
Changes in the Gut Microbiota Composition of Juvenile Olive Flounder (Paralichthys olivaceus) Caused by Pathogenic Bacterial Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogenic Bacterial Strains and Culture Conditions
2.2. Animals, Sampling, and Infection Experiment
2.3. Intestinal Microbiota Analysis
2.4. Statistical Analysis
3. Results
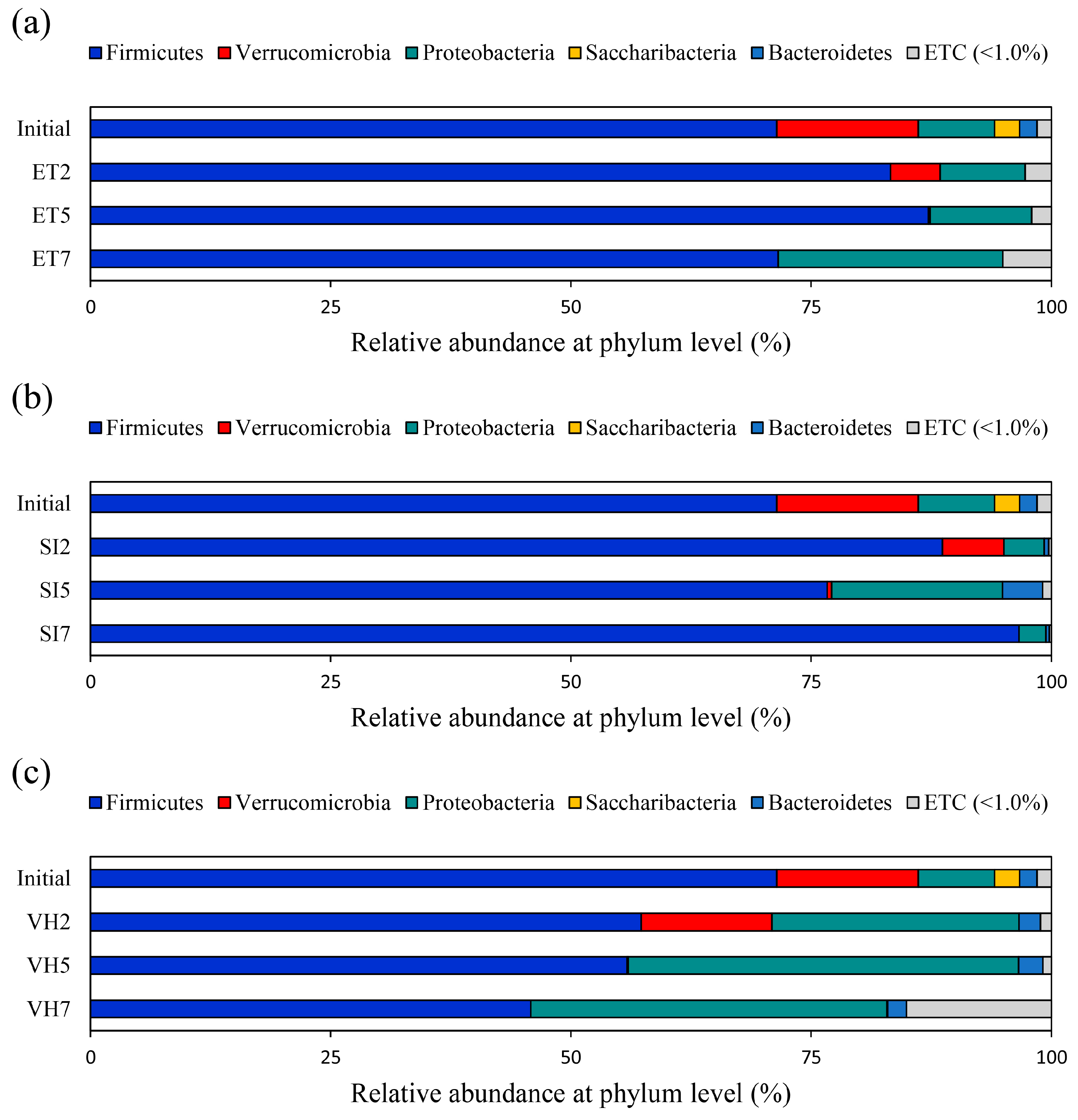
3.1. Changes in Gut Microbiota Composition at the Phylum Level
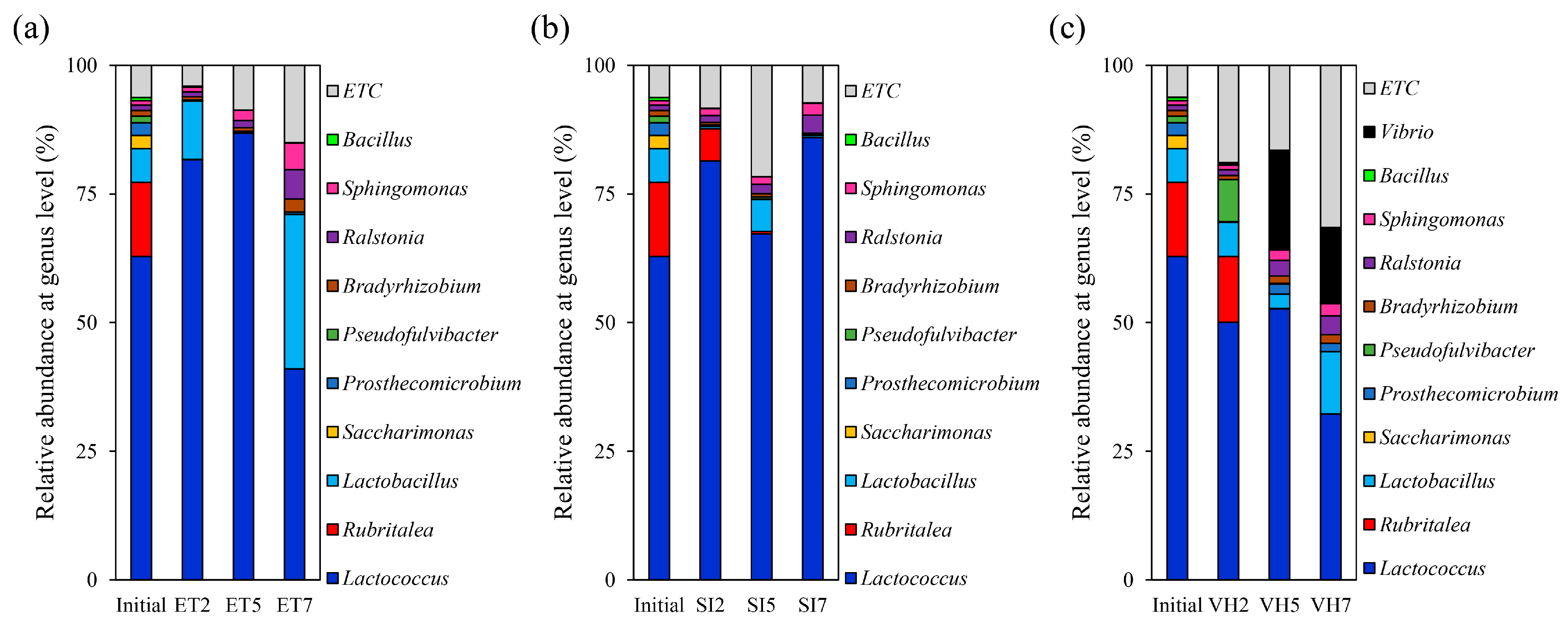
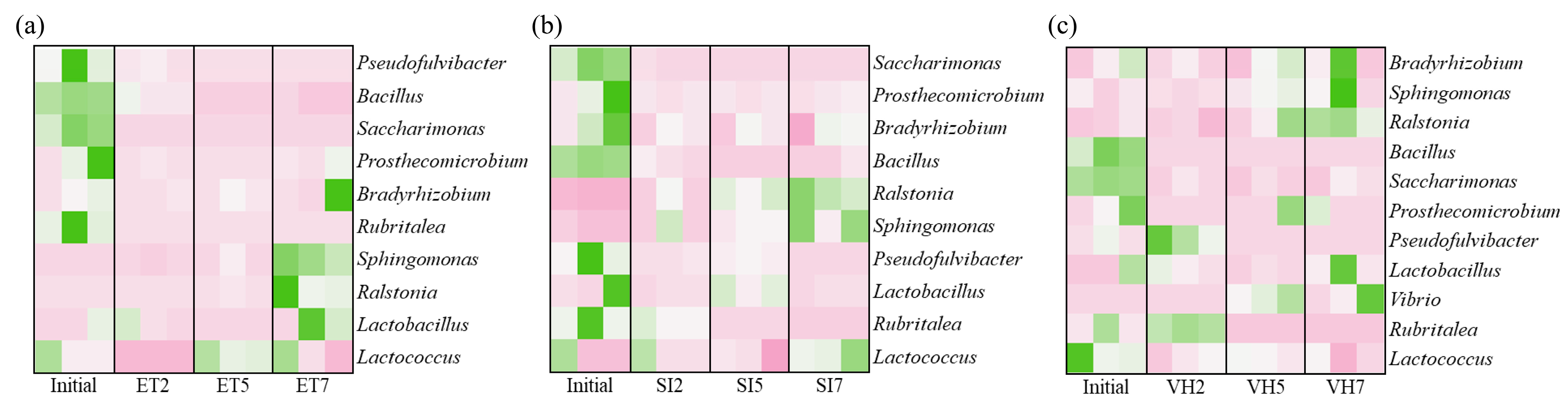
3.2. Changes in Gut Microbiota Composition at the Genus Level
