
Genetic and Epigenetic Study of Monozygotic Twins Affected by Parkinson’s Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill-Burns, E.M.; Ross, O.; Wissemann, W.; Soto-Ortolaza, A.; Zareparsi, S.; Siuda, J.; Lynch, T.; Wszolek, Z.; Silburn, P.; Mellick, G.; et al. Identification of Genetic Modifiers of Age-at-Onset for Familial Parkinson’s Disease. Hum. Mol. Genet. 2016, 25, 3849–3862. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Gonzalez-Latapi, P.; Marras, C.; Visanji, N.; Yang, W.; Sato, C.; Lang, A.; Rogaeva, E.; Zhang, M. Epigenetic Clock Acceleration Is Linked to Age at Onset of Parkinson’s Disease. Mov. Disord. 2022, 37, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xi, Z.; Ghani, M.; Jia, P.; Pal, M.; Werynska, K.; Moreno, D.; Sato, C.; Liang, Y.; Robertson, J.; et al. Genetic and Epigenetic Study of Als-Discordant Identical Twins with Double Mutations in Sod1 and Arhgef28. J. Neurol. Neurosurg. Psychiatry 2016, 87, 1268–1270. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Yunusova, Y.; van Blitterswijk, M.; Dib, S.; Ghani, M.; Moreno, D.; Sato, C.; Liang, Y.; Singleton, A.; Robertson, J.; et al. Identical Twins with the C9orf72 Repeat Expansion Are Discordant for Als. Neurology 2014, 83, 1476–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.; et al. Mds Clinical Diagnostic Criteria for Parkinson’s Disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Wu, J.; Zhao, J.; Huang, S.; Wang, Y.; Ge, J.; Wu, P.; Zuo, C.; Ding, Z.; Wang, J. Onset-Related Subtypes of Parkinson’s Disease Differ in the Patterns of Striatal Dopaminergic Dysfunction: A Positron Emission Tomography Study. Park. Relat. Disord. 2015, 21, 1448–1453. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and Guidelines for the Interpretation of Sequence Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, M.; Wang, W.; Zhao, J.; Wu, J.; Liu, Y.; Zhu, H.; Qu, J.; Zhou, M. Identification of Driver Genes and Somatic Mutations in Cell-Free DNA of Patients with Pulmonary Lymphangioleiomyomatosis. Int. J. Cancer 2020, 146, 103–114. [Google Scholar] [CrossRef]
