
Socioconnectomics: Connectomics Should Be Extended to Societies to Better Understand Evolutionary Processes


1
CNRS, IPHC UMR 7178, Université de Strasbourg, 67000 Strasbourg, France
2
Institut Universitaire de France, 75005 Paris, France
3
Anthropolab, ETHICS EA 7446, Université Catholique de Lille, 59000 Lille, France
Sci 2023, 5(1), 5; https://doi.org/10.3390/sci5010005
Submission received: 7 November 2022
/
Revised: 13 January 2023
/
Accepted: 29 January 2023
/
Published: 30 January 2023
(This article belongs to the Section Biology Research and Life Sciences)
{kind=link}
Abstract
:Connectomics, which is the network study of connectomes or maps of the nervous system of an organism, should be applied and expanded to human and animal societies, resulting in the birth of the domain of socioconnectomics compared to neuroconnectomics. This new network study framework would open up new perspectives in evolutionary biology and add new elements to theories, such as the social and cultural brain hypotheses. Answering questions about network topology, specialization, and their connections with functionality at one level (i.e., neural or societal) may help in understanding the evolutionary trajectories of these patterns at the other level. Expanding connectomics to societies should be done in comparison and combination with multilevel network studies and the possibility of multiorganization selection processes. The study of neuroconnectomes and socioconnectomes in animals, from simpler to more advanced ones, could lead to a better understanding of social network evolution and the feedback between social complexity and brain complexity.
1. Introduction
Connectomics is the study of connectomes [1,2,3,4] or maps of the nervous system of an organism, mainly the brain in vertebrates and its analogous structure in invertebrates. Connectomics enables researchers to understand how information is treated and stored on microscopic (neuron basis) and macroscopic (brain module basis) scales. For example, how a disease affects brain structure and function or the variability in brain structure and how it changes with age. Connectomics is a set of network tools that helps researchers understand how the connectional architecture of the brain is related to functional specialization. Recent computational advances have improved the measurement and storage of large datasets—those with thousands of connections between thousands of entities—analyzed primarily using graph theory or social network analyses (SNAs) [5,6]. These SNAs are used to assess how neural networks are structured regarding communities (i.e., modules), centralities (i.e., hubs and rich clubs), and efficiency (i.e., short communication paths), resulting in comparative and evolutionary applications concerning the function and selection of these neuronal architectures in different species [4]. Compared to neuroconnectomics, I propose that connectomics be applied and expanded to human and animal societies and social networks, leading to the birth of the domain of socioconnectomics. This involves transferring network analysis tools, applying natural selection concepts to social networks, and considering evolutionary processes (genetic selection, cultural selection, and multilevel selection [7,8]). This new study framework would open up new perspectives in evolutionary biology and add new components to theories such as the social brain [9] and cultural brain hypotheses [10]. According to the social brain hypothesis, brain size, particularly the neocortex, is related to group size. In contrast, the cultural brain hypothesis unifies more specific explanations centered on the environmental and social brain hypotheses, implying that the brain was chosen for its ability to store and manage information acquired through asocial or social learning [10,11]. Therefore, the cultural brain hypothesis is a bit more complex and includes more parameters than the social brain hypothesis. Managing the complexity of social relationships—not just group size but identifying them, memorizing them, and identifying the social and cultural roles of group members according to environmental conditions—is challenging for animals, including humans. It should result in a link between social network complexity and brain complexity, indicating a link between neuroconnectomics and socioconnectomics.
