
Human Schistosomiasis Vaccines as Next Generation Control Tools

{kind=link}
1. Introduction
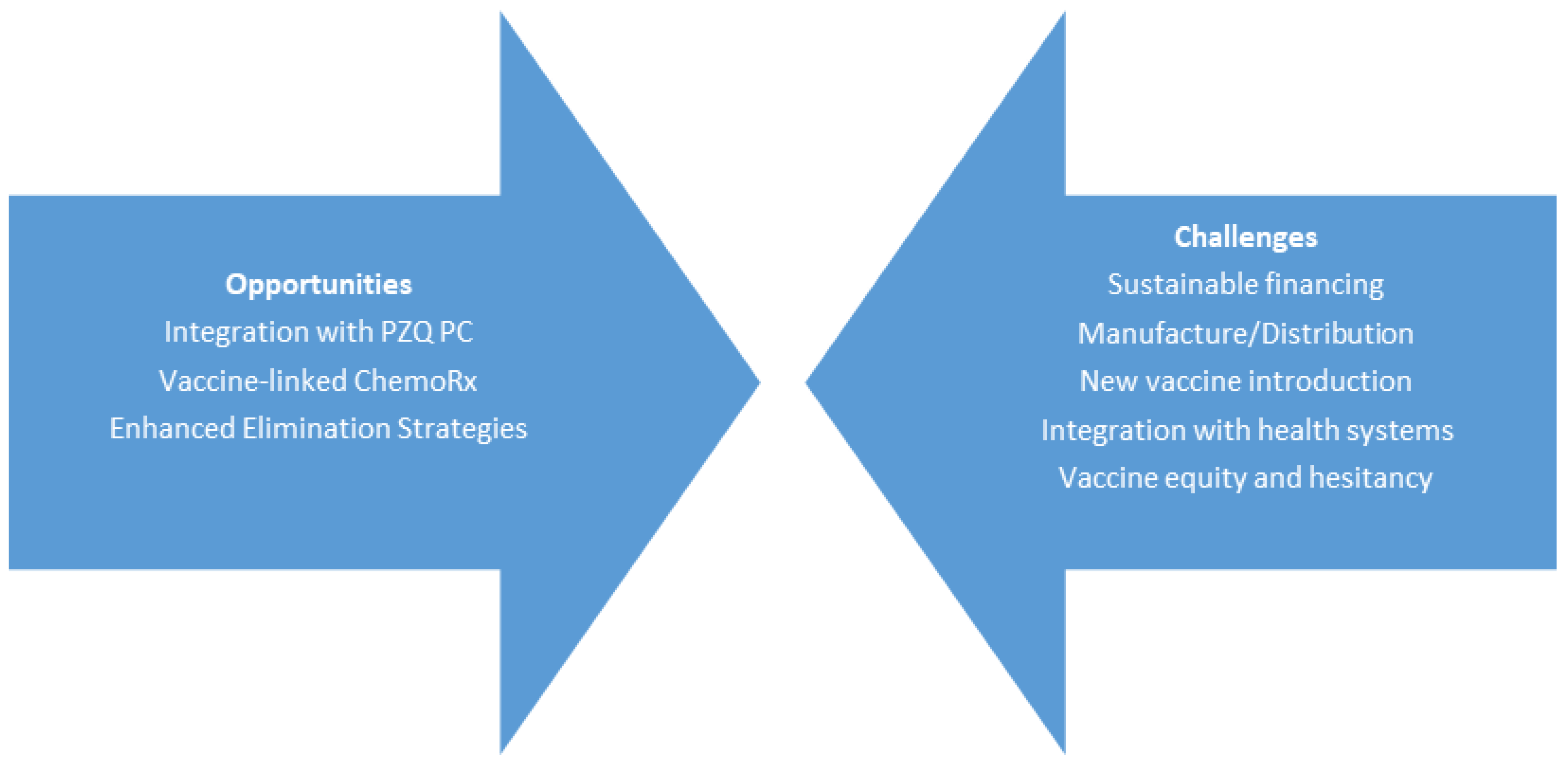
2. Opportunities for “Vaccine-Linked Chemotherapy”
3. The Current Pipeline
4. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Institute of Health Metrics and Evaluation—University of Washington. Global Burden of Disease Study 2019. Schistosomiasis —Level 3 Cause. Available online: https://www.healthdata.org/results/gbd_summaries/2019/schistosomiasis-level-3-cause (accessed on 6 February 2023).
