Confocal and Electron Microscopic Structure of the Cornea from Three Species of Penguin

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
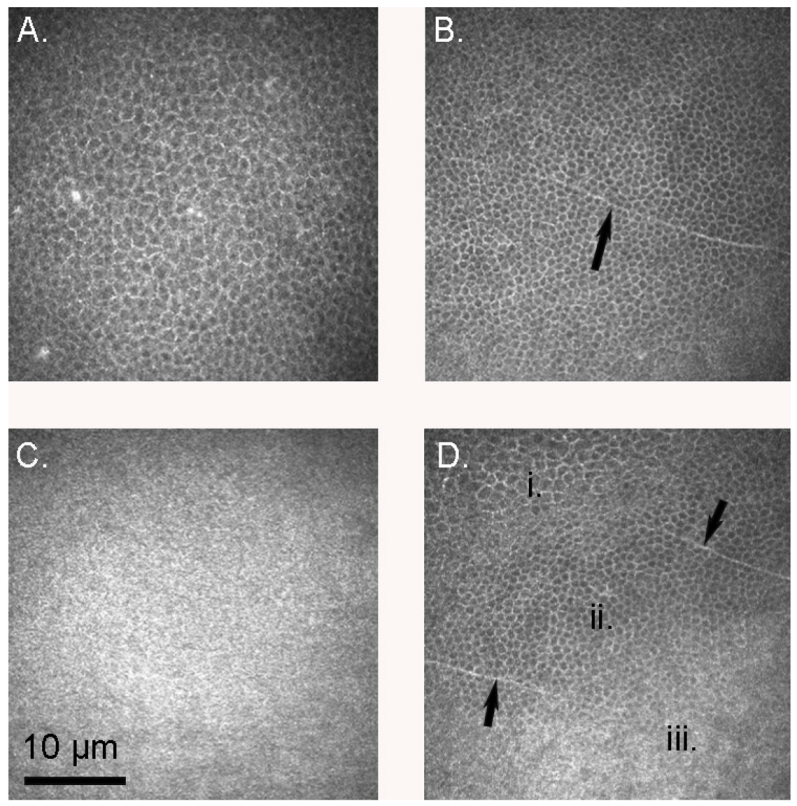
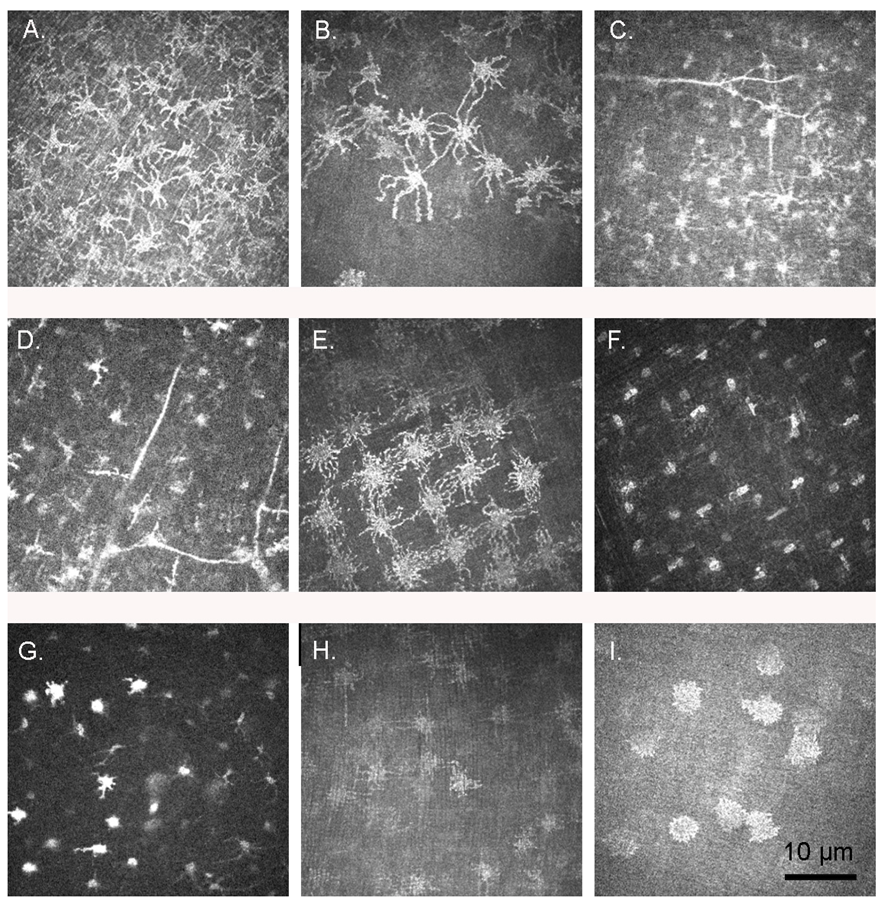
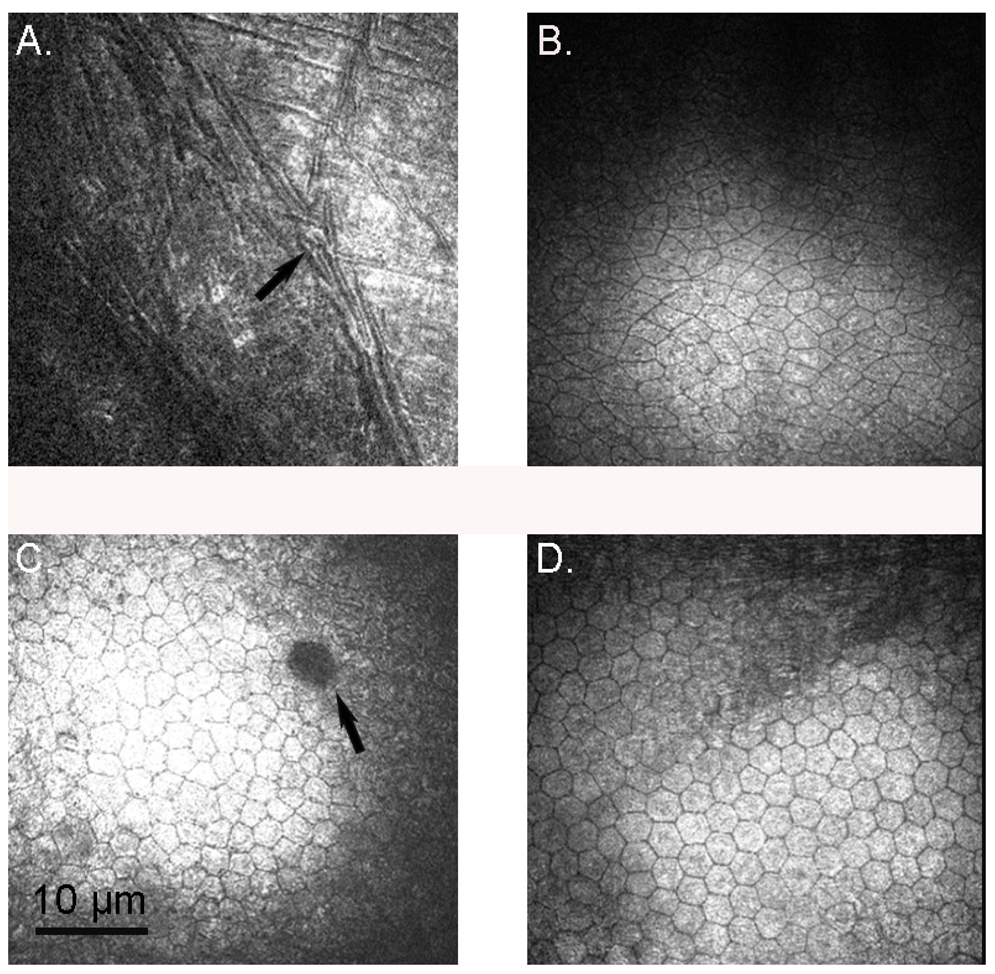
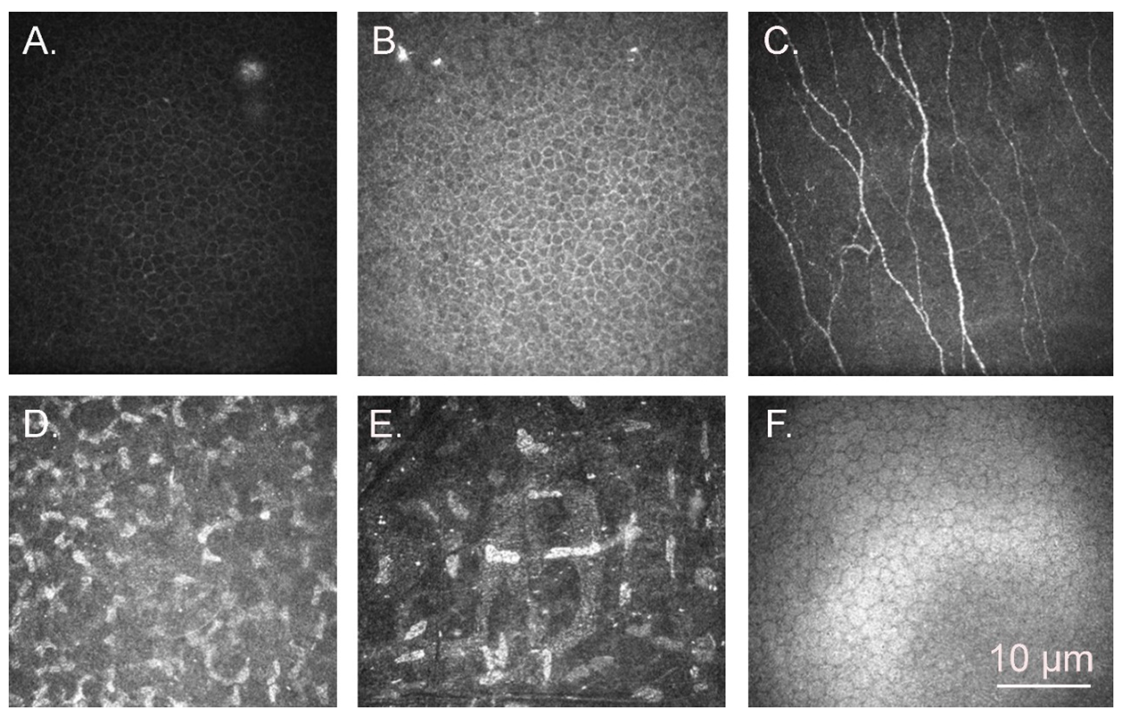
3. Results
3.1. Confocal Microscopy
3.2. Transmission Electron Microscopy (TEM)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Martin, G.R.; Young, S. The eye of the Humboldt penguin, Spheniscus humboldti: Visual fields and schematic optics. Proc. R. Soc. B 1984, 223, 197–222. [Google Scholar]
- Martin, G.R. Eye structure and foraging in king penguins Aptenodytes patagonicus. IBIS 1999, 141, 444–450. [Google Scholar] [CrossRef]
- Hadden, P.W.; Vorobyev, M.; Cassidy, S.B.; Gokul, A.; Simkin, S.K.; Tran, H.; McGhee, C.N.J.; Zhang, J. Selected ocular dimensions of three penguin species. Vis. Res. 2022, 201, 108122. [Google Scholar] [CrossRef]
- Collin, S.P.; Collin, H.B. Functional morphology of the cornea of the little penguin Eudyptula minor (Aves). J. Anat. 2021, 239, 732–746. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.C.; Mitchell, A.D.; Waas, J.R.; Paterson, A.M. An unexpected pattern of molecular divergence within the blue penguin (Eudyptula minor) complex. Notornis 2002, 49, 29–38. [Google Scholar]
- Grosser, S.; Scofield, R.P.; Waters, J.M. Multivariate skeletal analyses support a taxonomic distinction between New Zealand and Australian Eudyptula penguins (Sphenisciformes: Spheniscidae). Emu-Austral Ornithol. 2017, 117, 276–283. [Google Scholar] [CrossRef]
- Grosser, S. Coalescent modelling suggests recent secondary-contact of cryptic penguin species. PLoS ONE 2015, 10, e0144966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peucker, A.J.; Dann, P.; Burridge, C.P. Range-wide phylogeography of the little penguin (Eudyptula minor): Evidence of long-distance dispersal. Auk 2009, 126, 397–408. [Google Scholar] [CrossRef]
