

Evaluation of an Innovative Rosette Flight Plan Design for Wildlife Aerial Surveys with UAS
 , , , , and
, , , , and
Abstract
:1. Introduction
- To design an innovative flight plan and sampling protocol for game counts adapted to small UASs and their constraints, and to test it in under real conditions in the field.
- To evaluate its relevance by comparing the statistical performance of the new design to standard transects based on numerical simulations.
2. Materials and Methods
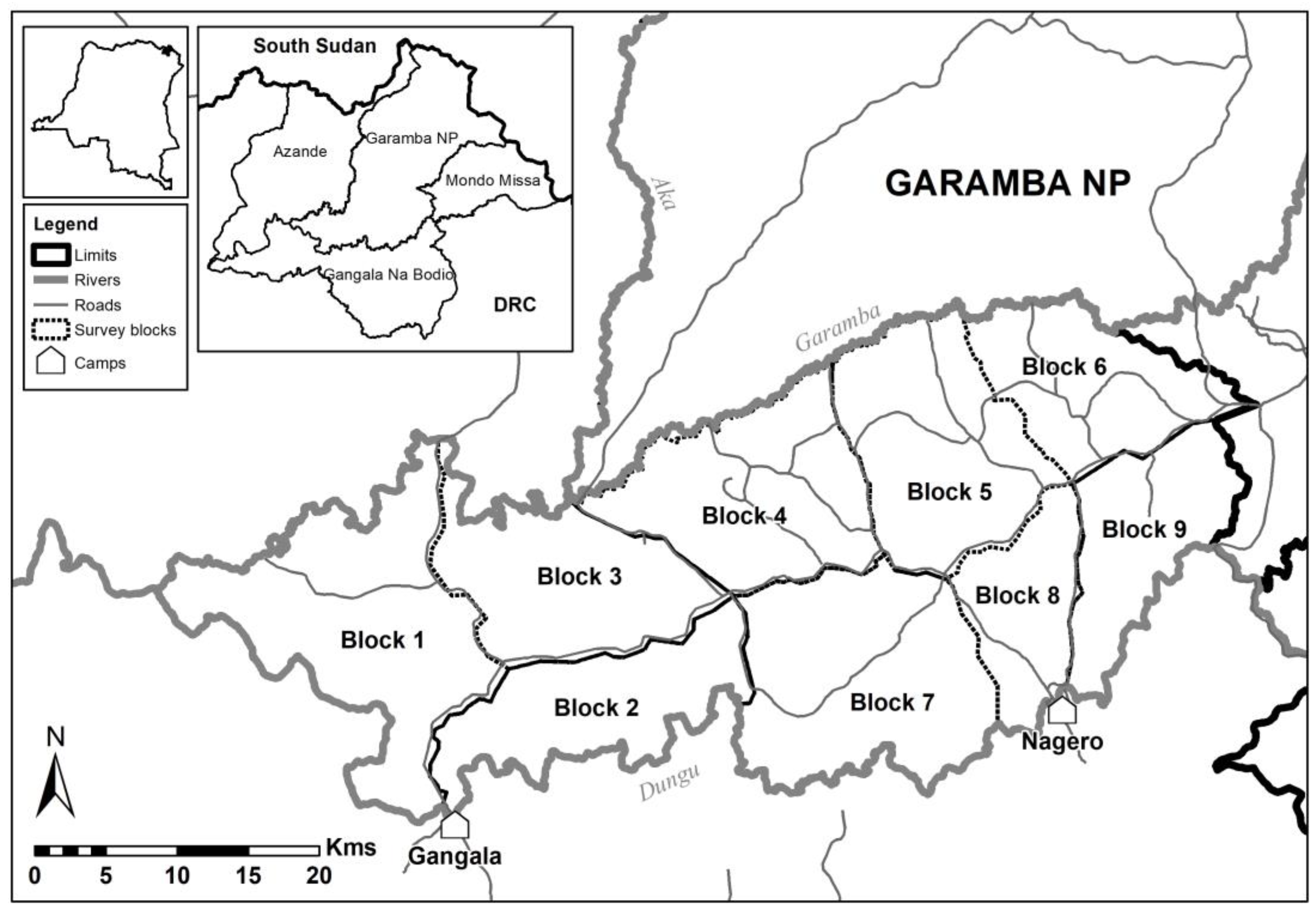
2.1. Study Area
2.2. Unmanned Aerial System
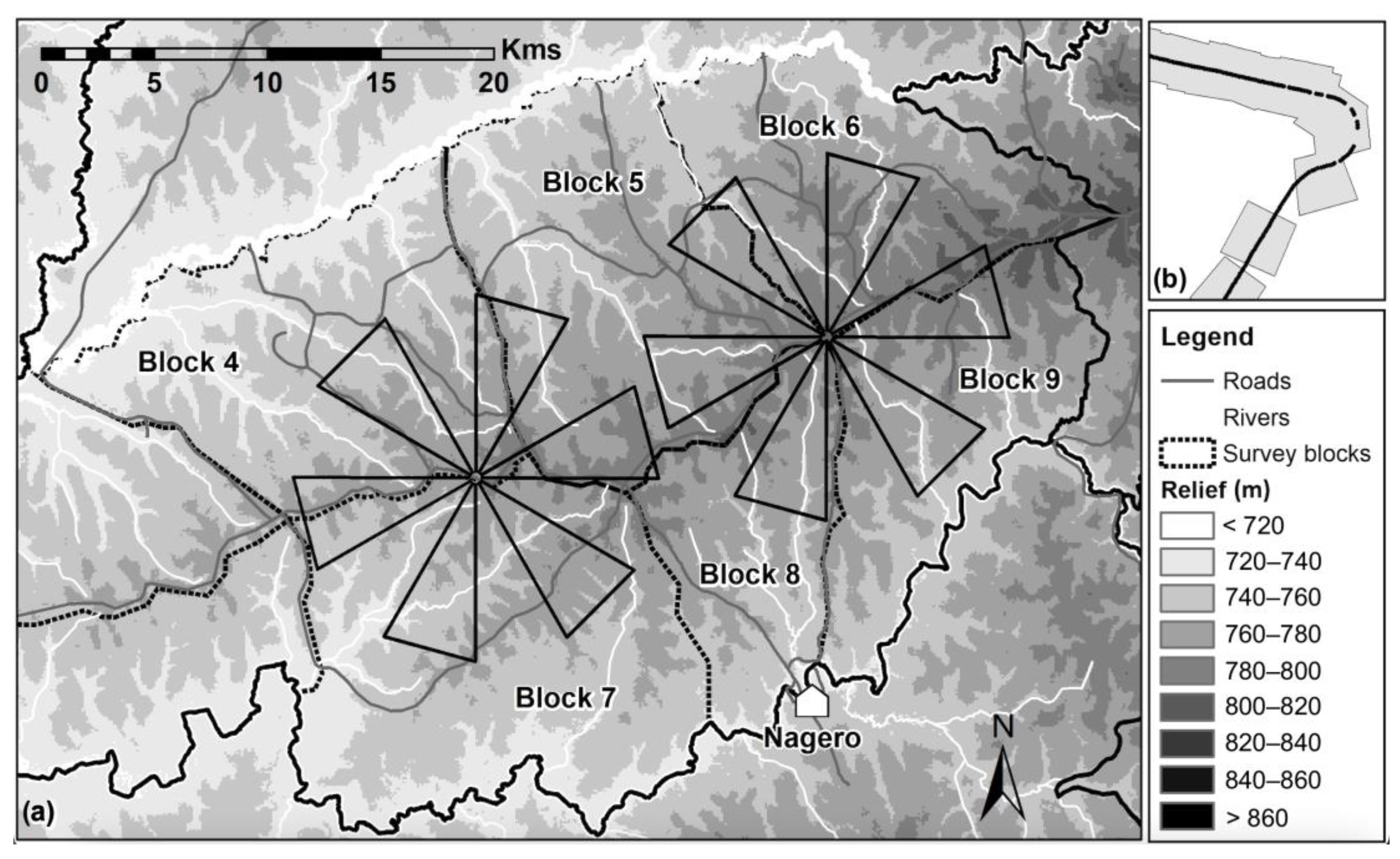
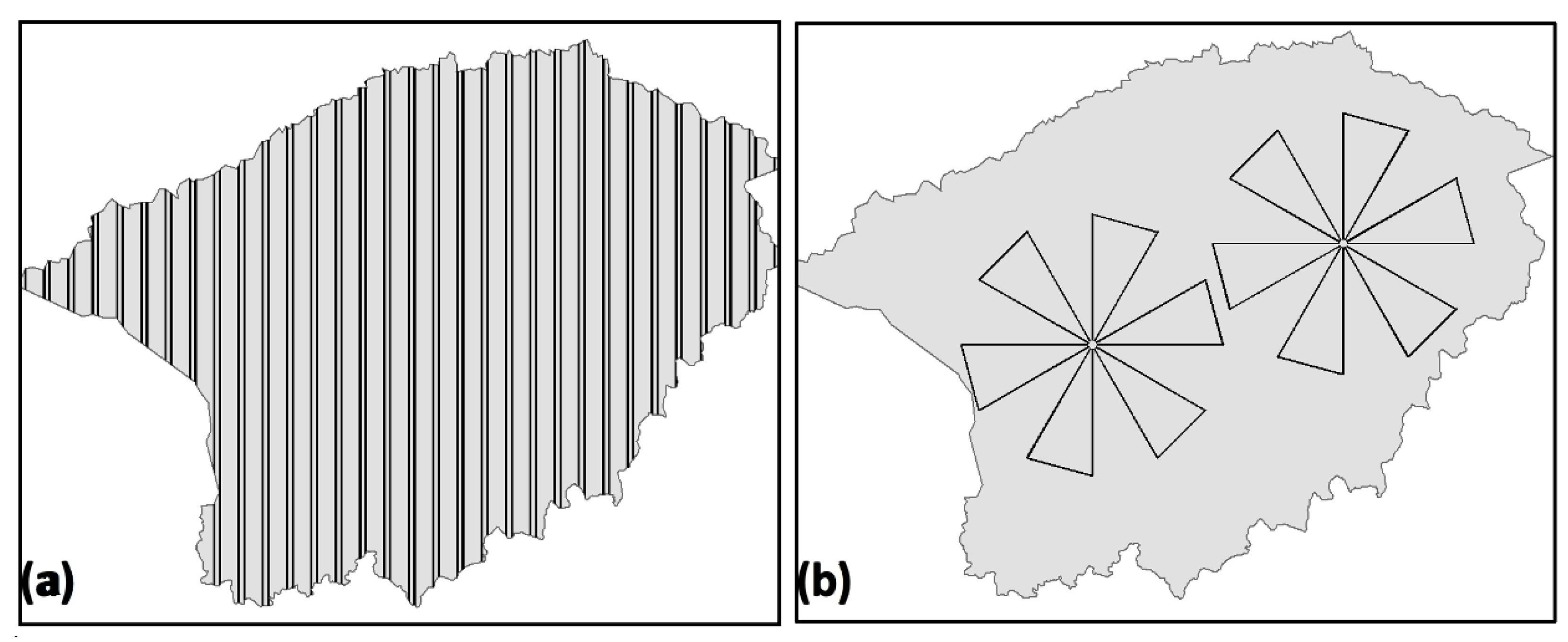