- Nalls, M.A.; Blauwendraat, C.; Vallerga, C.; Heilbron, K.; Bandres-Ciga, S.; Chang, D.; Tan, M.; Kia, D.; Noyce, A.; Xue, A.; et al. Identification of Novel Risk Loci, Causal Insights, and Heritable Risk for Parkinson’s Disease: A Meta-Analysis of Genome-Wide Association Studies. Lancet Neurol. 2019, 18, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.; Ono, S.; Imamura, A.; Okazaki, Y.; Kinoshita, A.; Mishima, H.; Nakane, H.; Ozawa, H.; Yoshiura, K.; Kurotaki, N. Deep sequencing reveals variations in somatic cell mosaic mutations between monozygotic twins with discordant psychiatric disease. Hum. Genome Var. 2017, 4, 17032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishioka, M.; Bundo, M.; Ueda, J.; Yoshikawa, A.; Nishimura, F.; Sasaki, T.; Kakiuchi, C.; Kasai, K.; Kato, T.; Iwamoto, K. Identification of somatic mutations in monozygotic twins discordant for psychiatric disorders. NPJ Schizophr. 2018, 4, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meltz Steinberg, K.; Nicholas, T.; Koboldt, D.; Yu, B.; Mardis, E.; Pamphlett, R. Whole genome analyses reveal no pathogenetic single nucleotide or structural differences between monozygotic twins discordant for amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2015, 16, 385–392. [Google Scholar] [CrossRef]
- Yu, E.; Rudakou, U.; Krohn, L.; Mufti, K.; Ruskey, J.; Asayesh, F.; Estiar, M.; Spiegelman, D.; Surface, M.; Fahn, S.; et al. Analysis of Heterozygous PRKN Variants and Copy-Number Variations in Parkinson’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2021, 36, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.F.; Dong, X.; Xu, Q.; Li, N.; Yan, X.; Xia, K.; Tang, B. Exon dosage analysis of parkin gene in Chinese sporadic Parkinson’s disease. Neurosci. Lett. 2015, 604, 47–51. [Google Scholar] [CrossRef]
- Islam, M.S.; Azim, F.; Saju, H.; Zargaran, A.; Shirzad, M.; Kamal, M.; Fatema, K.; Rehman, S.; Azad, M.; Ebrahimi-Barough, S. Pesticides and Parkinson’s disease: Current and future perspective. J. Chem. Neuroanat. 2021, 115, 101966. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Chen, H.; Weisskopf, M.; O’Reilly, E.; McCullough, M.; Calle, E.; Schwarzschild, M.; Thun, M. Pesticide exposure and risk for Parkinson’s disease. Ann. Neurol. 2006, 60, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Hsiao, I.T.; Huang, S.H.; Lui, C.C.; Yen, T.C.; Chang, W.N.; Huang, C.W.; Hsieh, C.J.; Chang, Y.Y.; Lin, K.J. ¹⁸F-FP-(+)-DTBZ positron emission tomography detection of monoaminergic deficient network in patients with carbon monoxide related parkinsonism. Eur. J. Neurol. 2015, 22, 845–852, e59–e60. [Google Scholar] [CrossRef]
