
Age and Growth of the Threatened Smalleye Round Ray, Urotrygon microphthalmum, Delsman, 1941, from Northeastern Brazil
 , and
, and
Abstract
:1. Introduction
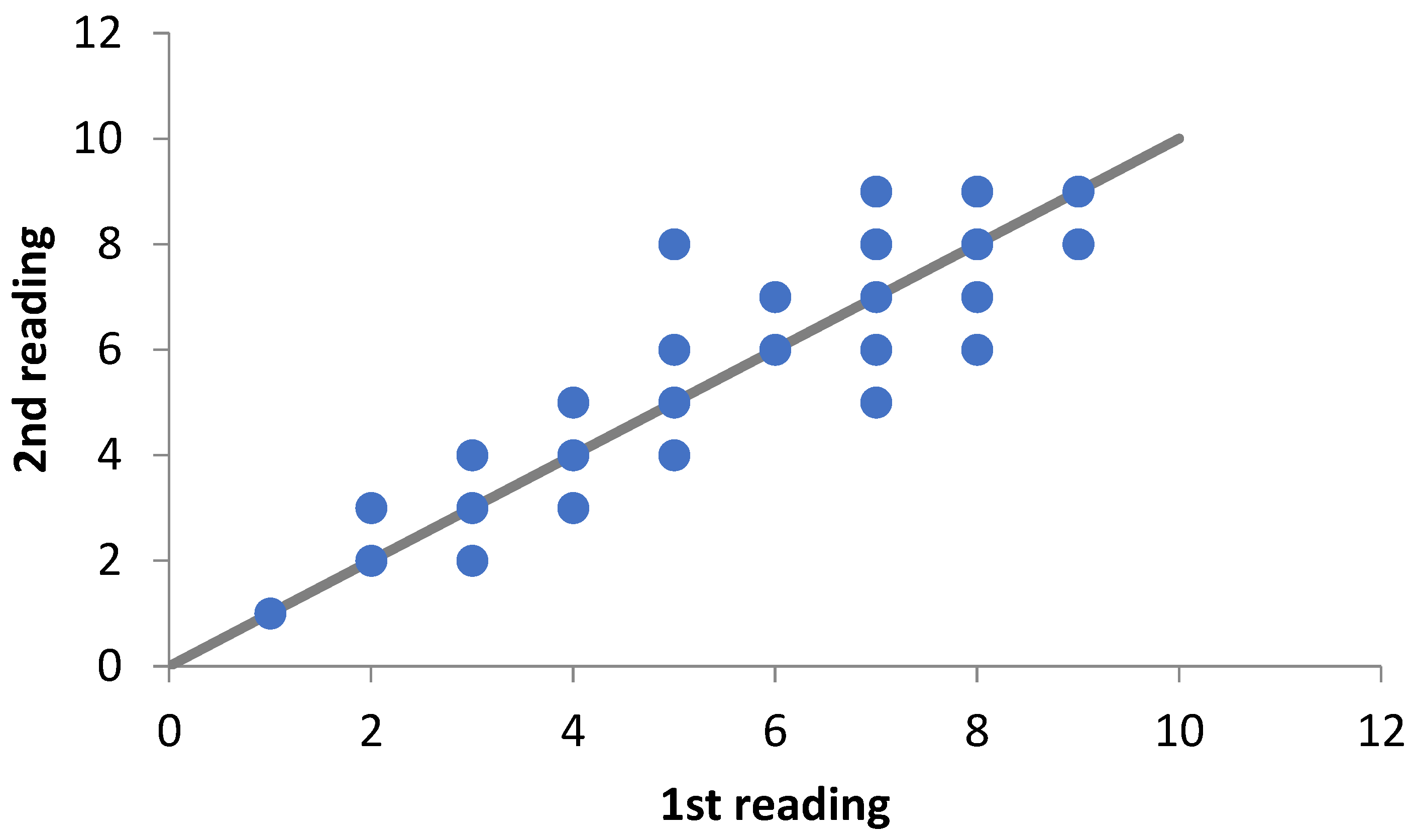
2. Materials and Methods
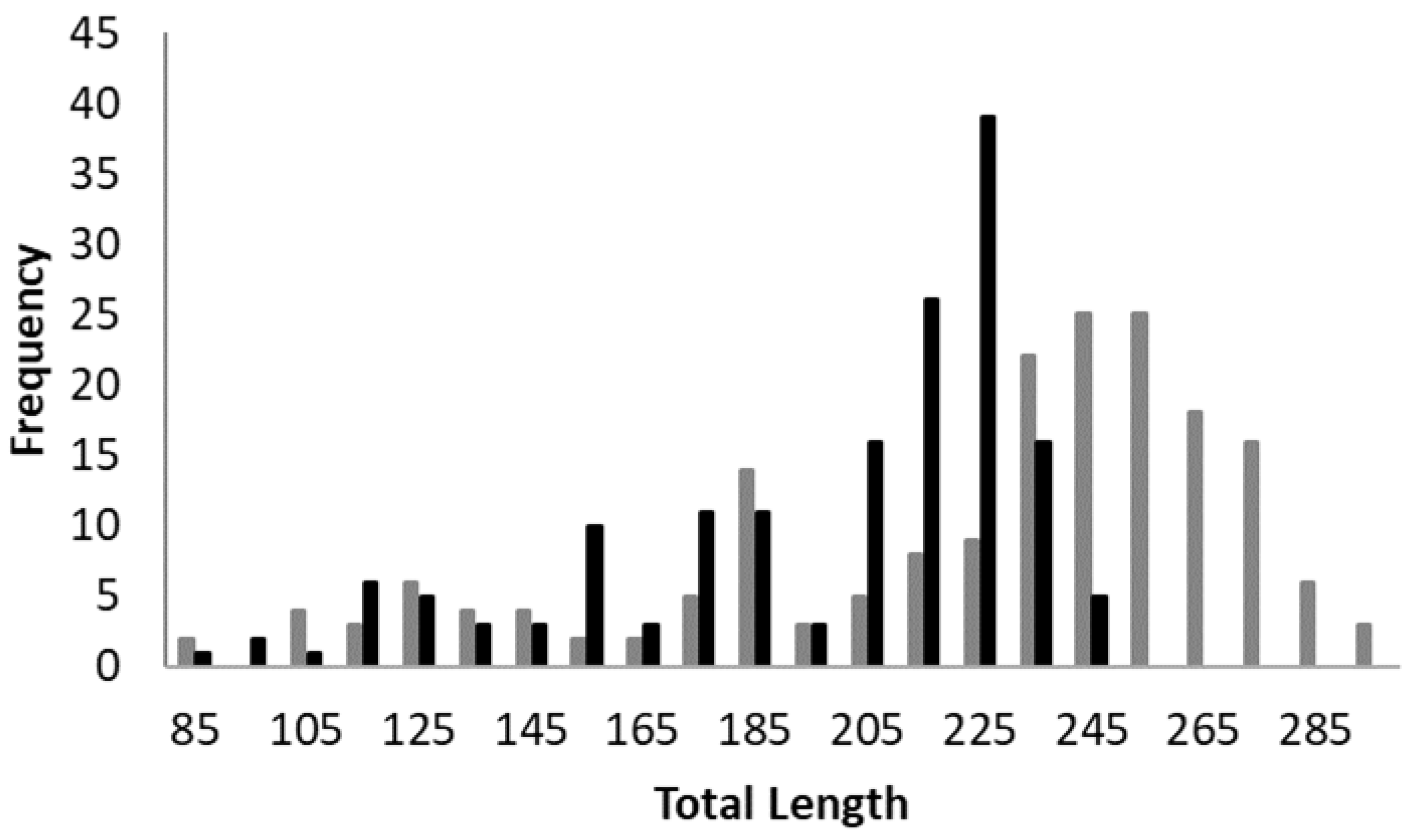
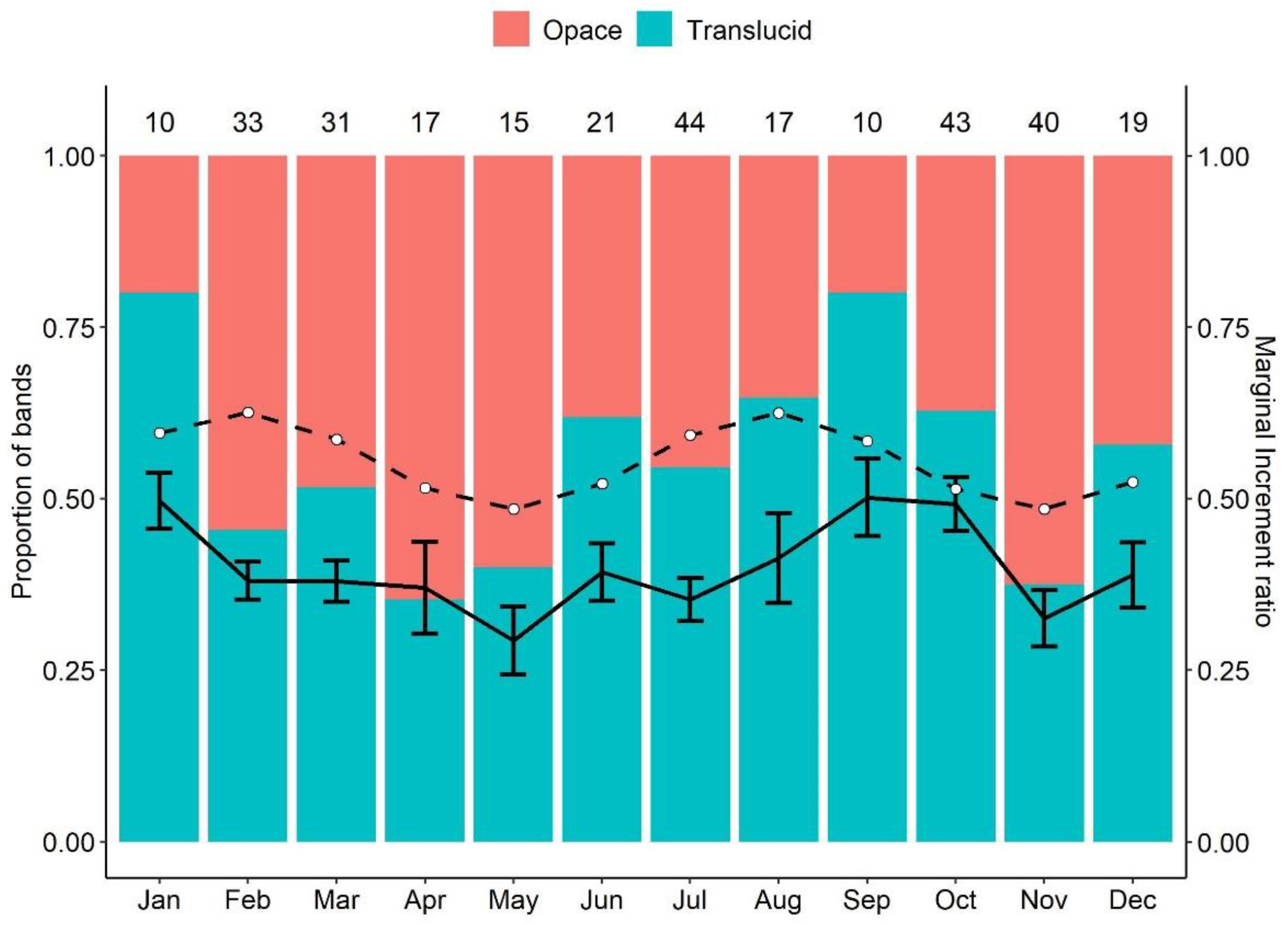
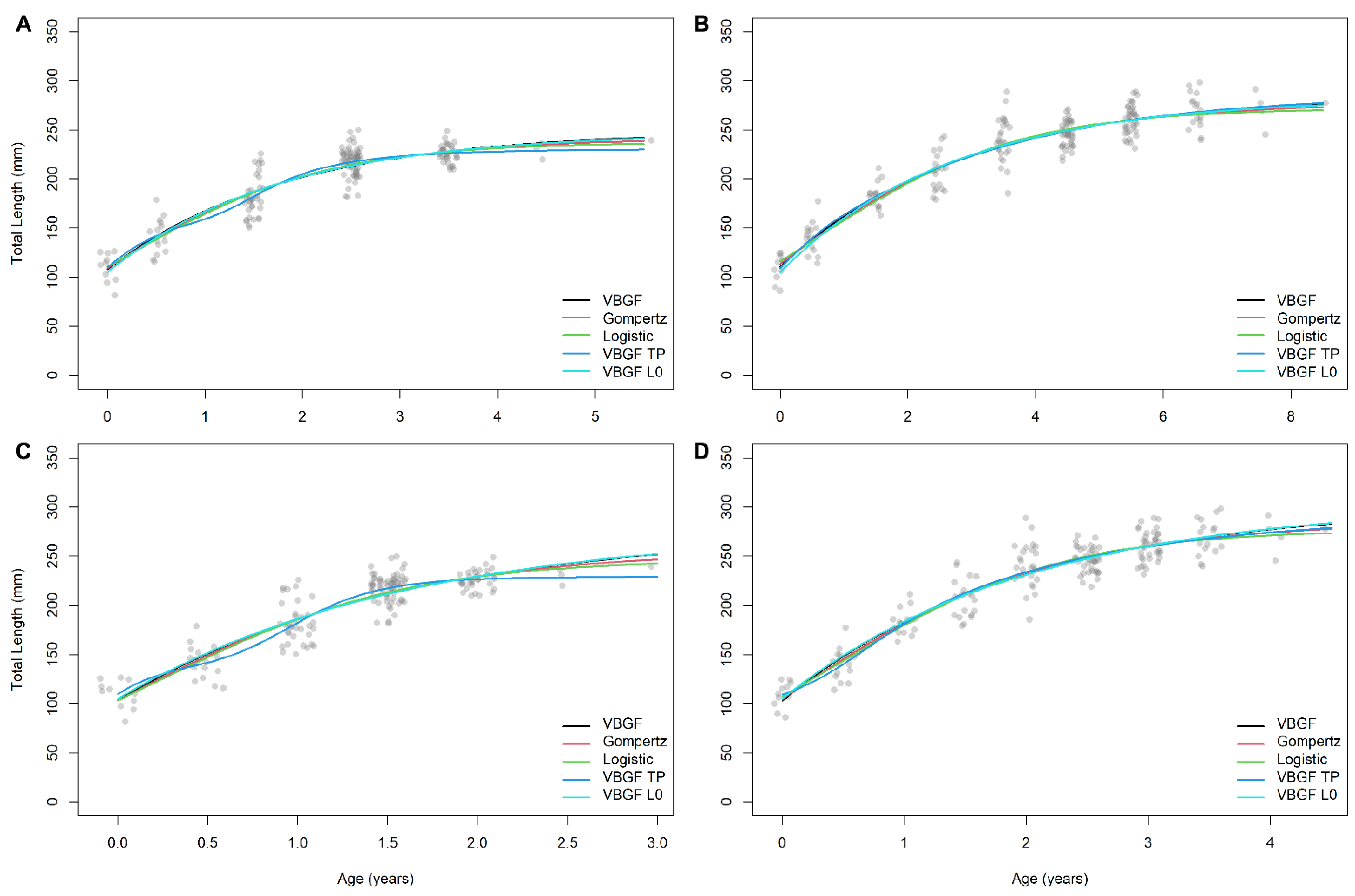
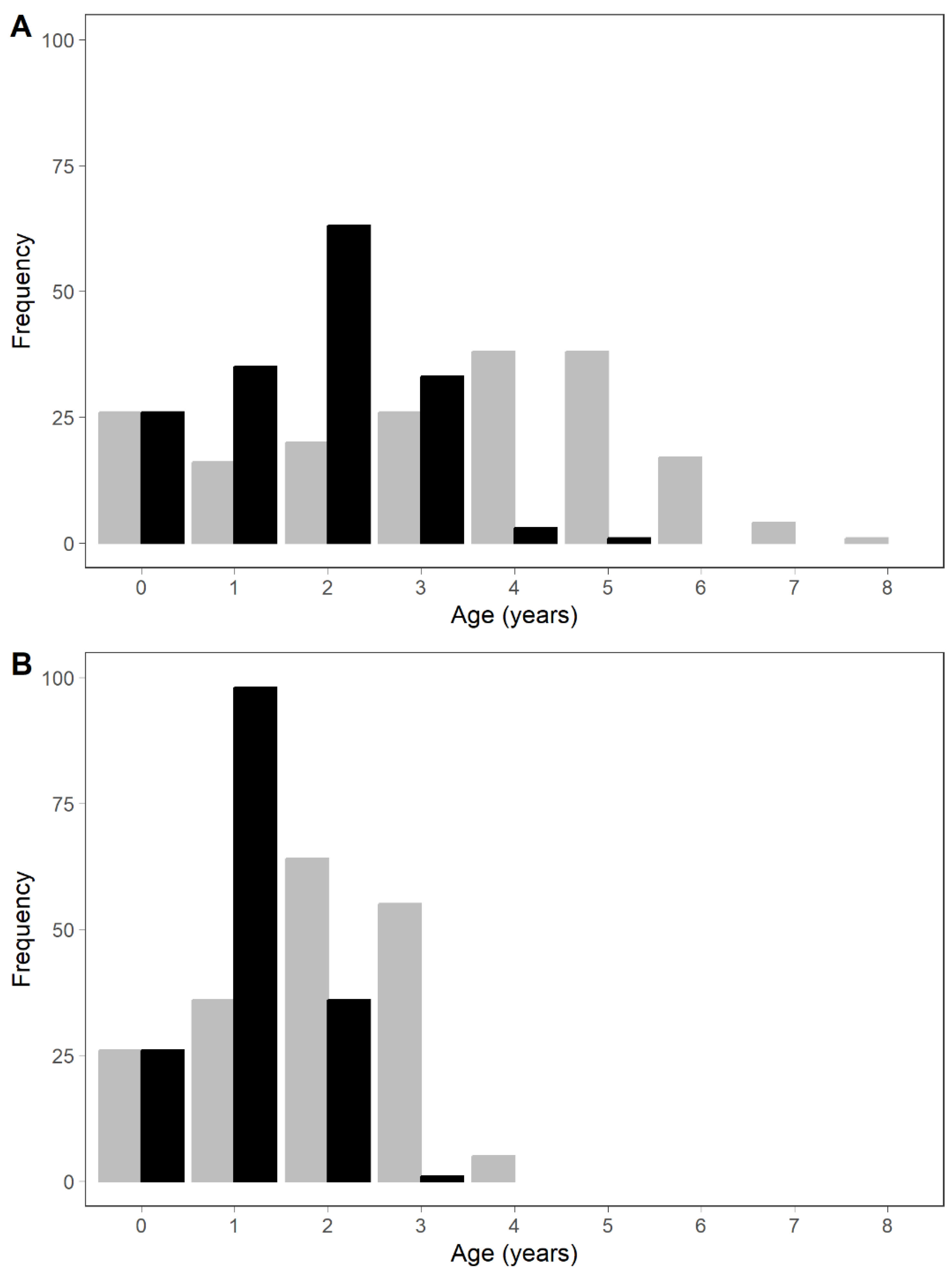
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Last, P.; Naylor, G.; Séret, B.; White, W.; de Carvalho, M.; Stehmann, M. Rays of the World; CSIRO Publishing: Clayton, Australia, 2016. [Google Scholar]
- Ebert, D.A.; Dando, M.; Fowler, S. Sharks of the World: A Complete Guide; Princeton University Press: Princeton, UK, 2021; 607p. [Google Scholar]
- Cailliet, G.M.; Goldman, K.J. Age determination and validation in chondrichthyan fishes. In Biology of Sharks and Their Relatives; Carrier, J., Musick, J.A., Heithaus, M.R., Eds.; CRC Press LLC: Boca Raton, FL, USA, 2004; pp. 399–447. [Google Scholar]
- Torres-Palacios, K.; Mejia-Falla, P.A.; Navia, A.F.; Cruz-Escalona, V.H.; Felix-Uraga, R.; Quinonez-Velazquez, C. Age and growth parameters of the Panamic stingray (Urotrygon aspidura). Fish. Bull. 2019, 117, 169–180. [Google Scholar] [CrossRef]
