1. Introduction
The African catfish (
Clarias gariepinus, Burchell, 1822) is one of the most popular fish species farmed today, both in and out of its natural habitats [
1]. Due to its excellent nutritional value and fatty acid profile, the demand for
C. gariepinus is increasing. So, many trials have been carried out recently to enhance its growth and health aspects [
2,
3,
4]. Despite
C. gariepinus being a fast-growing species resistant to harsh farming circumstances, infectious diseases may occur when exposed to various disease agents, parasites [
5], and aquatic contaminations [
2], thus inducing production losses [
6].
Globally, herbicides are increasingly used in agriculture with a consequent contamination of the water systems through the water drainage systems, which exert toxicological impacts on the aquatic environment [
4,
7]. Due to its negative impact on living organisms, the contamination of herbicides in aquatic and terrestrial environments has received increased attention to identify the problem and provide solutions. Oxyfluorfen (OXY) is a common pre- and post-emergence herbicide used in vegetable, rice, soybean, and peanut fields to control broad-leaved and annual grass weeds [
8]. The principal consequences of OXY exposure studies in living hosts were protoporphyrinogen oxidase inhibition, heme production suppression, anemia, and liver damage [
9]. Due to several physicochemical characteristics, such as its poor biodegradability, low vapor pressure, and low solubility in water, OXY could pose a major threat to aquatic creatures, including fish [
10]. The half-life of OXY in the soil is between 72 and 150 days [
11], and its presence in fresh water bodies and sediment was recorded [
12,
13]. The bioconcentration of OXY in the organs of fish and whole fishes has been documented [
14].
In freshwater aquaculture systems, the rate of fish development must be continuously monitored during the growing season. Any deviation from the expected growth rate may be a sign of disease, poor water quality, or feeding problems [
15]. A strong indicator of the external changes in the aquatic medium in which fish live is the antioxidant system in fish organs such as the kidney, intestinal tract, and liver, which can be tested biochemically and molecularly [
16]. Moreover, biochemical variables in blood are critical for general health, toxicity, and fish biomonitoring [
17,
18].
The primary line of defense against invasive diseases for fish is their innate immune system, which is more important for fish than mammals [
19]. Lysozyme activity, nitric oxide (NO), and complement 3 are key indicators of fish innate immunological defense [
20,
21]. Cytokines play a crucial role in the immune system via their unique receptors that attach to the cellular membrane, enhancing the cascade-enhancing induction, stimulation, or repression of genes regulated by nuclear cytokines [
22]. Several essential cytokines, including tumor necrosis factor-α (
TNF-α), interleukin-1β (
IL-1β), transforming growth factor-Β (
TGF-Β), interferon (
IFN), and various chemokines, have been identified in teleost fish [
23,
24]. Additionally, a positive correlation between antioxidant enzyme activity (CAT, SOD, GSH, and GPx) and innate immune response in fish has been proven [
25].
Multiple illnesses in fish and other aquatic organisms have been linked to OXY. For example, exposure to OXY in
Paramisgurnus dabryanus resulted in DNA damage [
26]. Similarly, stunted skeletal growth and negative genome-level consequences were observed in several fish species [
27]. Furthermore,
Gambusia affinis and
Oreochromis niloticus had less acetylcholinesterase (AChE) in their brains after exposure to OXY [
28]. Moreover, heat shock protein 70 was upregulated in the kidney and liver of
O. niloticus exposed to OXY [
29]. Exposure to sublethal OXY levels resulted in leukopenia, anemia, lymphopenia, monocytopenia, eosinopenia, and hepato-renal damage in
C. gariepinus [
8,
30] and
O. niloticus [
31]. Accordingly, the need for antistress, antitoxic, and growth enhancers to alleviate the negative impact of stressful conditions has been demonstrated [
32]. Natural plants can serve as better alternatives to antimicrobials in aquaculture, since they have a diverse range of functions, including growth promotion, antioxidant and antimicrobial activities, and their ability to reduce stress [
33,
34].