- Cole, T.L.; Zhou, C.; Fang, M.; Pan, H.; Ksepka, D.T.; Fiddaman, S.R.; Emerling, C.A.; Thomas, D.B.; Bi, X.; Fang, Q. Genomic insights into the secondary aquatic transition of penguins. Nat. Commun. 2022, 13, 3912. [Google Scholar] [CrossRef]
- Collin, H.B.; Collin, S.P. The corneal surface of aquatic vertebrates: Microstructures with optical and nutritional function? Philos. Trans. R. Soc. B 2000, 355, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Pigatto, J.A.; Laus, J.L.; Santos, J.M.; Cerva, C.; Cunha, L.S.; Ruoppolo, V.; Barros, P.S. Corneal endothelium of the Magellanic penguin (Spheniscus magellanicus) by scanning electron microscopy. J. Zoo Wildl. Med. 2005, 36, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.P.; Collin, H.B. The corneal epithelial surface in the eyes of vertebrates: Environmental and evolutionary influences on structure and function. J. Morphol. 2006, 267, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.V.; McGhee, C.N. Contemporary in vivo confocal microscopy of the living human cornea using white light and laser scanning techniques: A major review. Clin. Exp. Ophthalmol. 2007, 35, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Kafarnik, C.; Fritsche, J.; Reese, S. In vivo confocal microscopy in the normal corneas of cats, dogs and birds. Vet. Ophthalmol. 2007, 10, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Kooyman, G.; Cherel, Y.; Le Maho, Y.L.; Croxall, J.; Thorson, P.; Ridoux, V.; Kooyman, C. Diving behavior and energetics during foraging cycles in king penguins. Ecol. Monogr. 1992, 62, 143–163. [Google Scholar] [CrossRef]
- Chiaradia, A.; Ropert-Coudert, Y.; Kato, A.; Mattern, T.; Yorke, J. Diving behaviour of little penguins from four colonies across their whole distribution range: Bathymetry affecting diving effort and fledging success. Mar. Biol. 2007, 151, 1535–1542. [Google Scholar] [CrossRef]