- Cilia, R.; Cereda, E.; Akpalu, A.; Sarfo, F.; Cham, M.; Laryea, R.; Obese, V.; Oppon, K.; Del Sorbo, F.; Bonvegna, S.; et al. Natural history of motor symptoms in Parkinson’s disease and the long-duration response to levodopa. Brain 2020, 143, 2490–2501. [Google Scholar] [CrossRef]
- Lai, C.Y.; Chou, M.; Lin, C.; Kao, C. Increased risk of Parkinson disease in patients with carbon monoxide intoxication: A population-based cohort study. Medicine 2015, 94, e869. [Google Scholar] [CrossRef]
- van der Plaat, D.A.; de Jong, K.; de Vries, M.; van Diemen, C.; Nedeljković, I.; Amin, N.; Kromhout, H.; Vermeulen, R.; Postma, D.; van Duijn, C.; et al. Occupational Exposure to Pesticides Is Associated with Differential DNA Methylation. Occup. Environ. Med. 2018, 75, 427–435. [Google Scholar] [CrossRef]
- Prunicki, M.; Stell, L.; Dinakarpandian, D.; de Planell-Saguer, M.; Lucas, R.; Hammond, S.; Balmes, J.; Zhou, X.; Paglino, T.; Sabatti, C.; et al. Exposure to No(2), Co, and Pm(2.5) Is Linked to Regional DNA Methylation Differences in Asthma. Clin. Epigenet. 2018, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, D.; Postuma, R.; Adler, C.; Bloem, B.; Chan, P.; Dubois, B.; Gasser, T.; Goetz, C.; Halliday, G.; Joseph, L.; et al. MDS research criteria for prodromal Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2015, 30, 1600–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balck, A.; Borsche, M.; Kasten, M.; Lohmann, K.; Seibler, P.; Brüggemann, N.; Klein, C. Discordance in monozygotic Parkinson’s disease twins—Continuum or dichotomy? Ann. Clin. Transl. Neurol. 2019, 6, 1102–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [Green Version]
- Bergsma, T.; Rogaeva, E. DNA Methylation Clocks and Their Predictive Capacity for Aging Phenotypes and Healthspan. Neurosci. Insights 2020, 15, 2633105520942221. [Google Scholar] [CrossRef]