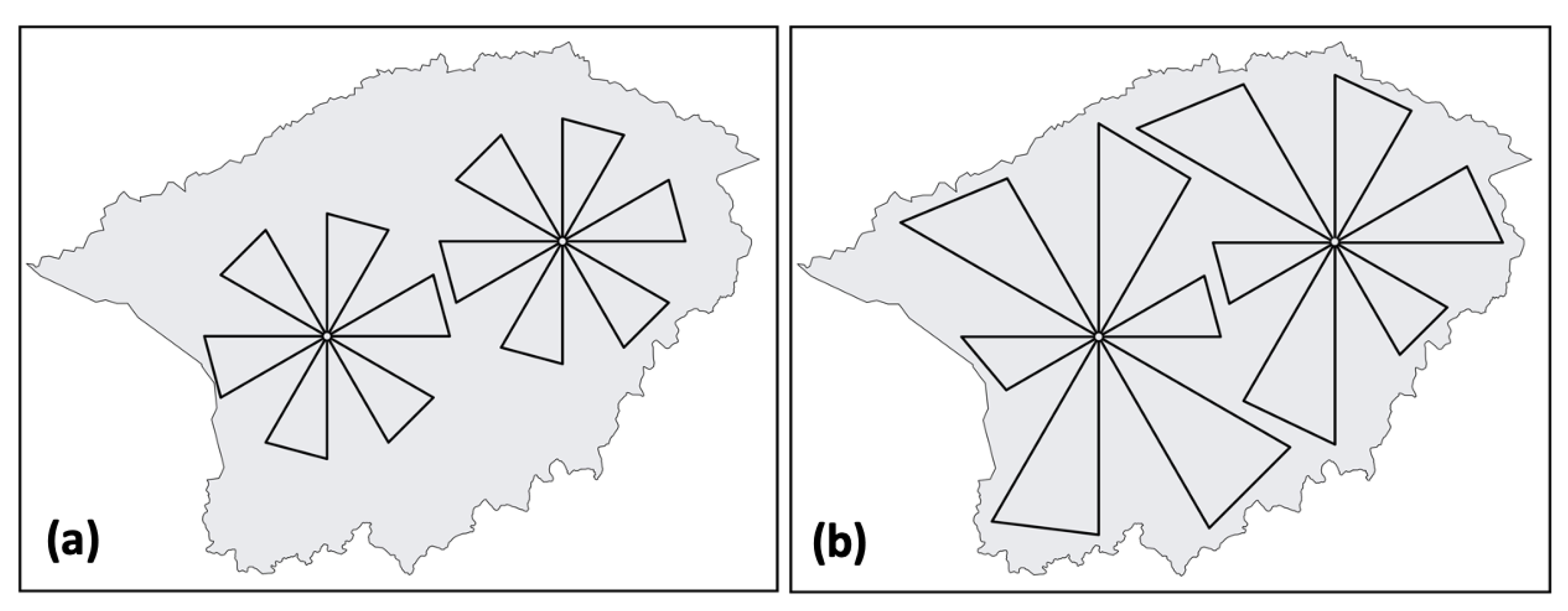
2.3. Flight Plan Design
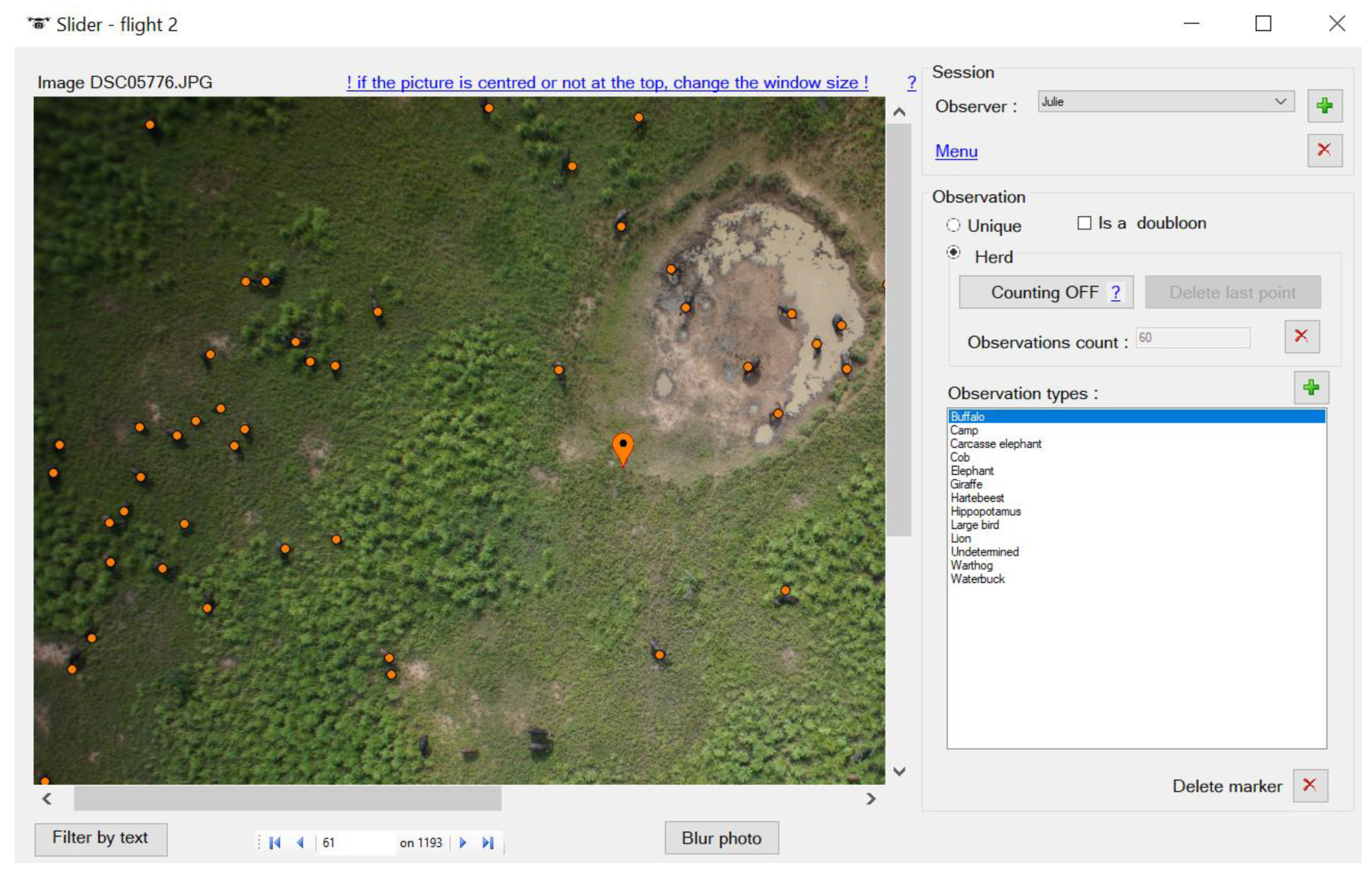
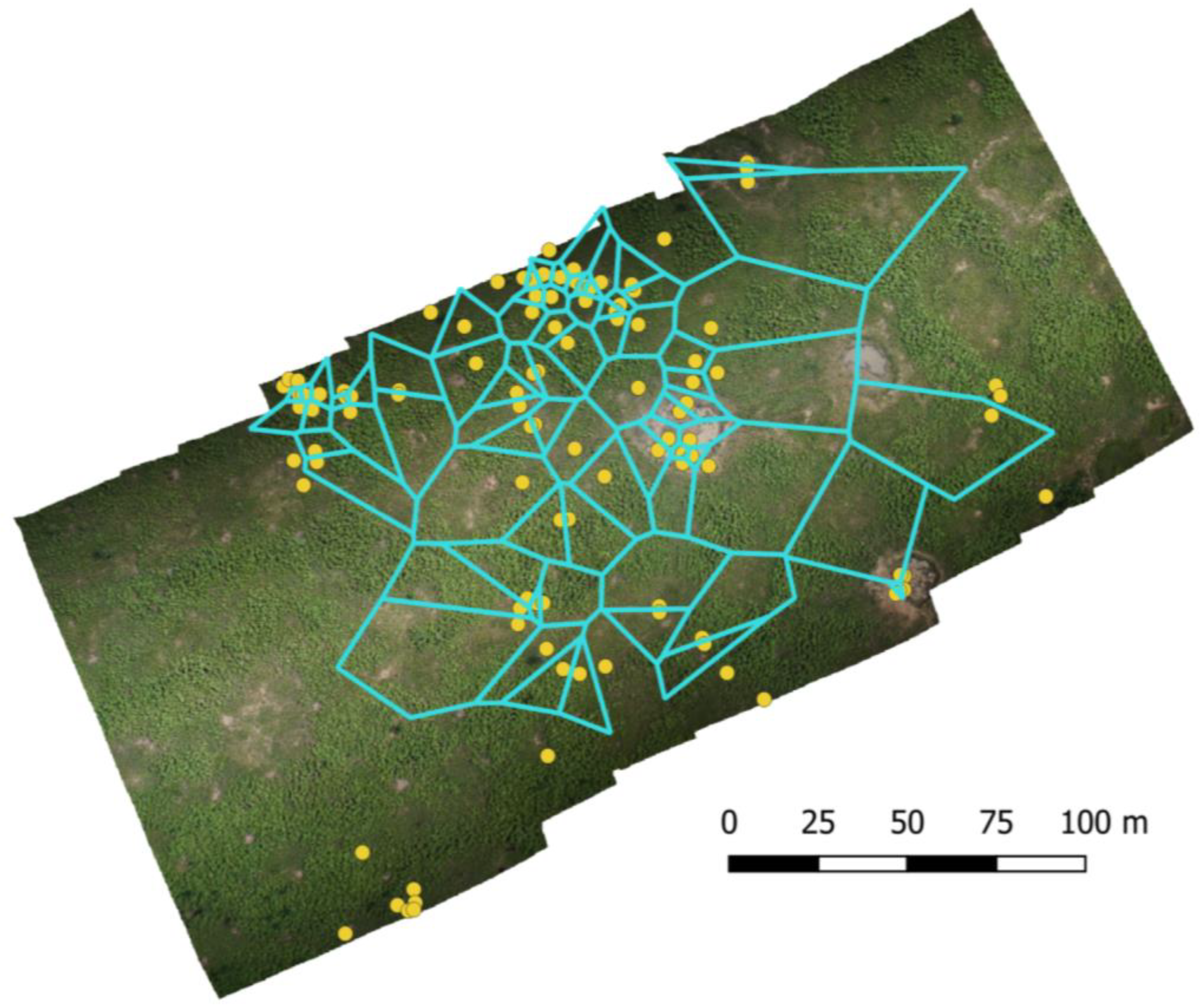
2.4. UAS Data Collection and Image Processing
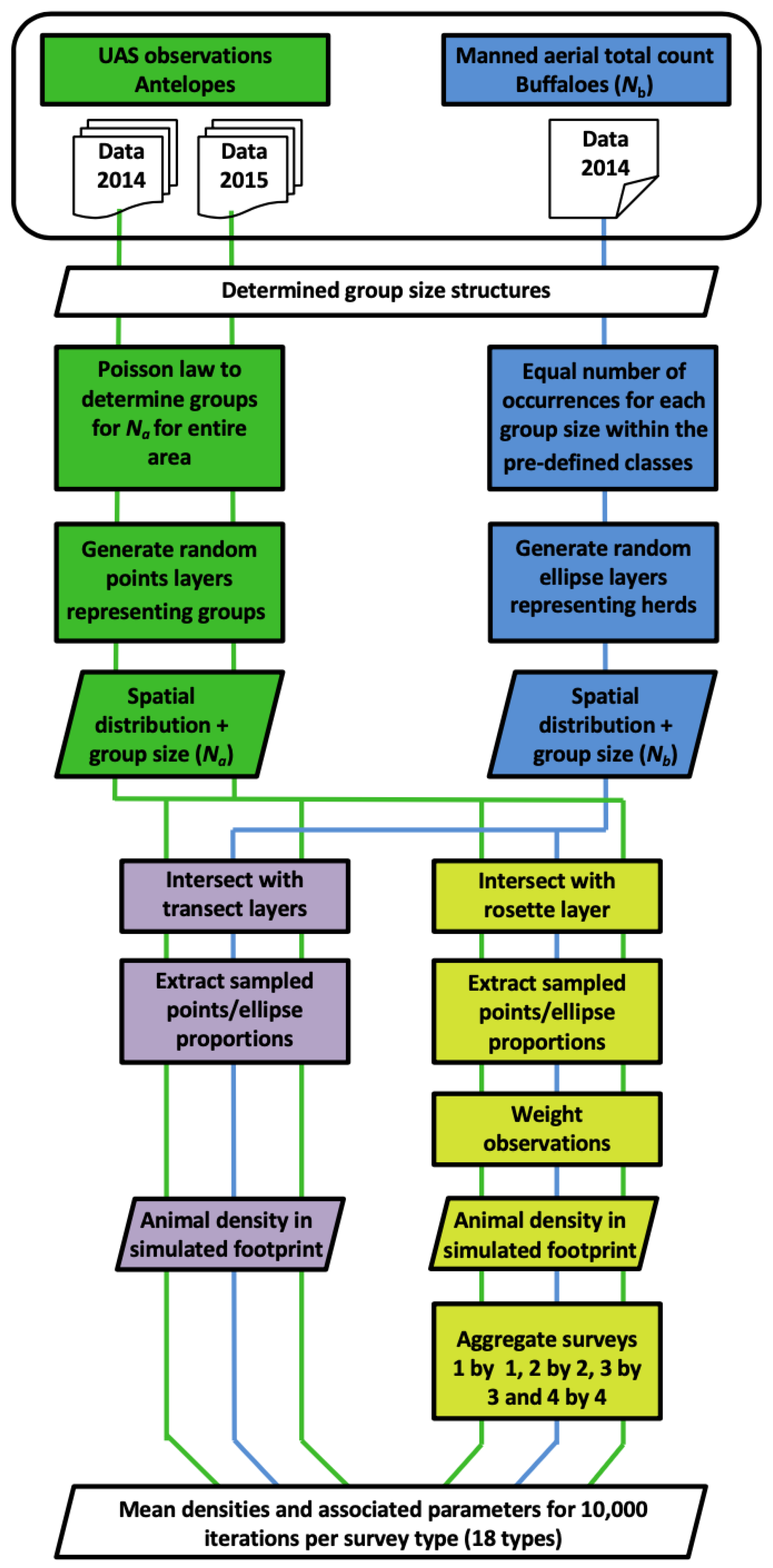
2.5. Simulations and Comparison with the Traditional Transect Method
2.5.1. Animal Population Structures and Distribution Layers
2.5.2. Simulated Sample Counts
2.5.3. Performance of the Flight Plans
3. Results
3.1. UAS Data Collection and Image Processing
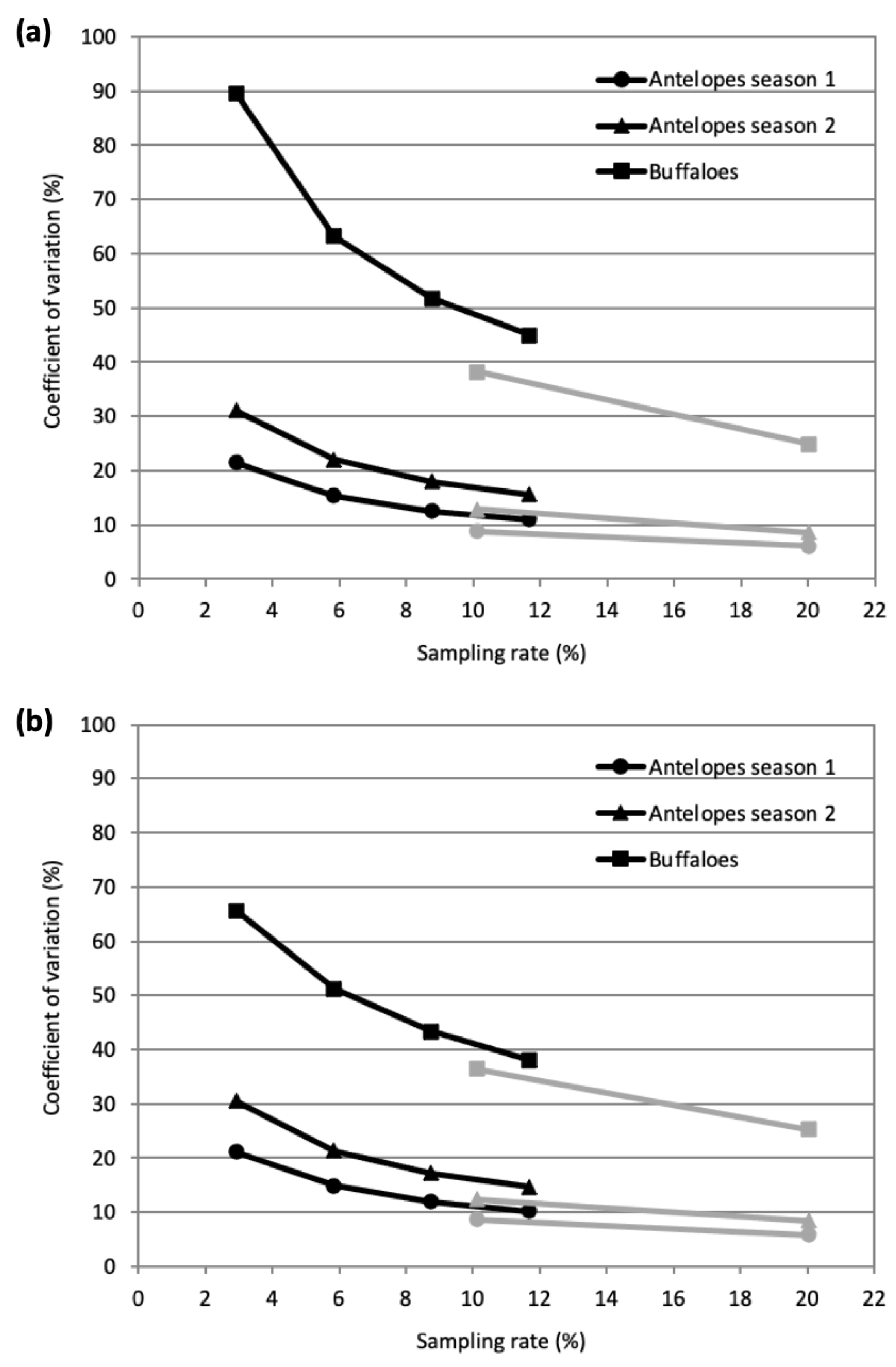
3.2. Simulations and Comparison with the Traditional Transect Method
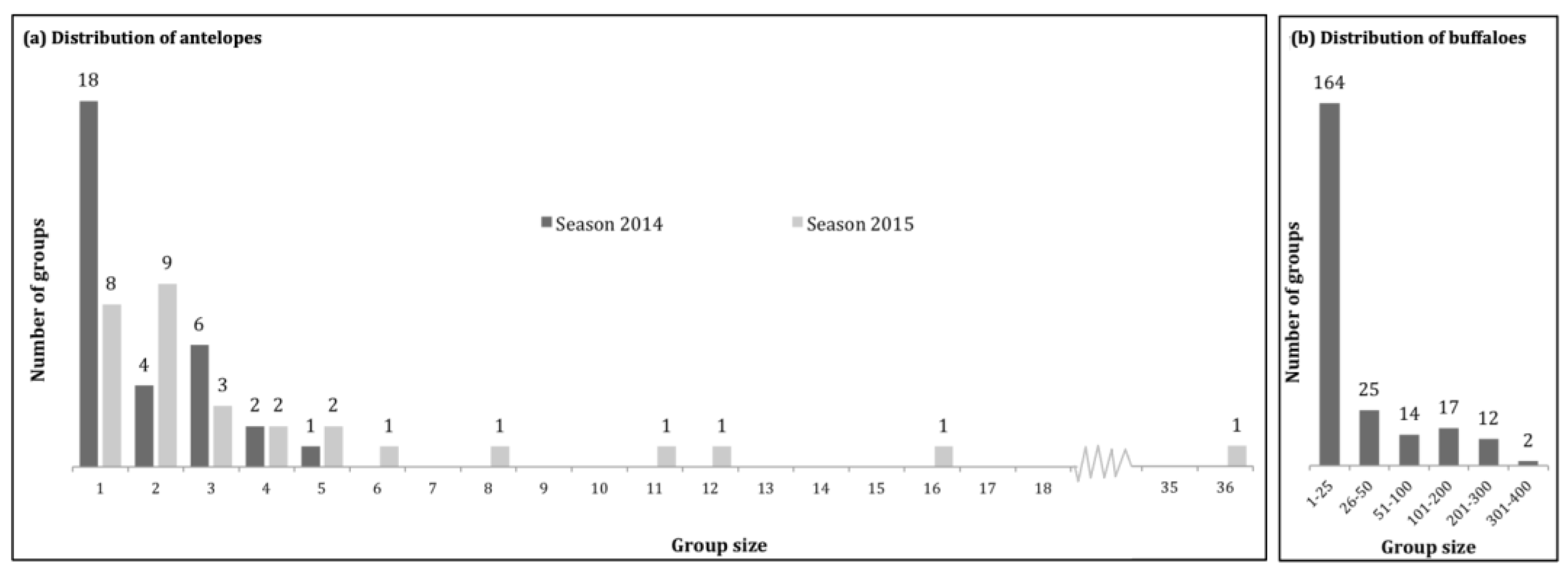
3.2.1. Population Structures and Distribution Layers