2. Universal Network Topological Properties
Based on the findings of Milo et al. [12], Oltvai and Barabási, in their study [13], proposed that the universal topological properties (e.g., scale-free connectivity, modularity, and hierarchy) observed from cellular to neural and ecosystem networks should be governed by similar principles. Other studies later proposed that such common features should reflect constraints imposed by selection forces and be shaped by evolutionary pressures [14,15]. Irrespective of the selection mechanism, we may wonder why networks—brain or society—“look the way they do and not any other way” [14]. Questions about neural networks are raised for human and animal societies, regarding how genetic and environmental interactions influence topology and its variability, how diseases affect structures, and how information is identified and shared [15]. Connectome architecture may follow general wiring principles that favor functionally important topological attributes that benefit efficient communication and global integration while minimizing biological resource expenditure. Many animals, including humans, live in societies because socialization improves fitness [16], allowing them to learn more about food, reproduction, and predators. However, proximity and social relationships increase the risk of disease. Individuals may become specialized to increase group efficiency, such as elders sharing information and leading, as societies face trade-offs of opposing forces increasing modularity [17,18]. Hence, answering questions about topology, specialization, and their links to functionality at one level (i.e., neural or societal) may help in understanding the evolutionary trajectories of these patterns at the other level. Henrich revealed that human evolutionary processes at the brain and social levels are always constant due to constant exchanges between these two structures [19]. Specific behavioral traits or personalities may be favored by genetic selection to a specific phenotypic composition of a group [20] and network topology [20,21]. For example, aggressive traits may be connected to modular networks. Similarly, cultural selection influences the transmission of adaptive behavior, and group members may choose to observe the best performers, modifying social network centralization [22,23]. Similar network “morphospaces” should be observed at these different levels if general selection principles apply to different levels of life organization [14]. Selection of network properties such as efficiency or modularity has been observed at the brain [24,25] and the group levels [26,27,28], implying that similar evolutionary processes may be responsible for these network topologies. For example, the network modularity of primate groups is higher than the modularity of theoretical networks with similar properties (e.g., size and density) [27], indicating a selection process supported by the fact that this network property and efficiency are linked to brain capacities in primates [26]. The same reasoning could be applied to ecosystems and their tropic and mutualistic networks [29].
3. Network Analyses at Different Life Organizations
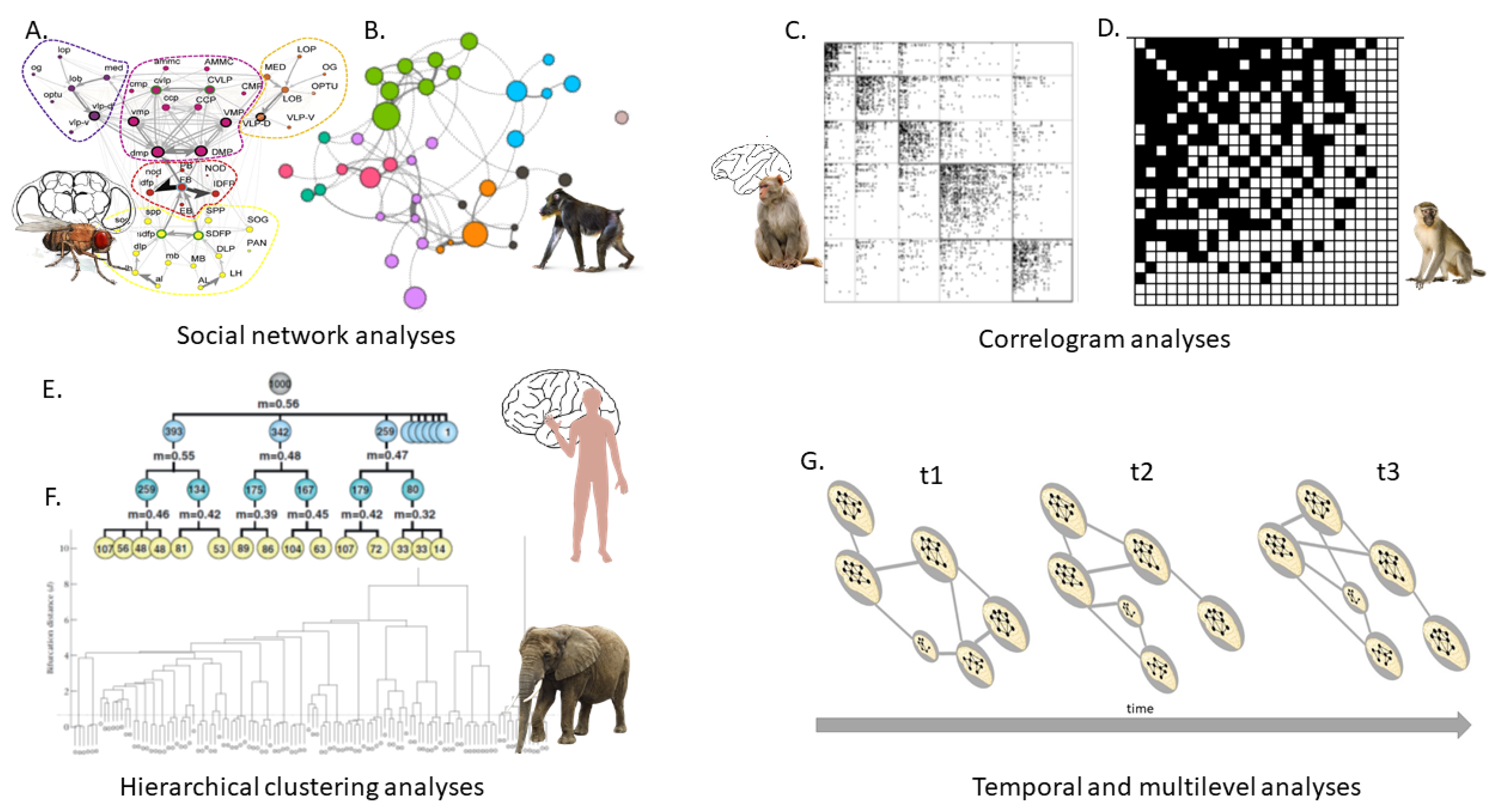
Neural and social networks share a theoretical foundation, and they also share network analysis tools. This is not new, as social network analyses have been used in both domains for approximately 20 years. However, this statement contributes to a more solid understanding of socioconnectomics as neuroconnectomics. Measuring the connection or activity of an entity—whether a neuron or an individual—differs from neuroconnectomics, which is based on magnetic resonance imaging, electron microscopy, and histological techniques [1]. In contrast, socioconnectomics is based on ethological observations or loggers [30]. However, the same network analyses are used to work on connectomes when these