Artemisia cina (AC), also known as Santonica, wormwood, and worm seed, is a perennial herbaceous plant in the daisy family [
35]. Since ancient times, it has been and continues to be applied in traditional remedies to cure various diseases, including bacterial, fungal and viral infections, cancer, and malaria [
36]. Artemisinin, quercetin, guaianolide, barrelin, capillarisin, artemalin, fisetin, and barrelierin are among the examples of the physiologically active compounds isolated from plants of the
Artemisia genus [
37]. The terpenoid santonin is the most prevalent and well-researched component of this plant. This substance has potent anthelmintic properties [
38]. Additionally, medications containing artemisinin have an antiparasitic capability, and earlier studies have demonstrated artemisinin to be an effective anticancer agent [
39]. Adding AC at 3% and 5% to
C. gariepinus diets considerably enhanced growth and immune response [
40]. AC diet supplementation in concentrations showed efficacy in treating
Gyrodactylus rysavyi of
Cyprinus carpio [
41]. Moreover, the addition of
A. infra at 5% in
C. gariepinus diets was shown to improve innate immune response and hematological parameters while increasing resistance to
Aeromonas hydrophila [
6]. In light of the previous assumptions, this study was designed to define the effects of the co-dietary administration of AC and OXY exposure on
C. gariepinus’s productive performance, physiological and health indicators, and ability to fight against monogeneous parasites of
Quadriacanthus aegypticus. 4. Discussion
This study analyzed the effect of OXY sub-lethal exposure on the growth performance and biological response of catfish,
C. gariepinus, as well as the potential beneficial role of AC dietary inclusion against OXY exposure. Fish exposed to OXY for 60 days displayed substantial growth retardation revealed by reduced FBW, WG, SGR, and feed intake relative to the control group. Similar growth reduction was observed in Nile tilapia after exposure to OXY [
31]. Furthermore, OXY resulted in the stunted skeletal growth of Japanese medaka [
27]. The growth inhibition effect of OXY could be triggered by the negative impact on the health status of exposed fish, a great loss of the digestive cells and decreased digestive enzyme activity [
12], and severe kidney injury combined with the poor health condition of organisms [
30,
72].
Meanwhile, AC dietary supplementation successfully improved the growth parameters and survival of catfish. It also alleviated the negative impact of OXY exposure on zootechnical performance. Comparably, Saleh, Sakr [
40] has shown that AC dietary supplementation can boost the growth performance of
C. gariepinus. AC was recently shown to possess a valuable array of active compounds, such as phenolic compounds, alkaloids, terpenoids, sterols, acetylenes, coumarins, and caffeoylquinic acids [
35]. The AC-growth-stimulating effects may be owed to the phytochemicals which act as growth promoters, such as santonin, betaine, choline, and tannins [
73]. In fish, phytochemicals and their metabolic products may act as prebiotics by selectively inhibiting harmful intestine bacteria and providing growth factors and fermentation substrates for healthy gastrointestinal bacteria [
74]. As a result, farmed fish have improved growth characteristics, feed effectiveness, and resistance to disease. Additionally, the AC-growth-enhancing effect could be correlated to its strong antibacterial effect [
75].
Here, fish exposed to OXY displayed obvious anemic conditions and a substantial decrease in total leukocyte count with a parallel decline in lymphocytes, eosinophils, heterophil, and monocytes. Comparable shifts in hematological parameters have been revealed for OXY-intoxicated
C. gariepinus [
8] and Nile tilapia [
31]. Furthermore, comparable anemic symptoms with various kinds of herbicides such as atrazine [
76], MCPA herbicide [
77], and pendimethalin [
78] have been reported in
C. gariepinus and
C. carpio, respectively. The increased rate of erythrocyte destruction caused by the stress situation imposed by prolonged OXY exposure, which has a detrimental impact on the erythrogram, hastened the unsaturated fatty acids’ peroxidation in the erythrocyte membranes [
31]. It has been noted that OXY prevents human protoporphyrinogen oxidase from producing hemoglobin [
79]. Additionally, the kidney’s interstitium is the primarily hematopoietic system in fish. Therefore, renal injury due to OXY, as presented here, may have caused a decline in the hematological markers. AC dietary supplementation, on the other hand, effectively corrected OXY-induced hematological changes. A similar positive effect of dietary supplementation with
A. infra [
75] and
A. cina [
40] on hematoimmunological indicators has been previously demonstrated in
C. gariepinus. Additionally, Soares, Cardoso [
80] reported that the 30-day dietary supplementation of
A. annua extract increased fish leukocytes and red blood cell function. A rise in RBC, HB, and PCV may indicate a plant’s ability to promote erythropoiesis [
6]. An increased RBC count may be caused by the RBC release from the spleen’s storage pool, which assists the erythrocytes in producing HB [
81]. Furthermore, these favorable outcomes could be linked to augmented oxygen-carrying efficacy, decreased physiological stress, and enhanced health status [
6].