- Croxall, J.P.; Davis, R.W.; Connell, M.J.O. Diving Patterns in Relation to Diet of Gentoo and Macaroni Penguins at South Georgia. Condor 1988, 90, 157–167. [Google Scholar] [CrossRef]
- Miller, A.K.; Karnovsky, N.J.; Trivelpiece, W.Z. Flexible foraging strategies of gentoo penguins Pygoscelis papua over 5 years in the South Shetland Islands, Antarctica. Mar. Biol. 2009, 156, 2527–2537. [Google Scholar] [CrossRef]
- Baker, A.J.; Pereira, S.L.; Haddrath, O.P.; Edge, K.-A. Multiple gene evidence for expansion of extant penguins out of Antarctica due to global cooling. Proc. R. Soc. B 2006, 273, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Vianna, J.A.; Fernandes, F.A.; Frugone, M.J.; Figueiró, H.V.; Pertierra, L.R.; Noll, D.; Bi, K.; Wang-Claypool, C.Y.; Lowther, A.; Parker, P. Genome-wide analyses reveal drivers of penguin diversification. Proc. Natl. Acad. Sci. USA 2020, 117, 22303–22310. [Google Scholar] [CrossRef]
- Masters, B.R.; Böhnke, M. Confocal microscopy of the human cornea in vivo. Int. Ophthalmol. 2001, 23, 199–206. [Google Scholar] [CrossRef]
- Hogan, M.J. Histology of the Human Eye. An Atlas and Textbook; W.B. Saunders Company: Philadelphia, PA, USA, 1971; pp. 55–182. [Google Scholar]
- Shirihai, H. A Complete Guide to Antarctic Wildlife: The Birds and Marine Mammals of the Antarctic Continent and the Southern Ocean, 3rd ed.; A. & C. Black: London, UK, 2007. [Google Scholar]
- Ku, J.Y.; Niederer, R.L.; Patel, D.V.; Sherwin, T.; McGhee, C.N. Laser scanning in vivo confocal analysis of keratocyte density in keratoconus. Ophthalmology 2008, 115, 845–850. [Google Scholar] [CrossRef] [PubMed]
- McCarey, B.E.; Edelhauser, H.F.; Lynn, M.J. Review of corneal endothelial specular microscopy for FDA clinical trials of refractive procedures, surgical devices, and new intraocular drugs and solutions. Cornea 2008, 27, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederer, R.L.; McGhee, C.N.J. Clinical in vivo confocal microscopy of the human cornea in health and disease. Prog. Retin. Eye Res. 2010, 29, 30–58. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, N.; Tani, Y.; Lee, E.; Kikuchi, H.; Endoh, K.; Ichikawa, M.; Sugita, S. Microstructure characteristics of the cornea in birds and mammals. J. Vet. Med. Sci. 2010, 72, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.V.; Ku, J.Y.F.; Johnson, R.; McGhee, C.N.J. Laser scanning in vivo confocal microscopy and quantitative aesthesiometry reveal decreased corneal innervation and sensation in keratoconus. Eye 2009, 23, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Le, Q.; Hong, J.; Xu, J. Comparison of human corneal cell density by age and corneal location: An in vivo confocal microscopy study. BMC Ophthalmol. 2016, 16, 109. [Google Scholar] [CrossRef] [Green Version]
- Petroll, W.M.; Kivanany, P.B.; Hagenasr, D.; Graham, E.K. Corneal fibroblast migration patterns during intrastromal wound healing correlate with ECM structure and alignment. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7352–7361. [Google Scholar] [CrossRef]
- Emmerton, J.; Schwemer, J.; Muth, I.; Schlecht, P. Spectral transmission of the ocular media of the pigeon (Columba livia). Investig. Ophthalmol. Vis. Sci. 1980, 19, 1382–1387. [Google Scholar]
- Williams, T.D.; Briggs, D.R.; Croxall, J.P.; Naito, Y.; Kato, A. Diving pattern and performance in relation to foraging ecology in the gentoo penguin, Pygoscelis papua. J. Zool. 1992, 227, 211–230. [Google Scholar] [CrossRef]
- Rodriguez, C.; Millero, F.J. Modeling the density and isentropic compressibility of seawater. J. Solut. Chem. 2013, 42, 303–316. [Google Scholar] [CrossRef]