connections are measured (Figure 1). The generalized mathematical language of graph theory is now used in many scientific disciplines, allowing possible comparisons of connectomes at different levels. SNAs are a set of measures used at different levels of the network [6]: the entity level (a neuron or an individual), the intermediate level with the presence of clusters and their relationships, and the network level to assess global properties such as efficiency, diameter, density, or modularity [6,31]. When it comes to understanding the role of certain neurons or brain areas in linking others and explaining brain asymmetry, efficiency, or robustness (i.e., the ability to withstand failures and perturbations, mainly alternative pathways between two entities), betweenness centrality is also a popular and useful measure in animal social networks to understand the roles of individuals. For example, individuals with a high betweenness link different communities, allowing the transfer of information and leadership in dolphins [32] and dominance and policing characteristics in macaques (Figure 1A,B) [33]. Correlograms, which are used to show the existence of modules in the brain, are also used to determine communities in primates and how these communities are nested based on dominance (Figure 1C,D) [34,35]. Dendrograms and hierarchical clustering analyses can also be used to assess communities (Figure 1E,F). This has been demonstrated in elephants, where units fuse and split depending on the dryness of the season [36].
4. Same Challenges to Understanding Evolution
The fact that the same toolkit can be used for neuroconnectomics and socioconnectomics is significant because this implies that the results can be compared between these two levels, allowing for comparative connectomics not only between brains [4] or societies [38] of different species but also between brains and societies. However, trials comparing different levels of life organization in network topologies are extremely rare [12,13,35,39,40] because of a lack of exchange between scientific communities and different ways of considering evolutionary processes driving network topology. The same challenges arise when combining connectomics with comparative ethology and comparative neuroscience to discover common organizational plans while accounting for species diversity. However, scientists in both domains are concerned with the same methodological issues in obtaining, storing, and analyzing large datasets [2], necessitating the development of new tools. Both neuroscientists and behavioral ecologists would benefit from sharing technologies in this context. Beyond this technological comparison, the field of “comparative network analysis” [4] is used to examine similarities and differences across a range of network classes. It provides a powerful approach to answering the question of which topological network attributes are specific to nervous structures and which represent more universal network organization properties as human and animal societies. Therefore, it offers a unique tool for determining which network properties are unique to brain networks and which are shared with social networks.
5. Future Research and Challenges
Expanding connectomics to societies should be done in comparison and combination with multilevel studies and the possibility of multiorganization selection processes [7]. Evolutionary processes may favor the phenotypic composition of a group [41], implying that individuals with different personalities and, thus, social traits in a group may favor different network topologies. This is known as “collective social niche construction” [7]. This type of multilevel connectomics could be envisioned in two ways: first, as a multispecies comparison, and second, as a lifespan comparison, allowing researchers to assess the selection processes (genetic versus cultural) leading to specific network topologies.