3.2.2. Simulated Sample Counts and Performance
4. Discussion
4.1. Rosette Flight Plan
4.2. Simulated Survey Results
4.3. Performance of the Rosette Flight Plan in the Field
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2014, Late Rainy Season | 2015, Early Rainy Season | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rosette 1 | Rosette 2 | Rosette 1 | Rosette 2 | |||||||||||
| Flight Number | R1F1 | R1F2 | R1F3 | R2F1 | R2F2 | R2F3 | R1F1 | R1F2 | R1F3 | R2F1 | R2F2 | * R2F3a | * R2F3b | |
| Flight Time (min) | 50 | 45 | 45 | 50 | 45 | 50 | 50 | 55 | 55 | 50 | 45 | 50 | 25 | |
| Conditions and camera parameters | Weather | Cloudy, wind 2–6 m/s | Cloudy, stormy, wind 3–6 m/s | Sunny to partially cloudy, wind 2–5 m/s | Sunny to partially cloudy, wind 2–5 m/s | |||||||||
| ISO | 200 | 400 | 200 | 200 | ||||||||||
| Shutter speed | 1600 | 2000 | 2000 | 2000 | ||||||||||
| Trigger distance (m) | 12 | 16 | 25 | 25 | ||||||||||
| Photos | Total number | 784 | 622 | 587 | 1300 | 993 | 1087 | 1048 | 1330 | 1316 | 1372 | 1382 | 981 | 670 |
| Discarded | 136 | 90 | 71 | 130 | 80 | 179 | 83 | 132 | 86 | 96 | 138 | 377 | 88 | |
| Blurry | 3 | 2 | 3 | 15 | 7 | 3 | 4 | 5 | 8 | 18 | 20 | 5 | 5 | |
| Reviewed for animals | 645 | 530 | 513 | 1155 | 906 | 905 | 961 | 1193 | 1222 | 1258 | 1224 | 599 | 577 | |
Appendix B

Appendix C

Appendix D

- d: population density (animal/km2)
- : weighted number of animals inside a rosette
- area: area of the sample bands (km2)
- m: number of groups of animals inside the sample bands
- : number of animals of group I that are inside the sample bands
- : weighting factor for group i
- : distance between group I and the rosette center (m)
- np: number of petals in the rosette
- bw: width of the bands surveyed by the UAV (m).
- : mean weight considering an entire rosette.
- : in the case of tangential segments, the weighting considers the fact that these segments represent approximately half the circumference of the circle in which the rosette is inserted.