- Collin, S.P.; Collin, H.B. A comparative study of the corneal endothelium in vertebrates. Clin. Exp. Optom. 1998, 81, 245–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, R.W.; Edelhauser, H.F.; Stern, M.E. Specular microscopy of vertebrate corneal endothelium: A comparative study. Exp. Eye Res. 1987, 44, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Visser, N.; McGhee, C.N.J.; Patel, D.V. Laser-scanning in vivo confocal microscopy reveals two morphologically distinct populations of stromal nerves in normal human corneas. Br. J. Ophthalmol. 2009, 93, 506–509. [Google Scholar] [CrossRef] [PubMed]
- Piatt, J.F.; Nettleship, D.N. Diving depths of four alcids. Auk 1985, 102, 293–297. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Penguin Species and Identification 1 | Age (Years) | Sex | Preservation Prior to Examination | Right Corneal Curvature 2 (D) and Instrument | Left Corneal Curvature 2 (D) and Instrument | Remarks |
|---|---|---|---|---|---|---|
| Little penguins Eudyptula minor | ||||||
| L1 (B80231) | Adult | Unknown | Fresh 3 | 40.48 IOLMaster | 40.72 IOLMaster | |
| L2 | Adult | Male | 72 h at 4 °C 4, in air | 40.25 Nidek OPD | 40.25 Nidek OPD | |
| Gentoo penguins Pygoscelis papua | ||||||
| G1 (G140, Horse) | 26 | Male | 36 h at 4 °C 4, in normal saline | 26.95 Pentacam | Left eye only | |
| G2 (G198, Stanley) | 26 | Male | 36 h at 4 °C 4, in normal saline | 20.45 Pentacam | 22.55 Pentacam | |
| G3 (G144, Dennis) | 26 | Male | 36 h at 4 °C 4, in normal saline | 22.8 Pentacam | Left eye only | |
| G4 (G194, Twinkle) | 26 | Female | 24 h at 4 °C 4, in air | Right eye used for TEM | ||
| King penguins Aptenodytes patagonicus | ||||||
| K1 (K055, Eskie) | 13 | Female | 36 h at 4 °C 4, in air | 20.22 Nidek OPD | 20.255 Nidek OPD | |
| K2 (K201, no name) | 26 | Male | Fresh 3 | Left eye used for TEM | ||
| K3 (K203, no name) | 26 | Male | Fresh 3 | |||
| K4 (K158) | 30 | Female | 48 h at 4 °C 4, in Optisol culture medium |
| Little Penguin Eudyptula minor | Gentoo penguin Pygoscelis papua | King Penguin Aptenodytes patagonicus | Homogeneity of Variance Test p-Value (Based on Mean) 1 | p-Value One-Way ANOVA 1 | Significance | Human Cornea [24] | |