Multilevel selection: There is no argument that the brain network is subject to selection pressures and evolves to reach a certain level of optimality [24,25,42,43]. However, it is more difficult to propose the same evolutionary theory for social network topology [44,45,46,47,48,49]. Hence, it is challenging to apply connectomics to societies in the same way it is applied to the brain. However, the social network topology is governed by the group behaviors of group members [8,50], whereas individual behavior is governed by genetic [51,52,53] and cultural selection [54,55] ([7] for a review of mechanisms). Pressures such as predation, access to information, pathogens, access to food, and mating may favor certain behaviors of individuals to increase their fitness by making them more central in their networks, closer to relatives, and so on. Therefore, it is possible to observe the selection of some network properties that may eventually favor the fitness of both some and all group members, even if the benefit is not distributed equally. Hence, rather than considering selection on a group level, I consider it on multiple levels [8,45]. Multilevel selection typically favors phenotypic benefits that can be shared by all group members [56]. The coding of a group phenotypic composition [41] for network topology may be favored by evolutionary processes and could be referred to as “collective social niche construction” [7]. This consideration makes it possible to apply connectomics to societies.
Multilevel connectomics: The study of neuroconnectomes and socioconnectomes in animals, from simpler to more advanced ones, could lead to a better understanding of social evolution and the feedback between social and brain complexity. It would be specific about how social structures influence individual fitness and, consequently, the selection of heritable social traits. The cultural brain hypothesis states that brain complexity in humans is not only due to our social complexity but also to the ability of our brain to store, manage, and share information [10,11]. Directly comparing neuroconnectomes with socioconnectomes of different individuals in different species, as with the cultural brain hypothesis, by comparing different ethnic groups could add more elements to confirm or deny the social brain hypothesis. A similarity index could be measured between indices (e.g., efficiency, modularity) of the neuroconnectome and the socioconnectome of an individual, human, or animal, and the similarity index could be compared across species, environmental conditions, and other variables. One important question is how the brain and social complexity are linked. Several studies have found a positive correlation between the two connectomes in mammals [9,57], giving rise to the social brain hypothesis and, later, the cultural brain hypothesis. However, these hypotheses are still being debated [58,59,60,61]. The link is not obvious in other animal taxa and may even be inversed, as in eusocial insects, whose brain structures may be simpler than in vertebrates. Nevertheless, societies remain highly complex [42,62]. Moreover, it would be critical to show how certain neural connectomes underpin specific social behaviors. We already know that certain brain parts are linked with some behaviors, such as language, emotions, and other capacities. Apart from the connection between different parts of the brain allowing the emergence of consciousness, which is linked to the theory of mind and social complexity, we do not know how neuroconnectomes may influence socioconnectomes. Beyond simple correlation studies, one could consider selecting specific neuroconnectomes in species such as drosophila and assess how they are linked to the social behavior of the species and the emergent social network.
Aging connectomics: Several studies have found a link between social complexity and brain size based on the social brain hypothesis [9]. However, developing social interaction capability in vertebrates and their corresponding neural networks involves a critical temporal dimension (Figure 1G) [63]. Nonetheless, this longitudinal aspect is just gaining momentum. Animal brain size, structures, and social relationships take time to develop, and may take several years [64,65,66]. This implies that there is a temporal dimension to investigate in developing these two structures. This hypothesis has recently been tested intraspecifically with some individual brain structures linked to several social partners [67]. However, very few studies have been conducted to investigate how some brain structures are linked to social centralities of group members and, more generally, network topologies, and how age affects neuroconnectomes and socioconnectomes. Measuring the two parameters at different ages or life stages of individuals would reveal processes that link sociality to longevity and brain aging. Multilevel connectomics, which means linking the neuroconnectome to the socioconnectome, could also be envisaged across the lifespan [3] of an individual, from birth to death, as both sociality and the brain change with age. The social network of an individual specifically is flexible throughout their life and appears to be partly independent of chronological age, with regulatory processes and bidirectional effects on individual senescence and fitness [63]. The question of the directionality of the effect between changes in the brain and social network remains unanswered, and a detailed longitudinal study of connectomes spanning the lives of individuals could provide an answer to the causality issue. It could also lead to direct applications in brain diseases linked to sociality. It is difficult to study this developmental covariation between neuroconnectomes and socioconnectomes because it requires measuring. For example, the brain structure of all group members undergoes changes throughout their lives. Technologies such as fMRIs exist only for animals of a certain size and are still expensive for large and long-term studies.
6. Conclusions
To better understand how our brain and social world are structured, we can use network analysis tools to connect two disparate domains while answering similar questions about network properties and functionalities. In the same way we study brain structure (neuroconnectomics), we should study the evolution of social structures (connectomics). This way of thinking, which incorporates multilevel selection, could lead to a better understanding of the back and forth between different levels of organizations. Further, it advances our understanding of brain complexity concerning sociality, adding elements to the social and cultural brain hypotheses. Hence, as neuroconnectomics is the study of brain structures and the evolutionary processes underlying them, socioconnectomics should become the study of social networks and the evolutionary processes underlying them. This framework could also be extended to other types of networks, potentially resolving one of the most important scientific questions concerning the universality and evolution of some physical and biological rules. Further, supposed selection and evolution were found to apply to many levels of living organizations [12,14]. In that case, there is no reason why they should not also apply to all of them, particularly to animal and human social networks [7,68,69].