- McEachran, J.D.; Carvalho, M.R. Urotrygonidae. In The Living Marine Resources of the Western Central Atlantic. Volume 1: Introduction, Molluscs, Crustaceans, Hagfishes, Sharks, Batoid Fishes, and Chimaeras; Carpenter, K.E., Ed.; FAO Species Identification Guide for Fishery Purposes and American Society of Ichthyologists and Herpetologists Special Publication No. 5; FAO: Rome, Italy, 2000; pp. 572–574. [Google Scholar]
- Boeseman, M. Notes on new or rare marine fishes from Surinam. Zool. Mededelin. 1963, 38, 295–301. [Google Scholar]
- Cervigón, F.; Alcalá, A. Los Peces Marinos de Venezuela. Volume 5. Tiburones y Rayas; Fundación Museo del Mar: Ceuta, Spain, 1999. [Google Scholar]
- Almeida, Z.; Nunes, J.; Costa, C. Presencia de Urotrygon microphthalmum (Elasmobranchii: Urolophidae) en aguas bajas de Maranhão (Brasil) y notas sobre su biología. Bol. Invest. Mar. Cost. 2000, 29, 67–72. [Google Scholar] [CrossRef]
- Gadig, O.B.F.; Bezerra, M.A.; Feitosa, R.D.; Furtado-Neto, M.A.A. Ictiofauna marinha do Estado do Ceará, Brasil: I. Elasmobranchii. Arq. Ciências Mar 2000, 33, 127–132. [Google Scholar]