- King, C.H.; Galvani, A.P. Underestimation of the global burden of schistosomiasis. Lancet 2018, 391, 307–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotez, P.J.; Engels, D.; Gyapong, M.; Ducker, C.; Malecela, M.N. Female Genital Schistosomiasis. N. Engl. J. Med. 2019, 381, 2493–2495. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J. Malnutrition vaccines for an imminent global food catastrophe. Trends Pharmacol. Sci. 2022, 43, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, D.H.; Asamoa-Bah, A.; Fenwick, A.; Savioli, L.; Hotez, P. The history of the neglected tropical disease movement. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 169–175. [Google Scholar] [CrossRef]
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Kumaresan, J.; Sachs, S.E.; Sachs, J.D.; Savioli, L. Control of neglected tropical diseases. N. Engl. J. Med. 2007, 357, 1018–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotez, P.J.; Fenwick, A.; Molyneux, D.H. Collateral Benefits of Preventive Chemotherapy—Expanding the War on Neglected Tropical Diseases. N. Engl. J. Med. 2019, 380, 2389–2391. [Google Scholar] [CrossRef]
- Kura, K.; Hardwick, R.J.; Truscott, J.E.; Toor, J.; Hollingsworth, T.D.; Anderson, R.M. The impact of mass drug administration on Schistosoma haematobium infection: What is required to achieve morbidity control and elimination? Parasites Vectors 2020, 13, 554. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Schistosomiasis and soil transmitted helminthiases: Progress report, 2021. Weekly Epidemiol. Rec. 2022, 97, 621–631. [Google Scholar]
- World Health Organization. 34th meeting of the International Task Force for Disease Eradication, 19–20 September 2022. Weekly Epidemiol. Rec. 2023, 98, 41–52. [Google Scholar]
- Kura, K.; Ayabina, D.; Hollingsworth, T.D.; Anderson, R.M. Determining the optimal strategies to achieve elimination of transmission for Schistosoma mansoni. Parasites Vectors 2022, 15, 55. [Google Scholar] [CrossRef]
- Kura, K.; Hardwick, R.J.; Truscott, J.E.; Anderson, R.M. What is the impact of acquired immunity on the transmission of schistosomiasis and the efficacy of current and planned mass drug administration programmes? PLoS Negl. Trop. Dis. 2021, 15, e0009946. [Google Scholar] [CrossRef] [PubMed]
- Trainor-Moss, S.; Mutapi, F. Schistosomiasis therapeutics: Whats in the pipeline? Expert Rev. Clin. Pharmacol. 2016, 9, 157–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kura, K.; Collyer, B.S.; Toor, J.; Truscott, J.E.; Hollingsworth, T.D.; Keeling, M.J.; Anderson, R.M. Policy implications of the potential use of a novel vaccine to prevent infection with Schistosoma mansoni with or without mass drug administration. Vaccine 2020, 38, 4379–4386. [Google Scholar] [CrossRef]
- Kura, K.; Truscott, J.E.; Toor, J.; Anderson, R. Modelling the impact of a Schistosoma mansoni vaccine and mass drug administration to achieve morbidity control and transmission elimination. PLoS Negl. Trop. Dis. 2019, 13, e0007349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergquist, N.R.; Leonardo, L.R.; Mitchell, G.F. Vaccine-linked chemotherapy: Can schistosomiasis control benefit from an integrated approach? Trends Parasitol. 2005, 21, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Aula, O.; McManus, D.; Jones, M.; Gordon, C. Schistosomiasis with a Focus on Africa. Trop. Med. Infect. Dis. 2021, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Zoni, A.C.; Catalá, L.; Ault, S.K. Schistosomiasis Prevalence and Intensity of Infection in Latin America and the Caribbean Countries, 1942-2014: A Systematic Review in the Context of a Regional Elimination Goal. PLoS Negl. Trop. Dis. 2016, 10, e0004493. [Google Scholar] [CrossRef] [Green Version]