Funding
This research is funded by the French Academic Institute (IUF).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Morgan, J.L.; Lichtman, J.W. Why Not Connectomics? Nat. Methods 2013, 10, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, J.W.; Pfister, H.; Shavit, N. The Big Data Challenges of Connectomics. Nat. Neurosci. 2014, 17, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.-N.; He, Y.; Betzel, R.F.; Colcombe, S.; Sporns, O.; Milham, M.P. Human Connectomics across the Life Span. Trends Cogn. Sci. 2017, 21, 32–45. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Bullmore, E.T.; Sporns, O. Comparative Connectomics. Trends Cogn. Sci. 2016, 20, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgatti, S.P.; Mehra, A.; Brass, D.J.; Labianca, G. Network Analysis in the Social Sciences. Science 2009, 323, 892–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa, S.; Sueur, C.; Puga-Gonzalez, I. Network Measures in Animal Social Network Analysis: Their Strengths, Limits, Interpretations and Uses. Methods Ecol. Evol. 2021, 12, 10–21. [Google Scholar] [CrossRef]
- Sueur, C.; Romano, V.; Sosa, S.; Puga-Gonzalez, I. Mechanisms of Network Evolution: A Focus on Socioecological Factors, Intermediary Mechanisms, and Selection Pressures. Primates 2019, 60, 167–181. [Google Scholar] [CrossRef]
- Fisher, D.; McAdam, A. Social Traits, Social Networks and Evolutionary Biology. J. Evol. Biol. 2017, 30, 2088–2103. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, R. The Social Brain Hypothesis. Evol. Anthropol. Issues News Rev. 1998, 6, 178–190. [Google Scholar] [CrossRef]
- Muthukrishna, M.; Doebeli, M.; Chudek, M.; Henrich, J. The Cultural Brain Hypothesis: How Culture Drives Brain Expansion, Sociality, and Life History. PLoS Comput. Biol. 2018, 14, e1006504. [Google Scholar] [CrossRef]
- van Schaik, C.P.; Isler, K.; Burkart, J.M. Explaining Brain Size Variation: From Social to Cultural Brain. Trends Cogn. Sci. 2012, 16, 277–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltvai, Z.N.; Barabási, A.-L. Life’s Complexity Pyramid. Science 2002, 298, 763–764. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Shen-Orr, S.; Itzkovitz, S.; Kashtan, N.; Chklovskii, D.; Alon, U. Network Motifs: Simple Building Blocks of Complex Networks. Science 2002, 298, 824–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avena-Koenigsberger, A.; Goñi, J.; Solé, R.; Sporns, O. Network Morphospace. J. R. Soc. Interface 2015, 12, 20140881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, V.; MacIntosh, A.J.J.; Sueur, C. Stemming the Flow: Information, Infection, and Social Evolution. Trends Ecol. Evol. 2020, 35, 849–853. [Google Scholar] [CrossRef]
- Silk, J.B.; Alberts, S.C.; Altmann, J. Social Bonds of Female Baboons Enhance Infant Survival. Science 2003, 302, 1231–1234. [Google Scholar] [CrossRef] [Green Version]
- McComb, K.; Moss, C.; Durant, S.M.; Baker, L.; Sayialel, S. Matriarchs As Repositories of Social Knowledge in African Elephants. Science 2001, 292, 491–494. [Google Scholar] [CrossRef] [Green Version]
- Nagy, M.; Akos, Z.; Biro, D.; Vicsek, T. Hierarchical Group Dynamics in Pigeon Flocks. Nature 2010, 464, 890–893. [Google Scholar] [CrossRef] [Green Version]
- Henrich, J. The Secret of Our Success: How Culture Is Driving Human Evolution, Domesticating Our Species, and Making Us Smarter; Princeton University Press: Princeton, NJ, USA, 2017; ISBN 0-691-17843-7. [Google Scholar]
- Farine, D.R.; Montiglio, P.O.; Spiegel, O. From Individuals to Groups and Back: The Evolutionary Implications of Group Phenotypic Composition. Trends Ecol. Evol. 2015, 30, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Bijma, P. A General Definition of the Heritable Variation That Determines the Potential of a Population to Respond to Selection. Genetics 2011, 189, 1347–1359. [Google Scholar] [CrossRef]