- Rosa, R.S. Urotrygon microphthalmum. In IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. Available online: www.iucnredlist.org (accessed on 30 September 2011).
- Santander-Neto, J.; Lessa, R.P.T. Hermaphroditic smalleyed roundray (Urotrygon microphthalmum) from north-eastern Brazil. Mar. Biodivers. Rec. 2013, 6, e60. [Google Scholar] [CrossRef]
- Garcia-Júnior, J.; Nóbrega, M.F.; Oliveira, J.E.L. Coastal fishes of Rio Grande do Norte, northeastern Brazil, with new records. CheckList 2015, 11, 1659. [Google Scholar] [CrossRef] [Green Version]
- Santander-Neto, J.; Araújo, M.L.G.; Lessa, R.P. Reproductive biology of Urotrygon microphthalmum (Baitodea: Urotrygonidae) from North-East Brazil, tropical west Atlantic Ocean. J. Fish Biol. 2016, 89, 1026–1042. [Google Scholar] [CrossRef] [PubMed]
- Santander-Neto, J.; Freitas, D.J.V.; Bornatowski, H.; Lessa, R. Feeding habits of Urotrygon microphthalmum caught off northeastern Brazil. Neotrop. Ichthyol. 2021, 19, e210039. [Google Scholar] [CrossRef]
- Valadez–González, C.; Aguilar–Palomino, B.; Hernández–Vázquez, S. Hábitos alimentarios de la raya Urobatis halleri (Cooper 1863) (Chondrichthyes: Urolophidae) capturada en la plataforma continental de Jalisco y Colima, México. Cienc. Mar. 2001, 27, 91–104. [Google Scholar] [CrossRef]