- was computed by numerical integration using raster calculator tool in QGIS. The value of 14.32 has been obtained considering the petal configuration presented in this study.
Appendix E
| Sampling Strip Area (km2) | Nbr Antelopes | Nbr Giraffes | Nbr Buffaloes | Nbr Warthogs | NBR Hippos | ||
|---|---|---|---|---|---|---|---|
| 2014 | Rosette 1 | 12.37 | 30 | / | 249 | 9 | 23 |
| Rosette 2 | 12.13 | 27 | / | 10 | 27 | 35 | |
| Total | 24.50 | 57 | / | 259 | 36 | 58 | |
| Density (n/km2) | 2.33 | / | 10.57 | 1.47 | 2.37 | ||
| Estimated total population | 2252 | / | 10,231 | 1422 | 2291 | ||
| Standard error (n/km2) | 136.41 | / | 13,210.42 | 1025.30 | 702.14 | ||
| CI 95% (n/km2) | ±189 | / | ±18,308 | ±1421 | ±973 | ||
| (%) | ±8.40 | / | ±178.96 | ±99.93 | ±42.48 | ||
| 2015 | Rosette 1 | 17.50 | 50 | 10 | 133 | 16 | 47 |
| Rosette 2 | 15.94 | 92 | 0 | 47 | 7 | 31 | |
| Total | 33.44 | 142 | 10 | 180 | 23 | 78 | |
| Density (n/km2) | 4.25 | 0.30 | 5.38 | 0.69 | 2.33 | ||
| Estimated total population | 4109 | 289 | 5209 | 666 | 2257 | ||
| Standard error (n/km2) | 1994.40 | 391.03 | 3182.99 | 325.14 | 507.01 | ||
| CI 95% (n/km2) | ±2764 | ±542 | ±4411 | ±451 | ±703 | ||
| (%) | ±67.26 | ±187.26 | ±84.68 | ±67.70 | ±31.13 | ||
Appendix F

References
- Jachmann, H. Estimating Abundance of African Wildlife: An Aid to Adaptive Management; Kluwer Academic Publishers: Boston, MA, USA, 2001; ISBN 978-1-4615-1381-0. [Google Scholar]
- Jachmann, H. Evaluation of Four Survey Methods for Estimating Elephant Densities. Afr. J. Ecol. 1991, 29, 188–195. [Google Scholar] [CrossRef]
- Wang, D.; Shao, Q.; Yue, H. Surveying Wild Animals from Satellites, Manned Aircraft and Unmanned Aerial Systems (UASs): A Review. Remote Sens. 2019, 11, 1308. [Google Scholar] [CrossRef] [Green Version]
- Bouché, P.; Lejeune, P.; Vermeulen, C. How to Count Elephants in West African Savannahs? Synthesis and Comparison of Main Gamecount Methods. Biotechnol. Agron. Sociol. Environ. 2012, 16, 77–91. [Google Scholar]
- Dunham, K.M. Trends in Populations of Elephant and Other Large Herbivores in Gonarezhou National Park, Zimbabwe, as Revealed by Sample Aerial Surveys. Afr. J. Ecol. 2012, 50, 476–488. [Google Scholar] [CrossRef]
- Watts, A.C.; Perry, J.H.; Smith, S.E.; Burgess, M.A.; Wilkinson, B.E.; Szantoi, Z.; Ifju, P.G.; Percival, H.F. Small Unmanned Aircraft Systems for Low-Altitude Aerial Surveys. J. Wildl. Manag. 2010, 74, 1614–1619. [Google Scholar] [CrossRef]
- Christie, K.S.; Gilbert, S.L.; Brown, C.L.; Hatfield, M.; Hanson, L. Unmanned Aircraft Systems in Wildlife Research: Current and Future Applications of a Transformative Technology. Front. Ecol. Environ. 2016, 14, 241–251. [Google Scholar] [CrossRef]
- Sasse, D.B. Job-Related Mortality of Wildlife Workers in the United States, 1937–2000. Wildl. Soc. Bull. 2003, 31, 1015–1020. [Google Scholar]
- Mulero-Pázmány, M.; Stolper, R.; van Essen, L.D.; Negro, J.J.; Sassen, T. Remotely Piloted Aircraft Systems as a Rhinoceros Anti-Poaching Tool in Africa. PLoS ONE 2014, 9, e83873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, D.W. Unoccupied Aircraft Systems in Marine Science and Conservation. Annu. Rev. Mar. Sci. 2019, 11, 439–463. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.; Bird, D.M. Wildlife Research and Management Methods in the 21st Century: Where Do Unmanned Aircraft Fit In? J. Unmanned Veh. Syst. 2015, 3, 137–155. [Google Scholar] [CrossRef] [Green Version]
- Linchant, J.; Lisein, J.; Ngabinzeke, J.S.; Lejeune, P.; Vermeulen, C. Are Unmanned Aircraft Systems (UASs) the Future of Wildlife Monitoring? A Review of Accomplishments and Challenges. Mammal Rev. 2015, 45, 239–252. [Google Scholar] [CrossRef]
- López, J.J.; Mulero-Pázmány, M. Drones for Conservation in Protected Areas: Present and Future. Drones 2019, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, A.; Kelly, N.; Peel, D. Unmanned Aerial Vehicles (UAVs) for Surveying Marine Fauna: A Dugong Case Study. PLoS ONE 2013, 8, e79556. [Google Scholar] [CrossRef] [Green Version]
- Michez, A.; Broset, S.; Lejeune, P. Ears in the Sky: Potential of Drones for the Bioacoustic Monitoring of Birds and Bats. Drones 2021, 5, 9. [Google Scholar] [CrossRef]
- Kloepper, L.N.; Kinniry, M. Recording Animal Vocalizations from a UAV: Bat Echolocation during Roost Re-Entry. Sci. Rep. 2018, 8, 7779. [Google Scholar] [CrossRef] [Green Version]
- Desrochers, A.; Tremblay, J.A.; Aubry, Y.; Chabot, D.; Pace, P.; Bird, D.M. Estimating Wildlife Tag Location Errors from a VHF Receiver Mounted on a Drone. Drones 2018, 2, 44. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Segaran, R.R.; Reid, I.; Terauds, A.; Koh, L.P. Drones Count Wildlife More Accurately and Precisely than Humans. Methods Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Rees, A.F.; Avens, L.; Ballorain, K.; Bevan, E.; Broderick, A.C.; Carthy, R.R.; Christianen, M.J.A.; Duclos, G.; Heithaus, M.R.; Johnston, D.W.; et al. The Potential of Unmanned Aerial Systems for Sea Turtle Research and Conservation: A Review and Future Directions. Endanger. Species Res. 2018, 35, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Morley, C.G.; Broadley, J.; Hartley, R.; Herries, D.; McMorran, D.; McLean, I.G. The Potential of Using Unmanned Aerial Vehicles (UAV) for Precision Pest Control of Possums (Trichosurus Vulpeca). Rethink. Ecol. 2017, 2, 27–39. [Google Scholar] [CrossRef]
- van Andel, A.C.; Wich, S.A.; Boesch, C.; Koh, L.P.; Robbins, M.M.; Kelly, J.; Kuehl, H.S. Locating Chimpanzee Nests and Identifying Fruiting Trees with an Unmanned Aerial Vehicle. Am. J. Primatol. 2015, 77, 1122–1134. [Google Scholar] [CrossRef] [PubMed]