- Bijma, P.; Wade, M. The Joint Effects of Kin, Multilevel Selection and Indirect Genetic Effects on Response to Genetic Selection. J. Evol. Biol. 2008, 21, 1175–1188. [Google Scholar] [CrossRef] [PubMed]
- Grampp, M.; Sueur, C.; van de Waal, E.; Botting, J. Social Attention Biases in Juvenile Wild Vervet Monkeys: Implications for Socialisation and Social Learning Processes. Primates 2019, 60, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Canteloup, C.; Hoppitt, W.; van de Waal, E. Wild Primates Copy Higher-Ranked Individuals in a Social Transmission Experiment. Nat. Commun. 2020, 11, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clune, J.; Mouret, J.B.; Lipson, H. The Evolutionary Origins of Modularity. Proc. Biol. Sci. 2013, 280, 20122863. [Google Scholar] [CrossRef] [Green Version]
- Bullmore, E.; Sporns, O. The Economy of Brain Network Organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef]
- Pasquaretta, C.; Levé, M.; Claidière, N.; van de Waal, E.; Whiten, A.; MacIntosh, A.J.J.; Pelé, M.; Bergstrom, M.L.; Borgeaud, C.; Brosnan, S.F.; et al. Social Networks in Primates: Smart and Tolerant Species Have More Efficient Networks. Sci. Rep. 2014, 4, 7600. [Google Scholar] [CrossRef] [Green Version]
- Romano, V.; Shen, M.; Pansanel, J.; MacIntosh, A.J.J.; Sueur, C. Social Transmission in Networks: Global Efficiency Peaks with Intermediate Levels of Modularity. Behav. Ecol. Sociobiol. 2018, 72, 154. [Google Scholar] [CrossRef]
- Romano, V.; Sueur, C.; MacIntosh, A.J.J. The Trade-off between Information and Pathogen Transmission in Animal Societies. Oikos 2022, 2022, e08290. [Google Scholar] [CrossRef]
- Thébault, E.; Fontaine, C. Stability of Ecological Communities and the Architecture of Mutualistic and Trophic Networks. Science 2010, 329, 853–856. [Google Scholar] [CrossRef]
- Strandburg-Peshkin, A.; Farine, D.R.; Couzin, I.D.; Crofoot, M.C. Shared Decision-Making Drives Collective Movement in Wild Baboons. Science 2015, 348, 1358–1361. [Google Scholar] [CrossRef]
- Sueur, C.; Jacobs, A.; Amblard, F.; Petit, O.; King, A.J. How Can Social Network Analysis Improve the Study of Primate Behavior? Am. J. Primatol. 2011, 73, 703–719. [Google Scholar] [CrossRef]
- Lusseau, D.; Conradt, L. The Emergence of Unshared Consensus Decisions in Bottlenose Dolphins. Behav. Ecol. Sociobiol. 2009, 63, 1067–1077. [Google Scholar] [CrossRef] [Green Version]
- Flack, J.C.; Girvan, M.; de Waal, F.B.; Krakauer, D.C. Policing Stabilizes Construction of Social Niches in Primates. Nature 2006, 439, 426–429. [Google Scholar] [CrossRef]
- Fruteau, C.; Voelkl, B.; van Damme, E.; Noë, R. Supply and Demand Determine the Market Value of Food Providers in Wild Vervet Monkeys. Proc. Natl. Acad. Sci. USA 2009, 106, 12007–12012. [Google Scholar] [CrossRef] [Green Version]
- Cantor, M.; Pires, M.M.; Marquitti, F.M.; Raimundo, R.L.; Sebastián-González, E.; Coltri, P.P.; Perez, S.I.; Barneche, D.R.; Brandt, D.Y.; Nunes, K.; et al. Nestedness across Biological Scales. PLoS ONE 2017, 12, e0171691. [Google Scholar] [CrossRef] [Green Version]
- Wittemyer, G.; Douglas-Hamilton, I.; Getz, W.M. The Socioecology of Elephants: Analysis of the Processes Creating Multitiered Social Structures. Anim. Behav. 2005, 69, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Honey, C.J.; Sporns, O.; Cammoun, L.; Gigandet, X.; Thiran, J.P.; Meuli, R.; Hagmann, P. Predicting Human Resting-State Functional Connectivity from Structural Connectivity. Proc. Natl. Acad. Sci. USA 2009, 106, 2035–2040. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, R.I.M. The Social Brain Meets Neuroimaging. Trends Cogn. Sci. 2012, 16, 101–102. [Google Scholar] [CrossRef]