- Amezcua, F.; Madrid-Vera, J.; Aguirre-Villaseñor, H. Efecto de la pesca artesanal de camarón sobre la ictiofauna en el sistema lagunar de Santa María la Reforma, suroeste del Golfo de California. Cienc. Mar. 2006, 32, 97–109. [Google Scholar] [CrossRef]
- Acevedo, K.; Bohórquez-Herrera, J.; Moreno, F.; Moreno, C.; Molina, E.; Grijalba-Bendeck, M.; Gómez-Canchong, P. Tiburones y rayas (subclase Elasmobranchii) descartados por la flota de arrastre camaronero en el Caribe de Colombia. Acta Biol. Colomb. 2007, 12, 7–81. [Google Scholar]
- Mejía-Falla, P.A.; Navia, A.F.; Cortés, E. Reproductive variables of Urotrygon rogersi (Batoidea: Urotrygonidae): A species with a triannual reproductive cycle in the Eastern Tropical Pacific Ocean. J. Fish Biol. 2012, 80, 1246–1266. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, M.K. Modifications to reduce by-catch in prawn trawls: A review and framework for development. Rev. Fish Biol. Fisher 2000, 10, 27–60. [Google Scholar] [CrossRef]
- Goldman, K.G. Age and growth of elasmobranch fishes. In Management Techniques for Elasmobranch Fisheries; Musick, J.A., Bonfil, R., Eds.; FAO Fisheries Technical Paper Number 474′; FAO: Rome, Italy, 2005; pp. 76–102. [Google Scholar]
- Cortés, E. Stock Assessment of Small Coastal Sharks in the U.S. Atlantic and Gulf of Mexico; Sustainable Fisheries Division Contribution SFD-01/02-152; NOAA Fisheries: Panama City, FL, USA, 2002. [Google Scholar]
- White, W.T.; Platell, M.E.; Potter, I.C. Relationship between reproductive biology and age composition and growth in Urolophus lobatus (Batoidea:Urolophidae). Mar. Biol. 2001, 138, 135–147. [Google Scholar] [CrossRef]
- White, W.T.; Hall, N.G.; Potter, I.C. Reproductive biology and growth during pre and postnatal life of Trygonoptera personata and T. mucosa (Batoidea:Urolophidae). Mar. Biol. 2002, 140, 699–712. [Google Scholar]
- White, W.T.; Potter, I.C. Reproductive biology, size and age compositions and growth of the batoid Urolophus paucimaculatus, including comparisons with other species of the Urolophidae. Mar. Freshw. Res. 2005, 56, 101–110. [Google Scholar] [CrossRef]
- Hale, L.F.; Lowe, C.G. Age and growth of the round stingray Urobatis halleri at Seal Beach California. J. Fish Biol. 2008, 73, 510–523. [Google Scholar] [CrossRef]