|---|---|---|---|---|---|---|---|
| Mean superficial epithelial cell density (cells/mm2) | 5756 ± 948 (2) 1 | 5725 ± 455 (4) | 5320 (1) | - | - | No significant difference (2-tailed t test) 2 | n/a |
| Mean basal epithelial cell density (cells/mm2) | 19,795 ± 2837 (3) | 9602 ± 2975 (4) | 16,180 (1) | - | - | p = 0.006 (if normally distributed) or p = 0.007 (if not normally distributed) 2,3 | 5823 ± 602 |
| Mean anterior stromal cell density (cells/mm2) | 447 ± 106 (4) | 193 ± 43 (4) | 288 ± 93 (8) | 0.329 | 0.004 | Little v Gentoo 0.003, Little v King 0.028, Gentoo v King 0.221 | 786 ± 244 |
| Mean mid-stromal cell density (cells/mm2) | 341 ± 113(4) | 132 ± 72 (4) | 231 ± 87 (8) | 0.522 | 0.021 | Little v Gentoo 0.016, Little v King 0.156, Gentoo v King 0.220 | n/a |
| Mean posterior stromal cell density (cells/mm2) | 387 ± 172 (4) | 149 ± 30 (3) | 265 ± 96 (8) | 0.022 | 0.029 | Little v Gentoo 0.133, Little v King 0.456, Gentoo v King 0.029 4 | 293 ± 35 |
| Mean endothelial cell density (cells/mm2) | 1873 ± 369 (2) | 2189 ± 632 (4) | 2001 ± 529 (7) | 0.651 | 0.667 | No significant difference | 2000–3000 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadden, P.W.; Gokul, A.; Amirapu, S.; Kurian, R.; McGhee, C.N.J.; Zhang, J. Confocal and Electron Microscopic Structure of the Cornea from Three Species of Penguin. Vision 2023, 7, 4. https://doi.org/10.3390/vision7010004
Hadden PW, Gokul A, Amirapu S, Kurian R, McGhee CNJ, Zhang J. Confocal and Electron Microscopic Structure of the Cornea from Three Species of Penguin. Vision. 2023; 7(1):4. https://doi.org/10.3390/vision7010004
Chicago/Turabian StyleHadden, Peter W., Akilesh Gokul, Satya Amirapu, Ratish Kurian, Charles N. J. McGhee, and Jie Zhang. 2023. "Confocal and Electron Microscopic Structure of the Cornea from Three Species of Penguin" Vision 7, no. 1: 4. https://doi.org/10.3390/vision7010004