- Sueur, C. Social Network, Information Flow and Decision-Making Efficiency: A Comparison of Humans and Animals. In Social Networking and Community Behavior Modeling; IGI Global: Hershey, PA, USA, 2011; pp. 164–177. [Google Scholar]
- Sueur, C.; King, A.J.; Pelé, M.; Petit, O. Fast and Accurate Decisions as a Result of Scale-Free Network Properties in Two Primate Species. In Proceedings of the Lecture Notes in Computer Science; Springer: Cham, Switzerland, 2012; pp. 579–584. [Google Scholar]
- Marshall, J.A.R.; Bogacz, R.; Dornhaus, A.; Planqué, R.; Kovacs, T.; Franks, N.R. On Optimal Decision-Making in Brains and Social Insect Colonies. J. R. Soc. Interface 2009, 6, 1065–1074. [Google Scholar] [CrossRef]
- Bogacz, R. Optimal Decision-Making Theories: Linking Neurobiology with Behaviour. Trends Cogn. Sci. 2007, 11, 118–125. [Google Scholar] [CrossRef]
- Nowak, M.A. Five Rules for the Evolution of Cooperation. Science 2006, 314, 1560–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traulsen, A.; Nowak, M.A. Evolution of Cooperation by Multilevel Selection. Proc. Nat. Acad. Sci. USA 2006, 103, 10952–10955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, M.A.; Tarnita, C.E.; Wilson, E.O. Nowak et al. Reply. Nature 2011, 471, E9–E10. [Google Scholar] [CrossRef] [Green Version]
- Leigh, E.G., Jr. The Group Selection Controversy. J. Evol. Biol. 2010, 23, 6–19. [Google Scholar] [CrossRef]
- Wilson, D.S. The Tide of Opinion on Group Selection Has Turned. In This View Life; Evolution Institute: Tampa, FL, USA, 2015; Volume 26. [Google Scholar]
- Wade, M.J. A Critical Review of the Models of Group Selection. Q. Rev. Biol. 1978, 53, 101–114. [Google Scholar] [CrossRef]
- Moscovice, L.R.; Sueur, C.; Aureli, F. How Socio-Ecological Factors Influence the Differentiation of Social Relationships: An Integrated Conceptual Framework. Biol. Lett. 2020, 16, 20200384. [Google Scholar] [CrossRef]
- Lesch, K.-P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Muller, C.R.; Hamer, D.H.; Murphy, D.L. Association of Anxiety-Related Traits with a Polymorphism in the Serotonin Transporter Gene Regulatory Region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Wilson, A.J.; Gelin, U.; Perron, M.-C.; Réale, D. Indirect Genetic Effects and the Evolution of Aggression in a Vertebrate System. Proc. Biol. Sci. 2009, 276, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Brent, L.J.N.; Heilbronner, S.R.; Horvath, J.E.; Gonzalez-Martinez, J.; Ruiz-Lambides, A.; Robinson, A.G.; Skene, J.H.P.; Platt, M.L. Genetic Origins of Social Networks in Rhesus Macaques. Sci. Rep. 2013, 3, srep01042. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, R. The Selfish Gene: 30th Anniversary Edition; Oxford University Press: Oxford, UK, 2006; ISBN 978-0-19-157406-1. [Google Scholar]
- Whiten, A.; Ayala, F.J.; Feldman, M.W.; Laland, K.N. The Extension of Biology through Culture. Proc. Natl. Acad. Sci. USA 2017, 114, 7775–7781. [Google Scholar] [CrossRef]
- Nonacs, P.; Kapheim, K.M. Social Heterosis and the Maintenance of Genetic Diversity. J. Evol. Biol. 2007, 20, 2253–2265. [Google Scholar] [CrossRef] [PubMed]
- Finarelli, J.A.; Flynn, J.J. Brain-Size Evolution and Sociality in Carnivora. Proc. Natl. Acad. Sci. USA 2009, 106, 9345–9349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeCasien, A.R.; Williams, S.A.; Higham, J.P. Primate Brain Size Is Predicted by Diet but Not Sociality. Nat. Ecol. Evol. 2017, 1, 112. [Google Scholar] [CrossRef] [PubMed]
- Shultz, S.; Dunbar, R.I. Both Social and Ecological Factors Predict Ungulate Brain Size. Proc. Biol. Sci. 2005, 273, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Burish, M.J.; Kueh, H.Y.; Wang, S.S. Brain Architecture and Social Complexity in Modern and Ancient Birds. Brain Behav. Evol. 2004, 63, 107–124. [Google Scholar] [CrossRef]