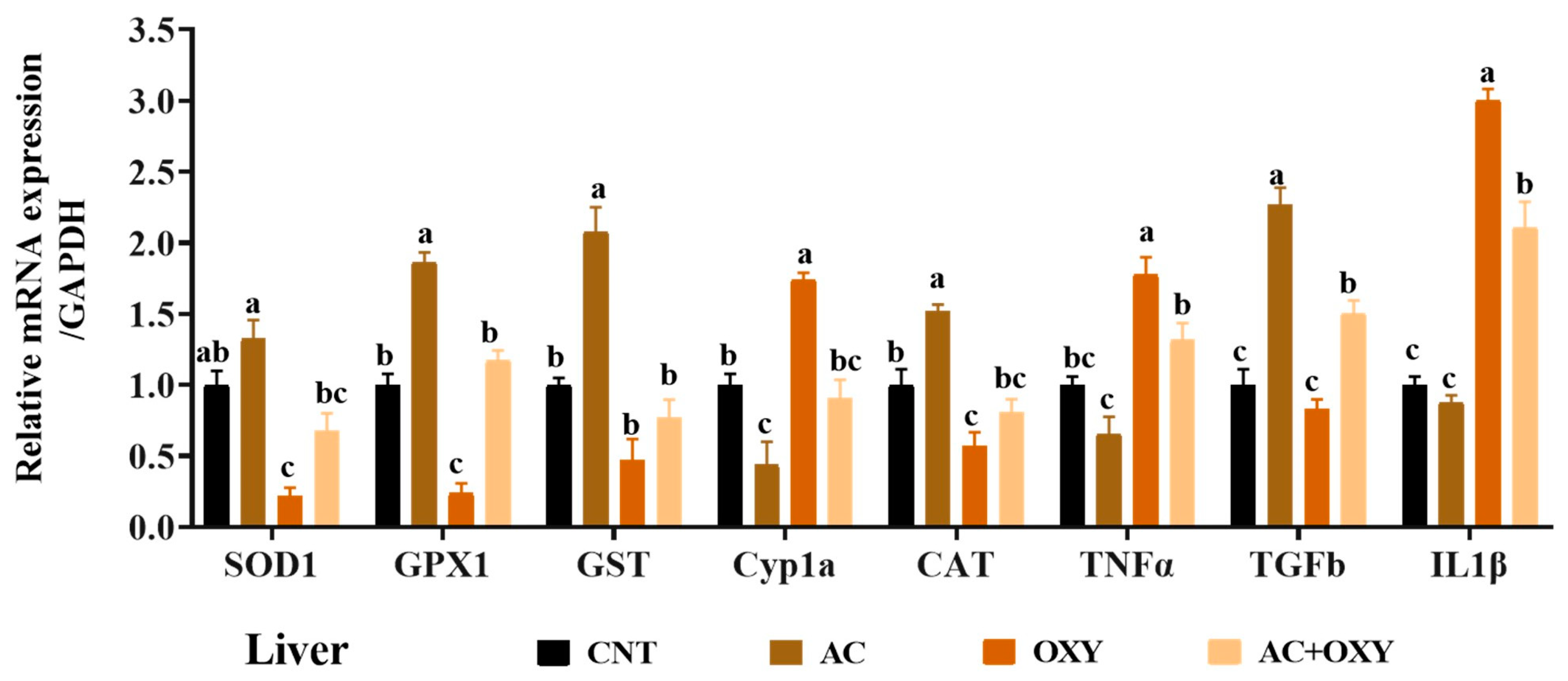
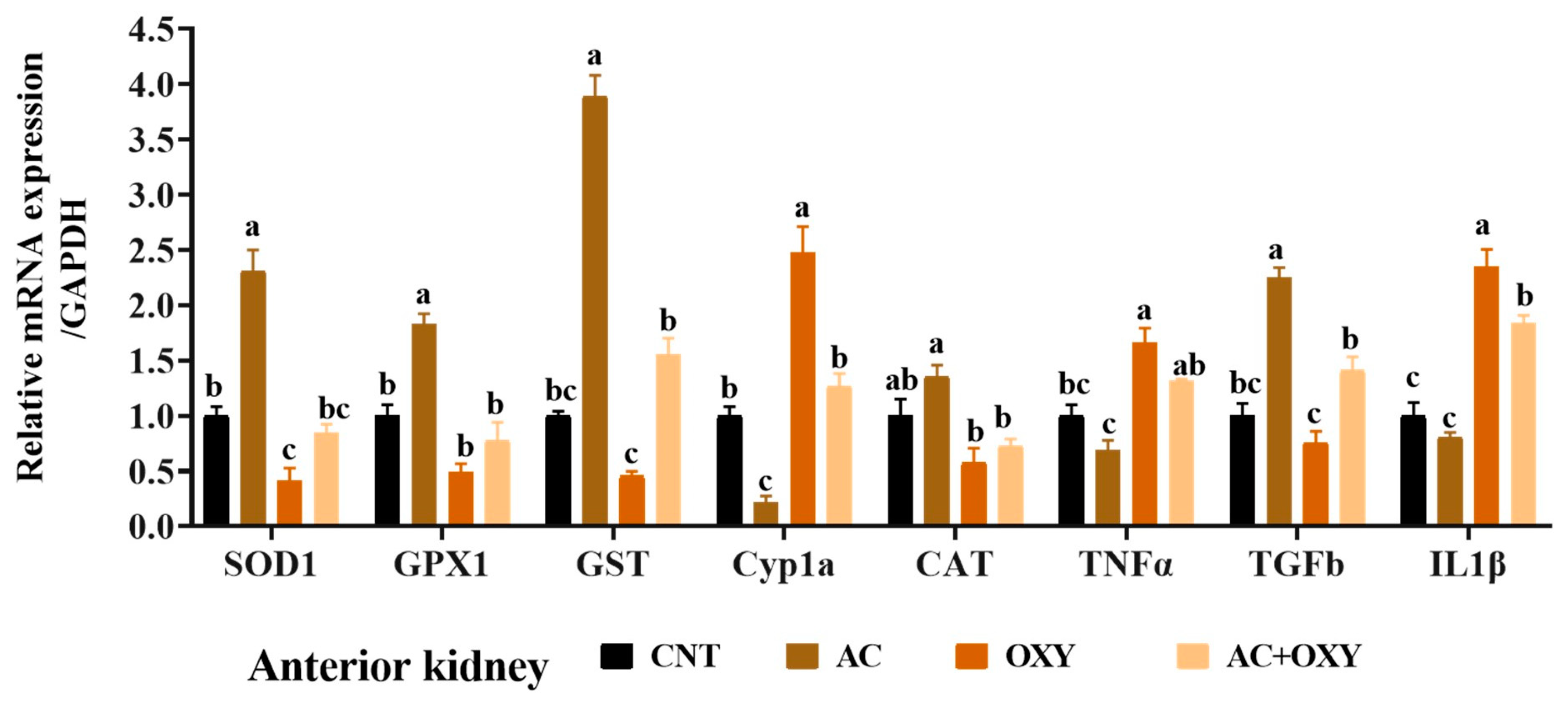
Due to their roles in xenobiotic detoxification, biotransformation, and excretion, the liver and kidneys are the most crucial target organs for most environmental pollutants. Exposure to OXY increases AST, ALT, ALP, creatinine, urea, and bilirubin. When aquatic organisms are exposed to OXY, their livers can bioaccumulate it at high bioconcentration factors of up to 5000, as observed in channel catfish [
8]. Consequently, OXY may disrupt the functional integrity and increase their permeability due to its potent pro-oxidative effects, which could release liver enzymes into the bloodstream and affect liver function [
9]. According to Hassanein, Banhawy [
29], protoporphyrinogen oxidase may be a possible mechanism by which OXY causes hepatorenal damage, as previously demonstrated in mouse kidneys and livers [
82]. Similarly, significant hepatorenal damage due to OXY has been described in
C. gariepinus by Abd El-Rahman, Ahmed [
8] and in tilapia fish by Ibrahim, Ghamry [
31]. Huang, Jia [
72] verified that OXY increased serum levels of kidney injury markers, including creatinine and urea, in zebrafish.