- Zhang, M.; McKeever, P.; Xi, Z.; Moreno, D.; Sato, C.; Bergsma, T.; McGoldrick, P.; Keith, J.; Robertson, J.; Zinman, L.; et al. DNA methylation age acceleration is associated with ALS age of onset and survival. Acta Neuropathol. 2020, 139, 943–946. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Tartaglia, M.; Moreno, D.; Christine, S.; McKeever, P.; Weichert, A.; Keith, J.; Robertson, J.; Zinman, L.; Rogaeva, E. DNA methylation age-acceleration is associated with disease duration and age at onset in C9orf72 patients. Acta Neuropathol. 2017, 134, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Picillo, M.; Lizarraga, K.; Friesen, E.; Chau, H.; Zhang, M.; Sato, C.; Rooke, G.; Munhoz, R.; Rogaeva, E.; Fraser, P.; et al. Parkinsonism due to A53E alpha-synuclein gene mutation: Clinical, genetic, epigenetic, and biochemical features. Mov. Disord. Off. J. Mov. Disord. Soc. 2018, 33, 1950–1955. [Google Scholar] [CrossRef]
- Horvath, S.; Ritz, B. Increased epigenetic age and granulocyte counts in the blood of Parkinson’s disease patients. Aging 2015, 7, 1130–1142. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
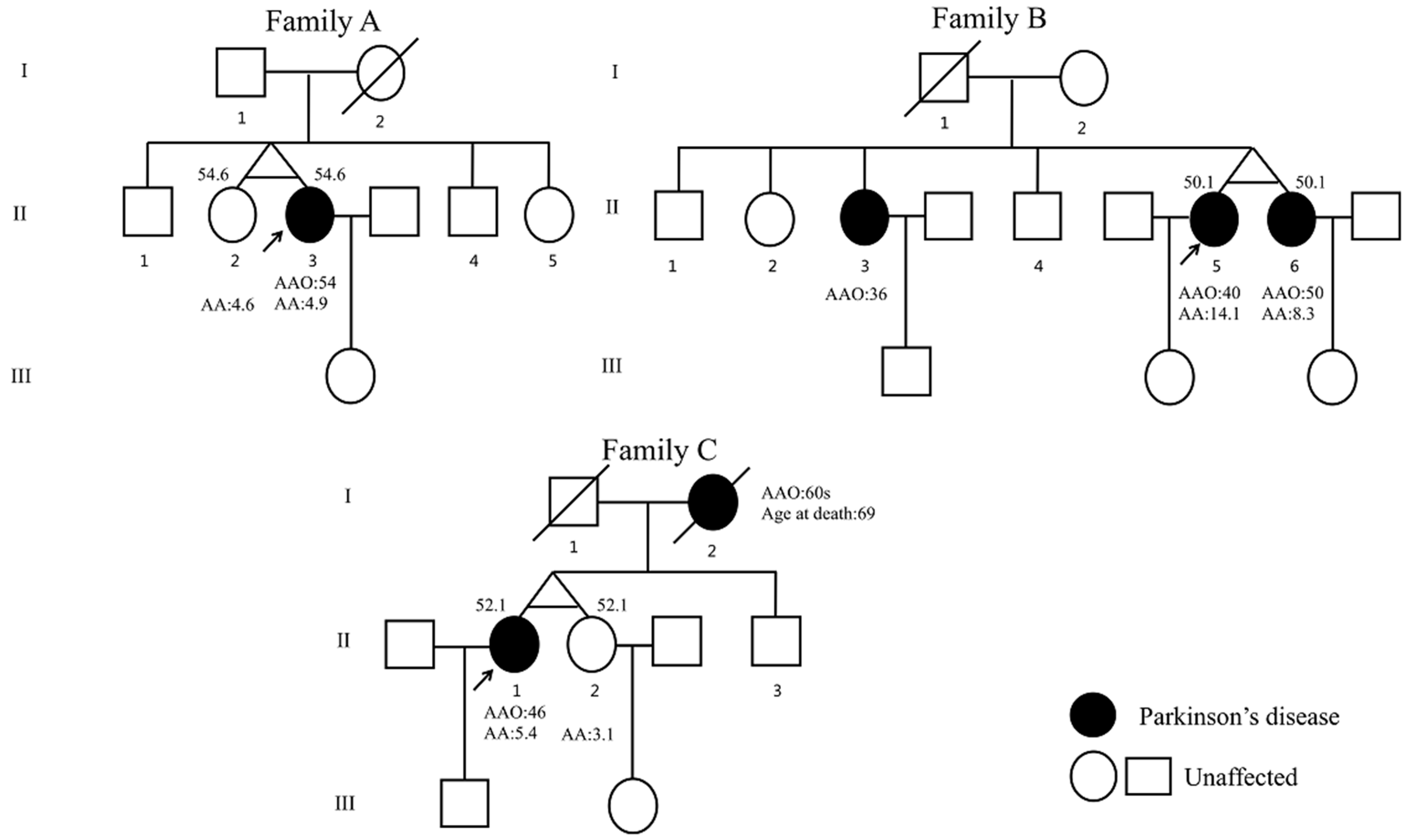
| Clinical Features | Family A | Family B | Family C | ||||
|---|---|---|---|---|---|---|---|
| II-2 | II-3 | II-3 | II-5 | II-6 | II-1 | II-2 | |
| Sex | Female | Female | Female | Female | Female | Female | Female |
| PD AAO, years | - | 54 | 36 | 40 | 50 | 46 | - |
| ASC, years | 55 | 55 | 54 | 51 | 51 | 52 | 52 |
| AOA, years | 55 | 55 | 54 | 51 | 51 | 52 | 52 |
| Types of diet | Normal | Normal | Normal | Normal | Normal | Normal | Normal |
| Occupation | Doctor | Retired cashier of a pesticide and chemical fertilizer company | NA | Worker | Worker in a shoe factory | Civil servant | Civil servant |
| Chemical toxins exposure | No | pesticide and chemical fertilizer | NA | No | No | carbon monoxide poisoning at age 45 | No |
| Head trauma | No | No | NA | No | No | Yes | No |
| Surgery with general anesthesia | No | No | NA | No | No | Ovariectomy | No |
| Cigarette smoking | No | No | NA | No | No | No | No |
| Alcoholic consumption | No | No | NA | light | light | No | No |
| Physical activity in leisure time | No | No | No | No | No | No | No |