- Barrett, L.; Henzi, P.; Rendall, D. Social Brains, Simple Minds: Does Social Complexity Really Require Cognitive Complexity? Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Leadbeater, E.; Chittka, L. Social Learning in Insects—From Miniature Brains to Consensus Building. Curr. Biol. 2007, 17, R703–R713. [Google Scholar] [CrossRef] [Green Version]
- Sueur, C.; Quque, M.; Naud, A.; Bergouignan, A.; Criscuolo, F. Social capital: An independent dimension of healthy ageing. Peer Community J. 2021, 1, e23. [Google Scholar] [CrossRef]
- Almeling, L.; Hammerschmidt, K.; Sennhenn-Reulen, H.; Freund, A.M.; Fischer, J. Motivational Shifts in Aging Monkeys and the Origins of Social Selectivity. Curr. Biol. CB 2016, 26, 1744–1749. [Google Scholar] [CrossRef] [Green Version]
- Rosati, A.G.; Hagberg, L.; Enigk, D.K.; Otali, E.; Thompson, M.E.; Muller, M.N.; Wrangham, R.W.; Machanda, Z.P. Social Selectivity in Aging Wild Chimpanzees. Science 2020, 370, 473–476. [Google Scholar] [CrossRef]
- Albery, G.F.; Clutton-Brock, T.H.; Morris, A.; Morris, S.; Pemberton, J.M.; Nussey, D.H.; Firth, J.A. Ageing Red Deer Alter Their Spatial Behaviour and Become Less Social. Nat. Ecol. Evol. 2022, 6, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Testard, C.; Brent, L.J.N.; Andersson, J.; Chiou, K.L.; Negron-Del Valle, J.E.; DeCasien, A.R.; Acevedo-Ithier, A.; Stock, M.K.; Antón, S.C.; Gonzalez, O.; et al. Social Connections Predict Brain Structure in a Multidimensional Free-Ranging Primate Society. Sci. Adv. 2022, 8, eabl5794. [Google Scholar] [CrossRef] [PubMed]
- Solé, R.; Moses, M.; Forrest, S. Liquid Brains, Solid Brains. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190040. [Google Scholar] [CrossRef] [PubMed]
- Almaatouq, A.; Noriega-Campero, A.; Alotaibi, A.; Krafft, P.; Moussaid, M.; Pentland, A. Adaptive Social Networks Promote the Wisdom of Crowds. Proc. Natl. Acad. Sci. USA 2020, 117, 11379–113686. [Google Scholar] [CrossRef]
Figure 1.
Instances of analyses used in neural and social networks study. (A,B) Common social network analyses used to assess individual (e.g., neuron or group member) and global properties, respectively, in the Drosophila brain and a group of mandrills. (C,D) Correlogram analyses measuring the existence of modules and nestedness [35] in the macaque brain and a group of vervet monkeys. (E,F) Analyses measuring clusterization in the human brain [37] and an African elephant population [36]. (G) Framework to study and compare socioconnectomes with neuroconnectomes over time. Figures are used with permission.
Figure 1.
Instances of analyses used in neural and social networks study. (A,B) Common social network analyses used to assess individual (e.g., neuron or group member) and global properties, respectively, in the Drosophila brain and a group of mandrills. (C,D) Correlogram analyses measuring the existence of modules and nestedness [35] in the macaque brain and a group of vervet monkeys. (E,F) Analyses measuring clusterization in the human brain [37] and an African elephant population [36]. (G) Framework to study and compare socioconnectomes with neuroconnectomes over time. Figures are used with permission.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sueur, C. Socioconnectomics: Connectomics Should Be Extended to Societies to Better Understand Evolutionary Processes. Sci 2023, 5, 5. https://doi.org/10.3390/sci5010005
AMA Style
Sueur C. Socioconnectomics: Connectomics Should Be Extended to Societies to Better Understand Evolutionary Processes. Sci. 2023; 5(1):5. https://doi.org/10.3390/sci5010005
Chicago/Turabian StyleSueur, Cédric. 2023. "Socioconnectomics: Connectomics Should Be Extended to Societies to Better Understand Evolutionary Processes" Sci 5, no. 1: 5. https://doi.org/10.3390/sci5010005