- Riveau, G.; Schacht, A.-M.; Dompnier, J.-P.; Deplanque, D.; Seck, M.; Waucquier, N.; Senghor, S.; Delcroix-Genete, D.; Hermann, E.; Idris-Khodja, N.; et al. Safety and efficacy of the rSh28GST urinary schistosomiasis vaccine: A phase 3 randomized, controlled trial in Senegalese children. PLoS Negl. Trop. Dis. 2018, 12, e0006968. [Google Scholar] [CrossRef] [Green Version]
- Tran, M.H.; Pearson, M.; Bethony, J.M.; Smyth, D.; Jones, M.; Duke, M.; Don, T.A.; McManus, D.; Correa-Oliveira, R.; Loukas, A. Tetraspanins on the surface of Schistosoma mansoni are protective antigens against schistosomiasis. Nat. Med. 2006, 12, 835–840. [Google Scholar] [CrossRef]
- Cheng, W.; Curti, E.; Rezende, W.C.; Kwityn, C.; Zhan, B.; Gillespie, P.; Plieskatt, J.; Joshi, S.B.; Volkin, D.B.; Hotez, P.J.; et al. Biophysical and formulation studies of the Schistosoma mansoni TSP-2 extracellular domain recombinant protein, a lead vaccine candidate antigen for intestinal schistosomiasis. Hum. Vaccines Immunother. 2013, 9, 2351–2361. [Google Scholar] [CrossRef] [Green Version]
- Curti, E.; Kwityn, C.; Zhan, B.; Gillespie, P.; Brelsford, J.; Deumic, V.; Plieskatt, J.; Rezende, W.C.; Tsao, E.; Kalampanayil, B.; et al. Expression at a 20L scale and purification of the extracellular domain of the Schistosoma mansoni TSP-2 recombinant protein: A vaccine candidate for human intestinal schistosomiasis. Hum. Vaccines Immunother. 2013, 9, 2342–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Hoeweler, L.; Keegan, B.; Peng, J.; Scholte, L.; Hotez, P.; Bottazzi, M.E.; Diemert, D.; Bethony, J. Potency testing for a recombinant protein vaccine early in clinical development: Lessons from the Schistosoma mansoni Tetraspanin 2 vaccine. Vaccine X 2021, 8, 100100. [Google Scholar] [CrossRef] [PubMed]
- Keitel, W.; Potter, G.; Diemert, D.; Bethony, J.; El Sahly, H.; Kennedy, J.; Patel, S.; Plieskatt, J.; Jones, W.; Deye, G.; et al. A phase 1 study of the safety, reactogenicity, and immunogenicity of a Schistosoma mansoni vaccine with or without glucopyranosyl lipid A aqueous formulation (GLA-AF) in healthy adults from a non-endemic area. Vaccine 2019, 37, 6500–6509. [Google Scholar] [CrossRef] [PubMed]
- Tendler, M.; Almeida, M.; Simpson, A. Development of the Brazilian Anti Schistosomiasis Vaccine Based on the Recombinant Fatty Acid Binding Protein Sm14 Plus GLA-SE Adjuvant. Front. Immunol. 2015, 6, 218. [Google Scholar] [CrossRef] [Green Version]
- Tendler, M.; Almeida, M.S.; Vilar, M.M.; Pinto, P.M.; Limaverde-Sousa, G. Current Status of the Sm14/GLA-SE Schistosomiasis Vaccine: Overcoming Barriers and Paradigms towards the First Anti-Parasitic Human(itarian) Vaccine. Trop. Med. Infect. Dis. 2018, 3, 121, Erratum in Trop. Med. Infect. Dis. 2019, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santini-Oliveira, M.; Pinto, P.M.; dos Santos, T.; Vilar, M.M.; Grinsztejn, B.; Veloso, V.; Paes-De-Almeida, E.C.; Amaral, M.A.Z.; Ramos, C.R.; Marroquin-Quelopana, M.; et al. Development of the Sm14/GLA-SE Schistosomiasis Vaccine Candidate: An Open, Non-Placebo-Controlled, Standardized-Dose Immunization Phase Ib Clinical Trial Targeting Healthy Young Women. Vaccines 2022, 10, 1724. [Google Scholar] [CrossRef]