| Exposure to pathogens or infectious agents | No | No | No | No | No | No | No |
| Other medical history | No | No | NA | lumbar disc herniation | lumbar disc herniation | No | No |
| Cardinal symptoms of PD | - | Bradykinesia, rigidity | Tremor, Bradykinesia, rigidity | Tremor, Bradykinesia, rigidity | Tremor, Bradykinesia, rigidity | Tremor, Bradykinesia, rigidity | - |
| Family history of PD | No | No | AR | AR | AR | AD | AD |
| Years of education | NA | 10 | 6 | 3 | 3 | 12 | - |
| LEDD, mg | - | NA | 451 | 150 | 250 | 800 | - |
| H&Y stage | - | 2 | 2 | 2 | 2 | 3 | - |
| MMSE score | - | NA | NA | 24 | 28 | NA | - |
| UPDRS-III score | - | 30 (Med-off) | 29 (Med-on) | NA | 33 (Med-off) | 35 (Med-on) | - |
| Methods of genetic analysis | MLPA + WGS | MLPA + WGS | MLPA | MLPA + WGS | MLPA + WGS | MLPA + WGS | WGS |
| Results of genetic analysis | exon 1–3 del (het) in PRKN | exon 1–3 del (het) in PRKN | Exon 2–4 del (het), c.2T > C, p.Met1Thr (het) (NM_004562.3) in PRKN | Exon 2–4 del (het), c.2T > C, p.Met1Thr (het) (NM_004562.3) in PRKN | Exon 2–4 del (het), c.2T > C, p.Met1Thr (het) (NM_004562.3) in PRKN | - | - |
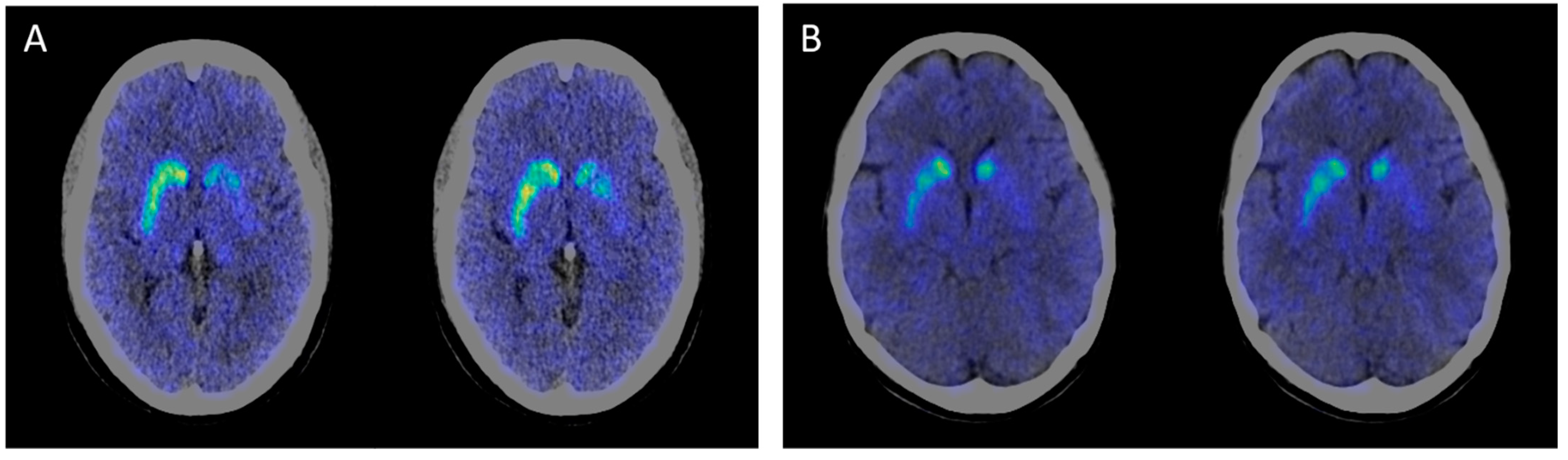
| 11C-CFT PET/CT results | NA | Decreased DAT uptake ratios in bilateral caudate and putamen, especially in the left side | NA | NA | NA | Decreased DAT uptake ratios in bilateral caudate and putamen, especially in the left side | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.-M.; Yang, W.-L.; Rogaeva, E.; Lang, A.E.; Wang, J.; Zhang, M. Genetic and Epigenetic Study of Monozygotic Twins Affected by Parkinson’s Disease. Clin. Transl. Neurosci. 2023, 7, 11. https://doi.org/10.3390/ctn7020011
Sun Y-M, Yang W-L, Rogaeva E, Lang AE, Wang J, Zhang M. Genetic and Epigenetic Study of Monozygotic Twins Affected by Parkinson’s Disease. Clinical and Translational Neuroscience. 2023; 7(2):11. https://doi.org/10.3390/ctn7020011
Chicago/Turabian StyleSun, Yi-Min, Wan-Li Yang, Ekaterina Rogaeva, Anthony E. Lang, Jian Wang, and Ming Zhang. 2023. "Genetic and Epigenetic Study of Monozygotic Twins Affected by Parkinson’s Disease" Clinical and Translational Neuroscience 7, no. 2: 11. https://doi.org/10.3390/ctn7020011