In the present study,
C. gariepinus displayed hypoproteinemia, hypoalbuminemia, hypoglobulinemia, and hypercholesterolemia. Overall, these results point to a fish liver that is less functional, especially in producing blood proteins and controlling lipid metabolism. Furthermore, the reduction in total proteins, albumin, and globulin may be caused by the greater emigration caused by renal failure spurred by exposure to OXY. The results demonstrated that an AC-enriched diet prevents the toxicity of OXY, as evidenced by the above maintenance of the markers at levels similar to or even close to those of the control, confirming the preventive properties of AC on the liver and kidneys. In this regard, Saleh, Sakr [
40] revealed a development pattern of the liver and spleen of
C. gariepinus fed AC.
The results of the current investigation confirmed that fish exposed to OXY exhibited immune-toxic damage and increased fish stress. This was demonstrated by a significant decrease in lysozyme activity, nitric oxide, C3, and phagocytic activity but an increase in cortisol and glucose levels. Likewise, Ibrahim, Ghamry [
31] described a considerable decrease in lysozyme activity and C3 and phagocytic activity following the exposure of
O. niloticus to OXY. Increased cortisol and glucose levels may result from severe liver damage via inhibiting lipid metabolism and glycolysis pathways in the liver [
83]. As opposed to this, fish fed a diet supplemented with AC exhibit improved immune response, enhanced immunological biomarkers, increased survivability, and decreased cortisol levels. Similarly, adding
A. afra leaf powder to the diet of
O. mossambicus increased lysozyme activity [
84]. Moreover, there was an increase in both lysozyme and phagocytic activity in
C. gariepinus fed with 5%
A.
afra [
6]. An earlier report has also shown that AC supplementation at a rate of 5% can boost immune response in
C. gariepinus. An increase in white blood cells indicates that AC inclusion in the diet enhances immunity [
85]. Artemisinin and its derivatives can also actively modify the host’s immune system [
86].
