, J.E.; Ennos, R.A. Cryptic genetic variation and adaptation to waterlogging in Caledonian Scots pine, Pinus sylvestris L. Ecol. Evol. 2018, 8, 8665–8675. [Google Scholar] [CrossRef] [PubMed]
- Kastally, C.; Niskanen, A.K.; Perry, A.; Kujala, S.T.; Avia, K.; Cervantes, S.; Haapanen, M.; Kesalahti, R.; Kumpala, T.A.; Mattila, T.M.; et al. Taming the massive genome of Scots pine with PiSy50k, a new genotyping array for conifer research. Plant J. 2022, 109, 1337–1350. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wem01 | Wem02 | Wem03 | Wem04 | Wem05 | Wem06 | Wem07 | Wem08 | Wem09 | Wem10 | Wem11 | Wem12 | Wem13 | Wem14 | |
| Wem01 | 5.38 | 8.14 | ||||||||||||
| Wem02 | 5.74 | |||||||||||||
| Wem03 | 6.16 | |||||||||||||
| Wem04 | 5.29 | 9.2 | ||||||||||||
| Wem05 | 6.79 | |||||||||||||
| Wem06 | 5.72 | |||||||||||||
| Wem07 | 6.19 | |||||||||||||
| Wem08 | 6.57 | |||||||||||||
| Wem09 | ||||||||||||||
| Wem10 | ||||||||||||||
| Wem11 | ||||||||||||||
| Wem12 | ||||||||||||||
| Wem13 | ||||||||||||||
| Wem14 |
| Site Chronology | Component Chronologies | Number of Samples | Length (Years) | Average Ring-Width (mm) | Sensitivity |
|---|---|---|---|---|---|
| WEM1_3 | - | 4 | 184 | 102.99 | 0.38 |
| WEM2_2 | - | 3 | 198 | 90.48 | 0.31 |
| WEM3_1 | - | 2 | 115 | 123.77 | 0.23 |
| WEM4_1 | WEM1_3 & WEM3_1 | 6 | 208 | 103.24 | 0.33 |
| Chronology | Tree | Chronology Years | Radiocarbon Age (14C Years BP) | Calibrated Age Range–Years (2 Sigma) |
|---|---|---|---|---|
| WEM2_2 | Wem01 | 191–198 | 4330 ± 30 (Beta–424347) | 3015–2895 cal. BCE |
| WEM4_1 | Wem10 | 199–208 | 4100 ± 30 (Beta–424348) | 2860–2505 cal. BCE |
| Publication | Site | Bog-Oak/Bog-Pine/Pine Pollen | 14C Age (Years BP) | Calibrated Age Range (2 Sigma) (Years cal. BCE/CE) | Calendar Date (Years BCE) | Artefact Dating (Years BCE) | Undated Tree-Ring Series |
|---|---|---|---|---|---|---|---|
| [72] | Whixall Moss | Pine | - | - | - | 1500–1000 | - |
| [53] | Whixall Moss | Pine | 2307 ± 110 | 761–106 BCE | - | - | - |
| [73] | Whixall Moss | Pine pollen | ca. 2000 | - | - | - | - |
| [78] | Crose Mere | Pine pollen | 2310 ± 85 | 753–164 BCE | - | - | - |
| [79] | Fenemere | Pine pollen | 1890 ± 50 | 232–248 CE | - | - | - |
| [74] | Whixall Moss | Pine | 2180 ± 50 | 397–3 BCE | - | - | ✓ |
| [75] | Whixall Moss | Pine | - | - | - | - | ✓ |
| [77] | Morris’ Bridge | Oak | - | - | 4596–4304 | - | - |
| [76] | Whixall Moss | Pine | (6 x 14C dates) Oldest: 3140 ± 45 Youngest: 2900 + 40 | 1503–937 BCE | - | - | - |
| Current | Wem Moss | Pine | 4330 ± 30 4100 ± 30 | 3015–2895 BCE 2860–2505 BCE | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lageard, J.G.A. ‘Pine Decline or pine declines?’ Analysis and Interpretation of Bog-Pines from Wem Moss, Shropshire, UK. Quaternary 2023, 6, 12. https://doi.org/10.3390/quat6010012
Lageard JGA. ‘Pine Decline or pine declines?’ Analysis and Interpretation of Bog-Pines from Wem Moss, Shropshire, UK. Quaternary. 2023; 6(1):12. https://doi.org/10.3390/quat6010012
Chicago/Turabian StyleLageard, Jonathan G. A. 2023. "‘Pine Decline or pine declines?’ Analysis and Interpretation of Bog-Pines from Wem Moss, Shropshire, UK" Quaternary 6, no. 1: 12. https://doi.org/10.3390/quat6010012