- Molehin, A.J.; Sennoune, S.R.; Zhang, W.; Rojo, J.U.; Siddiqui, A.J.; Herrera, K.A.; Johnson, L.; Sudduth, J.; May, J.; Siddiqui, A.A. Cross-species prophylactic efficacy of Sm-p80-based vaccine and intracellular localization of Sm-p80/Sm-p80 ortholog proteins during development in Schistosoma mansoni, Schistosoma japonicum, and Schistosoma haematobium. Parasitol. Res. 2017, 116, 3175–3188. [Google Scholar] [CrossRef]
- Le, L.; Molehin, A.J.; Nash, S.; Sennoune, S.R.; Ahmad, G.; Torben, W.; Zhang, W.; Siddiqui, A.A. Schistosoma egg-induced liver pathology resolution by Sm-p80-based schistosomiasis vaccine in baboons. Pathology 2018, 50, 442–449. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Molehin, A.J.; Zhang, W.; Ganapathy, P.K.; Kim, E.; Rojo, J.U.; Redman, W.K.; Sennoune, S.R.; Sudduth, J.; Freeborn, J.; et al. Sm-p80-based vaccine trial in baboons: Efficacy when mimicking natural conditions of chronic disease, praziquantel therapy, immunization, and Schistosoma mansoni re-encounter. Ann. N. Y. Acad. Sci. 2018, 1425, 19–37. [Google Scholar] [CrossRef]
- Zhang, W.; Ahmad, G.; Molehin, A.J.; Torben, W.; Le, L.; Kim, E.; Lazarus, S.; Siddiqui, A.J.; Carter, D.; Siddiqui, A.A. Schistosoma mansoni Antigen Sm-p80: Prophylactic Efficacy using TLR4 Agonist Vaccine Adjuvant Glucopyranosyl Lipid A-Alum in Murine and Non-Human Primate Models. J. Investig. Med. 2018, 66, 1124–1132. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Molehin, A.J.; Rojo, J.U.; Sudduth, J.; Ganapathy, P.K.; Kim, E.; Siddiqui, A.J.; Freeborn, J.; Sennoune, S.R.; May, J.; et al. Sm-p80-based schistosomiasis vaccine: Double-blind preclinical trial in baboons demonstrates comprehensive prophylactic and parasite transmission-blocking efficacy. Ann. N. Y. Acad. Sci. 2018, 1425, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Le, L.; Ahmad, G.; Molehin, A.J.; Siddiqui, A.J.; Torben, W.; Karmakar, S.; Rojo, J.U.; Sennoune, S.; Lazarus, S.; et al. Fifteen Years of Sm-p80-Based Vaccine Trials in Nonhuman Primates: Antibodies from Vaccinated Baboons Confer Protection in vivo and in vitro From Schistosoma mansoni and Identification of Putative Correlative Markers of Protection. Front. Immunol. 2020, 11, 1246. [Google Scholar] [CrossRef]
- Hotez, P.J.; Matshaba, M. Promise of new malaria vaccines. BMJ 2022, 379, o2462. [Google Scholar] [CrossRef] [PubMed]
- Lund, A.J.; Wade, K.J.; Nikolakis, Z.L.; Ivey, K.N.; Perry, B.W.; Pike, H.N.; Paull, S.H.; Liu, Y.; Castoe, T.A.; Pollock, D.D.; et al. Integrating genomic and epidemiologic data to accelerate progress toward schistosomiasis elimination. Elife 2022, 11, e79320. [Google Scholar] [CrossRef]
- Oladipo, E.K.; Jimah, E.M.; Irewolede, B.A.; Folakanmi, E.O.; Olubodun, O.A.; Adediran, D.A.; Akintibubo, S.A.; Odunlami, F.D.; Olufemi, S.E.; Ojo, T.O.; et al. Immunoinformatics design of multi-epitope peptide for the diagnosis of Schistosoma haematobium infection. J. Biomol. Struct. Dyn. 2022, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, G.; Attenborough, T.; Rawlinson, K.A.; Berriman, M. Schistosomes go single cell. Nat. Rev. Microbiol. 2022, 20, 705. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J. The poverty-related neglected diseases: Why basic research matters. PLoS Biol. 2017, 15, e2004186. [Google Scholar] [CrossRef] [PubMed]