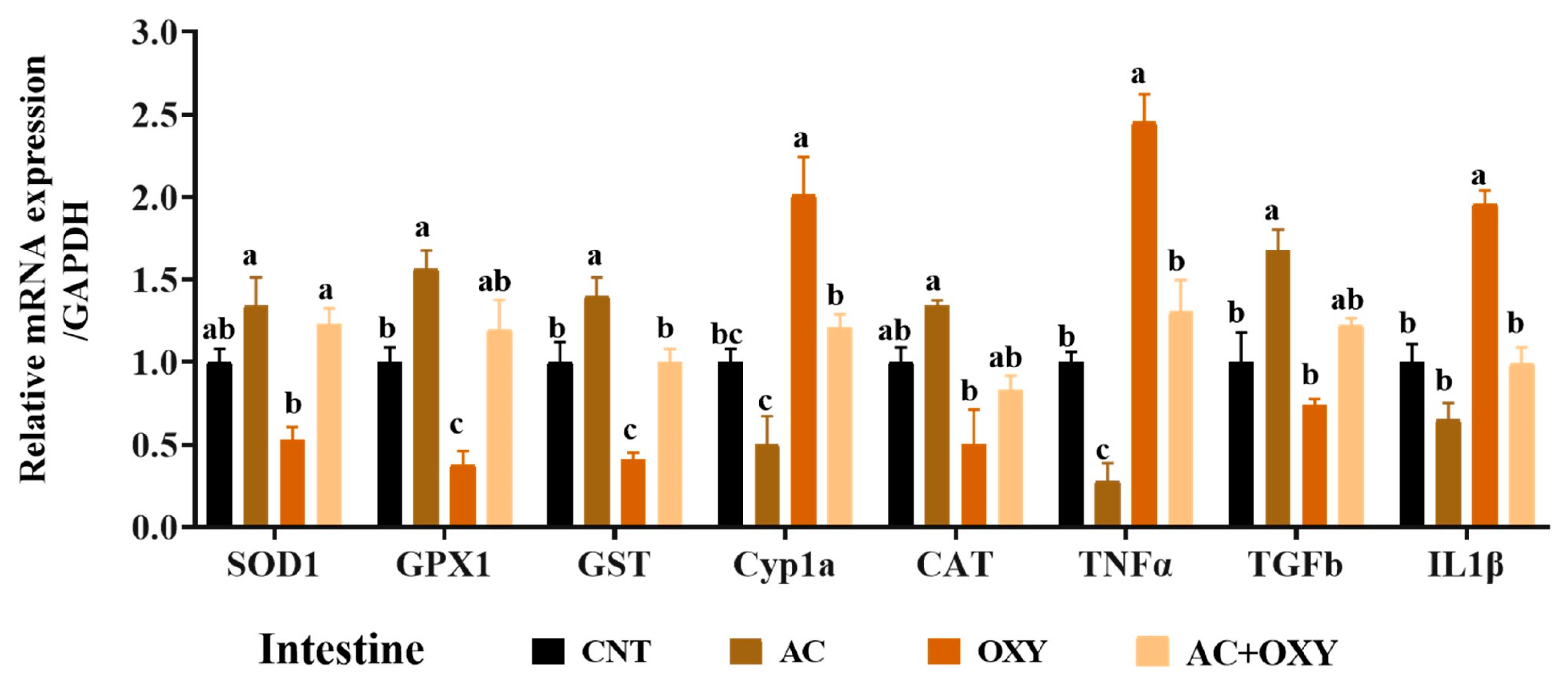
To study the immunotoxic effect of chronic OXY exposure on fish, the mRNA expression of some immune-related genes was assessed in the head kidney, liver, and intestine of
C. gariepinus. The genes analyzed, including TNF-α, IL-1β, and TGF-β1, have been reported to play an important role in the immune defense of
C. gariepinus [
87].
IL-1β and
TNF-α are examples of the inflammatory cytokines generated during inflammation [
88]. The anti-inflammatory cytokine
TGF-β1 reduces inflammation in fish [
89]. The levels of inflammatory and anti-inflammatory cytokines were upregulated in the tissues studied after chronic exposure to OXY. Similarly,
O. niloticus’s toxicity to OXY causes a down-regulation of immune-related gene expression [
31]. In zebrafish, OXY also triggered inflammation by increasing the inflammatory factors
TNF-α, IL-8, and IL-6 [
83]. On the other hand, AC dietary supplementation alone or with OXY significantly decreased the expression of
TNF-α and
IL-1β genes while upregulating
TGF-β1 in contrast to the only impact of OXY exposure. Consequently, the immunostimulant action of AC may have an anti-inflammatory impact as its underlying mechanism. The anti-inflammatory and immunomodulatory effects of sesquiterpene lactones from the Artemisia genus could improve the management of chronic diseases and, consequently, the efficacy of therapy [
90]. Sakipova, Giorno [
36] concluded that AC has probable anti-inflammatory and antinociceptive actions.