- Fust, P.; Loos, J. Development Perspectives for the Application of Autonomous, Unmanned Aerial Systems (UASs) in Wildlife Conservation. Biol. Conserv. 2019, 241, 108380. [Google Scholar] [CrossRef]
- Butcher, P.A.; Colefax, A.P.; Gorkin, R.A., III; Kajiura, S.M.; López, N.A.; Mourier, J.; Purcell, C.R.; Skomal, G.B.; Tucker, J.P.; Walsh, A.J.; et al. The Drone Revolution of Shark Science: A Review. Drones 2021, 5, 8. [Google Scholar] [CrossRef]
- Inman, V.L.; Kingsford, R.T.; Chase, M.J.; Leggett, K.E.A. Drone-Based Effective Counting and Ageing of Hippopotamus (Hippopotamus Amphibius) in the Okavango Delta in Botswana. PLoS ONE 2019, 14, e0219652. [Google Scholar] [CrossRef] [Green Version]
- Gooday, O.J.; Key, N.; Goldstien, S.; Zawar-Reza, P. An Assessment of Thermal-Image Acquisition with an Unmanned Aerial Vehicle (UAV) for Direct Counts of Coastal Marine Mammals Ashore. J. Unmanned Veh. Syst. 2018, 6, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Linchant, J.; Lhoest, S.; Quevauvillers, S.; Lejeune, P.; Vermeulen, C.; Ngabinzeke, J.S.; Belanganayi, B.L.; Delvingt, W.; Bouché, P. UAS Imagery Reveals New Survey Opportunities for Counting Hippos. PLoS ONE 2018, 13, e0206413. [Google Scholar] [CrossRef]
- Ratcliffe, N.; Guihen, D.; Robst, J.; Crofts, S.; Stanworth, A.; Enderlein, P. A Protocol for the Aerial Survey of Penguin Colonies Using UAVs. J. Unmanned Veh. Syst. 2015, 3, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.; Bird, D.M. Evaluation of an Off-the-Shelf Unmanned Aircraft System for Surveying Flocks of Geese. Waterbirds 2012, 35, 170–174. [Google Scholar] [CrossRef]
- Hodgson, J.C. Using Drones to Improve Wildlife Monitoring in a Changing Climate. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 2020. [Google Scholar]
- Fiori, L.; Doshi, A.; Martinez, E.; Orams, M.B.; Bollard-Breen, B. The Use of Unmanned Aerial Systems in Marine Mammal Research. Remote Sens. 2017, 9, 543. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, C.; Lejeune, P.; Lisein, J.; Sawadogo, P.; Bouché, P. Unmanned Aerial Survey of Elephants. PLoS ONE 2013, 8, e54700. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Shao, Q.; Jiang, Z. A Population Census of Large Herbivores Based on UAV and Its Effects on Grazing Pressure in the Yellow-River-Source National Park, China. Int. J. Environ. Res. Public Health 2019, 16, 4402. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Shao, Q.; Li, Y.; Wang, Y.; Wang, D.; Liu, J.; Fan, J.; Yang, F. Application of UAV Remote Sensing for a Population Census of Large Wild Herbivores—Taking the Headwater Region of the Yellow River as an Example. Remote Sens. 2018, 10, 1041. [Google Scholar] [CrossRef] [Green Version]
- Beaver, J.T.; Baldwin, R.W.; Messinger, M.; Newbolt, C.H.; Ditchkoff, S.S.; Silman, M.R. Evaluating the Use of Drones Equipped with Thermal Sensors as an Effective Method for Estimating Wildlife. Wildl. Soc. Bull. 2020, 44, 434–443. [Google Scholar] [CrossRef]
- Gentle, M.; Finch, N.; Speed, J.; Pople, A. A Comparison of Unmanned Aerial Vehicles (Drones) and Manned Helicopters for Monitoring Macropod Populations. Wildl. Res. 2018, 45, 586–594. [Google Scholar] [CrossRef]
- Strindberg, S.; Buckland, S.T. Zigzag Survey Designs in Line Transect Sampling. J. Agric. Biol. Environ. Stat. 2004, 9, 443–461. [Google Scholar] [CrossRef]
- Barreto, J.; Cajaíba, L.; Teixeira, J.B.; Nascimento, L.; Giacomo, A.; Barcelos, N.; Fettermann, T.; Martins, A. Drone-Monitoring: Improving the Detectability of Threatened Marine Megafauna. Drones 2021, 5, 14. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Baylis, S.M.; Mott, R.; Herrod, A.; Clarke, R.H. Precision Wildlife Monitoring Using Unmanned Aerial Vehicles. Sci. Rep. 2016, 6, 22574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, G.M. Sampling Methods for Aerial Censuses of Wildlife Populations. East Afr. Agric. For. J. 1969, 34, 46–49. [Google Scholar] [CrossRef]
- Fritsch, C.; Downs, C. Evaluation of Low-Cost Consumer-Grade UAVs for Conducting Comprehensive High-Frequency Population Censuses of Hippopotamus Populations. Conserv. Sci. Pract. 2020, 2, e281. [Google Scholar] [CrossRef]
- Bushaw, J.D.; Ringelman, K.M.; Rohwer, F.C. Applications of Unmanned Aerial Vehicles to Survey Mesocarnivores. Drones 2019, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Balimbaki, A. Etude Socio-Économique dans les Trois Domaines de Chasse Contigus au Parc National de la Garamba; Institut Congolais pour la Conservation de la Nature and African Parks Network: Parc National de la Garamba, Democratic Republic of the Congo, 2015; p. 76. [Google Scholar]
- Hillman-Smith, A.K. Garamba National Park, Hippo Count, March 1988; Institut Congolais pour la Conservation de la Nature: Garamba National Park, Democratic Republic of the Congo, 1988. [Google Scholar]
- De Saeger, H.; Baert, P.; de Moulin, G.; Denisoff, I.; Martin, J.; Micha, M.; Noirfalise, A.; Schoemaker, P.; Troupin, G.; Verschuren, J. Introduction. In Exploration du Parc National de la Garamba. Fascicule 1; Institut des Parcs Nationaux du Congo Belge: Bruxelles, Belgium, 1954. [Google Scholar]
- Linchant, J.; Lhoest, S.; Quevauvillers, S.; Semeki, J.; Lejeune, P.; Vermeulen, C. WiMUAS: A Tool to Review Wildlife Data from Various Flight Plans. In Proceedings of the ISPRS—International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences, La Grand Motte, France, 28 September–3 October 2015; Volume XL-3/W3. [Google Scholar]