- Stothard, J.R.; Kayuni, S.A.; Al-Harbi, M.H.; Musaya, J.; Webster, B.L. Future schistosome hybridizations: Will all Schistosoma haematobium hybrids please stand-up! PLoS Negl. Trop. Dis. 2020, 14, e0008201. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Jones, M.K.; Gordon, C.A.; Arganda, A.E.; Cai, P.; Al-Wassiti, H.; Pouton, C.W.; McManus, D.P. The mRNA Vaccine Technology Era and the Future Control of Parasitic Infections. Clin. Microbiol. Rev. 2023, e0024121, Online ahead of print. [Google Scholar] [CrossRef]
- McManus, D.P.; Bergquist, R.; Cai, P.; Ranasinghe, S.; Tebeje, B.M.; You, H. Schistosomiasis—From immunopathology to vaccines. Semin. Immunopathol. 2020, 42, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Versteeg, L.; Almutairi, M.M.; Hotez, P.J.; Pollet, J. Enlisting the mRNA Vaccine Platform to Combat Parasitic Infections. Vaccines 2019, 7, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottazzi, M.E.; Hotez, P.J. “Running the Gauntlet”: Formidable challenges in advancing neglected tropical diseases vaccines from development through licensure, and a “Call to Action”. Hum. Vaccines Immunother. 2019, 15, 2235–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collyer, B.S.; Turner, H.C.; Hollingsworth, T.D.; Keeling, M.J. Vaccination or mass drug administration against schistosomiasis: A hypothetical cost-effectiveness modelling comparison. Parasites Vectors 2019, 12, 499. [Google Scholar] [CrossRef] [PubMed]
- Engels, D.; Hotez, P.J.; Ducker, C.; Gyapong, M.; Bustinduy, A.L.; Secor, W.E.; Harrison, W.; Theobald, S.; Thomson, R.; Gamba, V.; et al. Integration of prevention and control measures for female genital schistosomiasis, HIV and cervical cancer. Bull. World Health Organ. 2020, 98, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Mbah, M.L.N.; Skrip, L.; Greenhalgh, S.; Hotez, P.; Galvani, A.P. Impact of Schistosoma mansoni on Malaria Transmission in Sub-Saharan Africa. PLoS Negl. Trop. Dis. 2014, 8, e3234. [Google Scholar] [CrossRef] [Green Version]
- Kinyoki, D.; Osgood-Zimmerman, A.E.; Bhattacharjee, N.V.; Local Burden of Disease Anaemia Collaborators; Kassebaum, N.J.; Hay, S.I. Anemia prevalence in women of reproductive age in low- and middle-income countries between 2000 and 2018. Nat. Med. 2021, 27, 1761–1782. [Google Scholar] [CrossRef]
- Hotez, P.; Whitham, M. Helminth infections: A new global women’s health agenda. Obstet. Gynecol. 2014, 123, 155–160. [Google Scholar] [CrossRef]
- Hotez, P.J. Will anti-vaccine activism in the USA reverse global goals? Nat. Rev. Immunol. 2022, 22, 525–526. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J. Blue Marble Health: An Innovative Plan to Fight Diseases of the Poor amid Wealth; Johns Hopkins University Press: Baltimore, MD, USA, 2016. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotez, P.J.; Bottazzi, M.E. Human Schistosomiasis Vaccines as Next Generation Control Tools. Trop. Med. Infect. Dis. 2023, 8, 170. https://doi.org/10.3390/tropicalmed8030170
Hotez PJ, Bottazzi ME. Human Schistosomiasis Vaccines as Next Generation Control Tools. Tropical Medicine and Infectious Disease. 2023; 8(3):170. https://doi.org/10.3390/tropicalmed8030170
Chicago/Turabian StyleHotez, Peter J., and Maria Elena Bottazzi. 2023. "Human Schistosomiasis Vaccines as Next Generation Control Tools" Tropical Medicine and Infectious Disease 8, no. 3: 170. https://doi.org/10.3390/tropicalmed8030170