According to the results of the numerous assays used to assess the oxidative status, the most likely reason for the reduced organ function in
C. gariepinus intoxicated with OXY was the incidence of oxidative stress damage. The high release of reactive oxygen species (ROS) and free radicals, as well as the incapacity of endogenous antioxidant mechanisms such as CAT, SOD, GST, and GPx to mitigate the harmful effects of these free radicals, are the usual causes of oxidative stress [
91]. Our investigation demonstrated that serum levels of SOD, CAT, GPx, and reduced GSH all significantly decreased after exposure to OXY. Additionally, among all experimental groups, the OXY group had the highest MDA levels, which is a lipid peroxidation marker. Our observations of decreased antioxidant enzyme activity in the serum of
C. gariepinus exposed to OXY matched the outcomes of gene expression analyses. Indeed, we revealed that the liver, kidney, and intestinal tissues of the fish subjected to sublethal OXY dosages considerably decreased the levels of mRNA expression of the genes coding for these antioxidant enzymes (SOD1, CAT, GST, and GPX1). These results indicated the inadequacy of the antioxidant systems to defend against oxidative stress damage triggered by the excessive generation of free radicals in response to exposure to OXY. Similar oxidative stress was described by Abd El-Rahman, Ahmed [
8] in
C. gariepinus and Ibrahim, Ghamry [
31] in
O. niloticus exposed to OXY. The results of the tests used in our investigation to define the level of
Cyp1a gene mRNA expression in the tissues of OXY-intoxicated
C. gariepinus offered additional proof in favor of the development of an oxidative stressful environment in these organs. Additionally, this expression can be related to DNA damage [
92]. The proposed mechanism of immunobiotics that intensify the activities of antioxidant enzymes may increase resistance to toxin exposure [
3]. Interestingly, adding AC, either separately or combined with OXY, enhanced the resistance against the oxidative damage imposed by OXY. The bioactive components of AC, such as thymol, carvacrol, phenolic compounds, betaine, choline, tannins, pigments, essential oil, and santonin, may contribute to its protective impact against oxidative damage induced by OXY [
93]. 8-OHdG is commonly used to diagnose ROS-induced oxidative DNA damage [
94]. In the current investigation, fish exposed to OXY for 60 days had considerably lower levels of AChE and increased 8-OHdG levels. Similarly, significant drops in AChE levels have been observed in the brain of
Gambusia affinis,
O. niloticus, and
C. gariepinus after exposure to sublethal doses of OXY [
8,
28,
31]. The increase in 8-OHdG occurring in serum might have resulted from oxidative stress induced by OXY [
95]. Dietary AC administration restored AChE levels in fish that had been exposed to OXY, perhaps as a result of the supplement’s capacity to mitigate the oxidative stress caused by OXY [
96].
In the current trial, the
C. gariepinus was challenged with
Q. aegypticus gill monogenea after OXY exposure for 60 days. The results showed that the percentage of OXY-exposed fish that survived the challenge reduced significantly. However, the number of infested fish with monogenea in the AC and OXY groups was lower than that in the control. AC’s efficacy as an antiparasitic agent in combating the monogenetic parasite,
Heterobranchus longifilis, has been reported in
C. carpio [
41]. The World Health Organization recommends the use of artemisinin to treat malaria, and this plant is the only known source of this compound [
97]. Artemisinin attacks the parasite’s mitochondria, which are essential for the parasite’s effective functioning [
96]. Additionally, the main component of wormseed is the crystalline primary, santonin, from which the drug’s anthelmintic effects are derived. The immunomodulatory, anti-inflammatory, antimicrobial, and antiulcer activities help in promoting the health of fish, and consequently, the relative survival rate. Exposure to OXY could disrupt parasite transmission via direct or indirect effects on hosts and parasites; this can explain the decreased number of infested fish and prevalence percentage. Sublethal exposure to OXY significantly altered the health state of intoxicated fish in our study, and these changes had a deleterious effect on the general well-being, and consequently, survival rate.


 ,
, 




{kind=link}
{kind=link}
{kind=link}