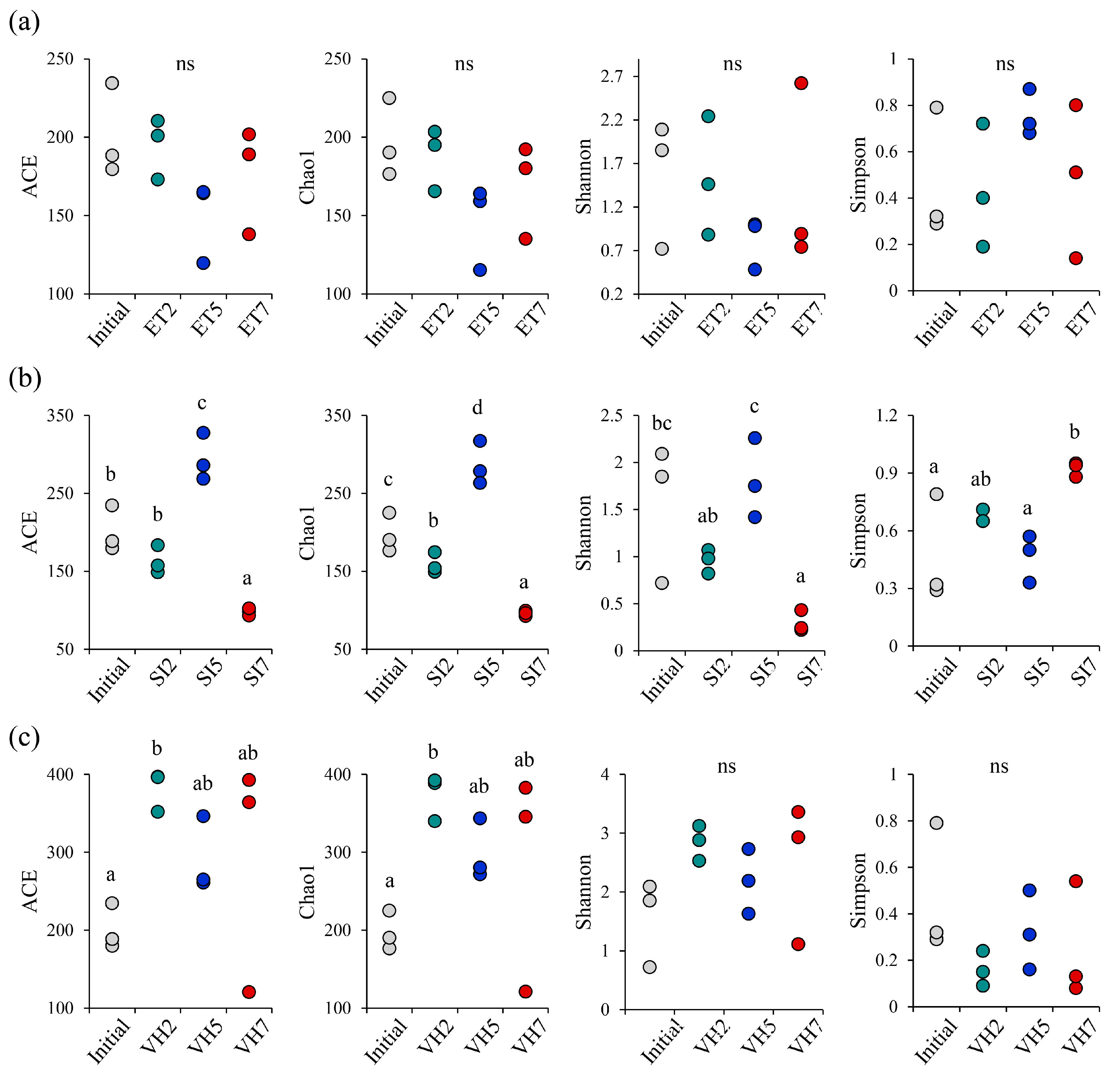
3.3. α-Diversity Changes with Pathogen Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Korean Statistical Information Service (KOSIS), Survey on the Status of Fish Culture. 2021. Available online: http://kosis.kr/ (accessed on 24 March 2022).