- Norton-Griffiths, M. Counting Animals. In Serengeti Ecological Monitoring Programme; African Wildlife Leadership Foundation: Nairobi, Kenya, 1978. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Griesser, M.; Ma, Q.; Webber, S.; Bowgen, K.; Sumpter, D.J.T. Understanding Animal Group-Size Distributions. PLoS ONE 2011, 6, e23438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegel, T.M.; Cushman, S.A.; Evans, J.; Huettmann, F. Chapter 16: Current State of the Art for Statistical Modelling of Species Distributions. In Spatial Complexity, Informatics, and Wildlife Conservation; Cushman, S.A., Huettmann, F., Eds.; Springer: New York, NY, USA, 2010; pp. 273–311. [Google Scholar]
- Mònico, M. Garamba National Park Aerial Survey March 2014; Institut Congolais pour la Conservation de la Nature and African Parks Network: Parc National de la Garamba, Democratic Republic of the Congo, 2014. [Google Scholar]
- CARMA Network. Monitoring Rangifer herds (population dynamics): MANUAL; Gunn, R., Russel, D., Eds.; CircumArctic Rangifer Monitoring and Assessment (CARMA) Network: Vancouver, BC, Canada, 2008; 50p. [Google Scholar]
- Ferreira, S.M.; Aarde, R.J. Aerial survey intensity as a determinant of estimates of African elephant population sizes and trends. S. Afr. J. Wildl. Res. 2009, 39, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Kabir, R.H.; Lee, K. Wildlife monitoring using a multi-UAV system with optimal transport theory. Appl. Sci. 2021, 11, 4070. [Google Scholar] [CrossRef]
- Eikelboom, J.A.J.; Wind, J.; van de Ven, E.; Kenana, L.M.; Schroder, B.; de Knegt, H.J.; van Langevelde, F.; Prins, H.H.T. Improving the Precision and Accuracy of Animal Population Estimates with Aerial Image Object Detection. Methods Ecol. Evol. 2019, 10, 1875–1887. [Google Scholar] [CrossRef] [Green Version]
- Maes, W.H.; Huete, A.R.; Steppe, K. Optimizing the Processing of UAV-Based Thermal Imagery. Remote Sens. 2017, 9, 476. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Ballard, G.; Schmidt, A.; Schwager, M. Multidrone aerial surveys of penguin colonies in Antarctica. Sci. Robot. 2020, 5, eabc3000. [Google Scholar] [CrossRef]
- Moreni, M.; Theau, J.; Foucher, S. Train Fast While Reducing False Positives: Improving Animal Classification Performance Using Convolutional Neural Networks. Geomatics 2021, 1, 34–49. [Google Scholar] [CrossRef]
- Kellenberger, B.; Marcos, D.; Tuia, D. Detecting Mammals in UAV Images: Best Practices to Address a Substantially Imbalanced Dataset with Deep Learning. Remote Sens. Environ. 2018, 216, 139–153. [Google Scholar] [CrossRef] [Green Version]
- Lenzi, J.; Barnas, A.F.; ElSaid, A.A.; Dessel, T.; Rockwell, R.F.; Ellis, S.N. Artificial intelligence for automated detection of large mammals creates path to upscale drone surveys. Sci. Rep. 2023, 13, 947. [Google Scholar] [CrossRef]
- Delplanque, A.; Foucher, S.; Théau, J.; Bussière, E.; Vermeulen, C.; Lejeune, P. From crowd to herd counting: How to precisely detect and count African mammals using aerial imagery and deep learning? ISPRS J. Photogramm. Remote Sens. 2023, 197, 167–180. [Google Scholar] [CrossRef]







| Animal Distribution (Season) | Survey Flight Plan | Coverage Type | Sampling Rate (%) | Animal Density (n/km2) | CV (%) | Biais (%) | t-Test | Jolly 2 | |
|---|---|---|---|---|---|---|---|---|---|
| Mean | Standard Deviation | p-Value | CV (%) | ||||||
| 2014 | Transects | 1.5 km | 20.04 | 3.288 | 0.196 | 6.00 | −0.02 | 0.689 | 5.89 |
| 3 km | 10.11 | 3.291 | 0.291 | 8.80 | 0.05 | 0.610 | 8.63 | ||
| Rosettes | 1 rep | 2.95 | 3.300 | 0.710 | 21.50 | 0.35 | 0.109 | 21.11 | |
| 2 rep | 5.90 | 3.299 | 0.505 | 15.30 | 0.29 | 0.056 | 14.86 | ||
| 3 rep | 8.85 | 3.297 | 0.412 | 12.50 | 0.24 | 0.057 | 11.99 | ||
| 4 rep | 11.80 | 3.295 | 0.359 | 10.91 | 0.18 | 0.098 | 10.23 | ||
| 2015 | Transects | 1.5 km | 20.04 | 3.289 | 0.282 | 8.60 | −0.01 | 0.890 | 8.41 |
| 3 km | 10.11 | 3.290 | 0.420 | 12.80 | 0.03 | 0.834 | 12.44 | ||
| Rosettes | 1 rep | 2.95 | 3.297 | 1.023 | 31.02 | 0.24 | 0.440 | 30.58 | |
| 2 rep | 5.90 | 3.301 | 0.726 | 22.01 | 0.35 | 0.114 | 21.38 | ||
| 3 rep | 8.85 | 3.303 | 0.592 | 17.91 | 0.44 | 0.016 | 17.18 | ||
| 4 rep | 11.80 | 3.302 | 0.512 | 15.51 | 0.38 | 0.014 | 14.64 | ||
| Animal Distribution (Season) | Survey Flight Plan | Coverage Type | Sampling Rate (%) | Animal Density (n/km2) | CV (%) | Bias (%) | t-Test | Jolly 2 | |
|---|---|---|---|---|---|---|---|---|---|
| Mean | Standard Deviation | p-Value | CV (%) | ||||||
| 2014 | Transects | 1.5 km | 20.04 | 4.964 | 1.230 | 24.80 | −0.28 | 0.260 | 25.30 |
| 3 km | 10.11 | 4.955 | 1.893 | 38.20 | −0.45 | 0.239 | 36.45 | ||
| Rosettes | 1 rep | 2.95 | 4.997 | 4.474 | 89.50 | 0.38 | 0.671 | 65.61 | |
| 2 rep | 5.90 | 5.022 | 3.175 | 63.20 | 0.90 | 0.159 | 51.19 | ||
| 3 rep | 8.85 | 5.022 | 2.598 | 51.70 | 0.90 | 0.086 | 43.35 | ||
| 4 rep | 11.80 | 5.013 | 2.257 | 45.00 | 0.72 | 0.113 | 38.04 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linchant, J.; Lejeune, P.; Quevauvillers, S.; Vermeulen, C.; Brostaux, Y.; Lhoest, S.; Michez, A. Evaluation of an Innovative Rosette Flight Plan Design for Wildlife Aerial Surveys with UAS. Drones 2023, 7, 208. https://doi.org/10.3390/drones7030208
Linchant J, Lejeune P, Quevauvillers S, Vermeulen C, Brostaux Y, Lhoest S, Michez A. Evaluation of an Innovative Rosette Flight Plan Design for Wildlife Aerial Surveys with UAS. Drones. 2023; 7(3):208. https://doi.org/10.3390/drones7030208
Chicago/Turabian StyleLinchant, Julie, Philippe Lejeune, Samuel Quevauvillers, Cédric Vermeulen, Yves Brostaux, Simon Lhoest, and Adrien Michez. 2023. "Evaluation of an Innovative Rosette Flight Plan Design for Wildlife Aerial Surveys with UAS" Drones 7, no. 3: 208. https://doi.org/10.3390/drones7030208