- Shim, J.D.; Hwang, S.D.; Jang, S.Y.; Kim, T.W.; Jeong, J.M. Monitoring of the mortalities in oliver flounder (Paralichthys olivaceus) farms of Korea. J. Fish Pathol. 2019, 32, 29–35. [Google Scholar] [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Treves-Brown, K.M. Applied Fish Pharmacology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 3. [Google Scholar]
- Hasan, M.T.; Jang, W.J. Effects of immunostimulants, prebiotics, probiotics, synbiotics, and potentially immunoreactive feed additives on olive flounder (Paralichthys olivaceus) aquaculture: A review. Rev. Fish. Sci. Aquac. 2019, 27, 417–437. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J. Synergistic effects of dietary Bacillus sp. SJ-10 plus β-glucooligosaccharides as a synbiotic on growth performance, innate immunity and streptococcosis resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish. Immunol. 2018, 82, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.J.; Jeon, M.H. Dietary supplementation of Bacillus sp. PM8313 with β-glucan modulates the intestinal microbiota of red sea bream (Pagrus major) to increase growth, immunity, and disease resistance. Front. Immunol. 2022, 13, 960554. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Sritunyalucksana, K.; Flegel, T.W.; Williams, B.A.; Withyachumnarnkul, B.; Itsathitphaisarn, O.; Bass, D. New paradigms to help solve the global aquaculture disease crisis. PLOS Pathog. 2017, 13, e1006160. [Google Scholar] [CrossRef]
- Seo, B.S.; Park, S.J.; Hwang, S.Y.; Lee, Y.I.; Lee, S.H.; Hur, S.W.; Lee, K.J.; Nam, T.J.; Song, J.W.; Kim, J.S.; et al. Effects of decreasing fishmeal as main source of protein on growth, digestive physiology, and gut microbiota of olive flounder (Paralichthys olivaceus). Animals 2022, 12, 2043. [Google Scholar] [CrossRef]
- Ramírez, C.; Romero, J. Fine flounder (Paralichthys adspersus) microbiome showed important differences between wild and reared specimens. Front. Microbiol. 2017, 8, 271. [Google Scholar] [CrossRef]
- Kim, P.S.; Shin, N.R.; Lee, J.B.; Kim, M.S.; Whon, T.W.; Hyun, D.W.; Yun, J.H.; Jung, M.J.; Kim, J.Y.; Bae, J.W. Host habitat is the major determinant of the gut microbiome of fish. Microbiome 2021, 9, 166. [Google Scholar] [CrossRef]
- Jang, W.J.; Lee, S.J.; Jeon, M.H.; Kim, T.Y.; Lee, J.M.; Hasan, M.T.; Lee, H.T.; Park, J.H.; Lee, B.J.; Hur, S.W.; et al. Characterization of a Bacillus sp. KRF-7 isolated from the intestine of rockfish and effects of dietary supplementation with mannan oligosaccharide in rockfish aquaculture. Fish Shellfish Immunol. 2021, 119, 182–192. [Google Scholar] [CrossRef]
- Jang, W.J.; Hasan, M.T.; Choi, W.; Hwang, S.; Lee, Y.; Hur, S.W.; Lee, S.H.; Lee, B.J.; Choi, Y.H.; Lee, J.M. Comparison of growth performance, non-specific immunity, and intestinal microbiota of olive flounder (Paralichthys olivaceus) fed with extruded pellet and moist pellet diets under field conditions in South Korea. Front. Microbiol. 2022, 13, 979124. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Mekasha, S.; Linke, D. Secretion systems in gram-negative bacterial fish pathogens. Front. Microbiol. 2021, 12, 782673. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, X.; Zhao, F.; Zhou, Z.; Zhao, F.; Wang, J.; Liu, T.; Yang, X.; Zhang, X.; Li, Z. Stress responses of the intestinal digestion, antioxidant status, microbiota and non-specific immunity in Songpu mirror carp (Cyprinus carpio L.) under starvation. Fish Shellfish Immunol. 2022, 120, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.T.; Xiong, F.; Hao, Y.T.; Zhang, J.; Wu, S.G.; Wang, G.T. Starvation influences the microbiota assembly and expression of immunity-related genes in the intestine of grass carp (Ctenopharyngodon idellus). Aquaculture 2018, 489, 121–129. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Palladino, G.; Rampelli, S.; Scicchitano, D.; Musella, M.; Quero, G.M.; Prada, F.; Mancuso, F.; Seyfarth, A.M.; Turroni, S.; Candela, M.; et al. Impact of marine aquaculture on the microbiome associated with nearby holobionts: The case of Patella caerulea living in proximity of Sea Bream aquaculture cages. Microorganisms 2021, 9, 455. [Google Scholar] [CrossRef]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: Factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- van Kessel, M.A.; Dutilh, B.E.; Neveling, K.; Kwint, M.P.; Veltman, J.A.; Flik, G.; Jetten, M.S.; Klaren, P.H.; Op den Camp, H.J. Pyrosequencing of 16S rRNA gene amplicons to study the microbiota in the gastrointestinal tract of carp (Cyprinus carpio L.). AMB Express 2011, 1, 41. [Google Scholar] [CrossRef]
- Ryan, M.P.; Adley, C.C. Ralstonia spp.: Emerging global opportunistic pathogens. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 291–304. [Google Scholar] [CrossRef]
- Carda-Diéguez, M.; Mira, A.; Fouz, B. Pyrosequencing survey of intestinal microbiota diversity in cultured sea bass (Dicentrarchus labrax) fed functional diets. FEMS Microbiol. Ecol. 2014, 87, 451–459. [Google Scholar] [CrossRef]
- Wu, S.; Gao, T.; Zheng, Y.; Wang, W.; Cheng, Y.; Wang, G. Microbial diversity of intestinal contents and mucus in yellow catfish (Pelteobagrus fulvidraco). Aquaculture 2010, 303, 1–7. [Google Scholar] [CrossRef]
- Kim, D.H.; Brunt, J.; Austin, B. Microbial diversity of intestinal contents and mucus in rainbow trout (Oncorhynchus mykiss). J. Appl. Microbiol. 2007, 102, 1654–1664. [Google Scholar] [CrossRef] [PubMed]
- Çanak, Ö.; AKAYLI, T.; Çiğdem, Ü.R.K.Ü. A mixed Bacillus gibsonii and Sphingomonas echinoides infection in cultured rainbow trout (Oncorhynchus mykiss). Mar. Life Sci. 2021, 3, 71–79. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-G.; Lee, S.-J.; Lee, J.M.; Lee, E.-W.; Jang, W.J. Changes in the Gut Microbiota Composition of Juvenile Olive Flounder (Paralichthys olivaceus) Caused by Pathogenic Bacterial Infection. Fishes 2023, 8, 294. https://doi.org/10.3390/fishes8060294
Kim D-G, Lee S-J, Lee JM, Lee E-W, Jang WJ. Changes in the Gut Microbiota Composition of Juvenile Olive Flounder (Paralichthys olivaceus) Caused by Pathogenic Bacterial Infection. Fishes. 2023; 8(6):294. https://doi.org/10.3390/fishes8060294
Chicago/Turabian StyleKim, Dong-Gyun, Su-Jeong Lee, Jong Min Lee, Eun-Woo Lee, and Won Je Jang. 2023. "Changes in the Gut Microbiota Composition of Juvenile Olive Flounder (Paralichthys olivaceus) Caused by Pathogenic Bacterial Infection" Fishes 8, no. 6: 294. https://doi.org/10.3390/fishes8060294