- Mejía-Falla, P.A.; Cortés, E.; Navia, A.F.; Zapata, F.A. Age and Growth of the Round Stingray Urotrygon rogersi, a Particularly Fast-Growing and Short-Lived Elasmobranch. PLoS ONE 2014, 9, e96077. [Google Scholar] [CrossRef]
- Frisk, M.G. Life history strategies of batoids. In Sharks and Their Relatives II: Biodiversity, Adaptive Physiology and Conservation; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: New York, NY, USA, 2010; pp. 283–316. [Google Scholar]
- Cortés, E. Life history patterns and correlations in sharks. Rev. Fish. Sci. 2000, 8, 299–344. [Google Scholar] [CrossRef]
- MMA. Portaria nº 300, de 13 de Dezembro de 2022. Ministério do Meio Ambiente/Gabinete do Ministro. Diário Oficial da República Federativa do Brasil, Poder Executivo, Brasília, DF, 2022. DOU de 14/12/2022 (edição 234, Seção 1, p. 75). Available online: https://www.in.gov.br/en/web/dou/-/portaria-sap/mapa-n-159-de-10-de-maio-de-2021-319027169 (accessed on 23 December 2022).
- Pollom, R.; Charvet, P.; Faria, V.; Herman, K.; Lasso-Alcalá, O.; Marcante, F.; Nunes, J.; Rincon, G. Urotrygon microphthalmum. The IUCN Red List of Threatened Species 2020: E.T44598A2998129. Available online: https://www.iucnredlist.org/species/44598/2998129 (accessed on 3 March 2021).
- Kempf, M. A plataforma continental de Pernambuco (Brasil): Nota preliminar sobre a natureza do fundo. Trab. Oceanogr. Univers. Fed. Pernamb 1970, 67/69, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Manso, V.A.V.; Corrêa, I.C.S.; Guerra, N.C. Morfologia e sedimentologia da Plataforma Continental Interna entre as praias Porto de Galinhas e Campos-Litoral sul de Pernambuco. Pesqui. Geociências 2003, 30, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Gruber, S.H.; Stout, R.G. Biological materials for the study of age and growth in a tropical marine elasmobranch the lemon shark. In Proceedings International Workshop on Age Determination of Oceanic Pelagic Fishes: Tunas, Billfishes, Sharks; Prince, E.D., Pulos, L.M., Eds.; NOAA Technical Report NMFS: Seattle, DC, USA, 1983; Volume 8, pp. 193–205. [Google Scholar]
- Casey, J.G.; Pratt, H.L., Jr.; Stillwell, C. Age and growth of the sandbar shark, Carcharhinus plumbeus, from the western North Atlantic. In Proceedings International Workshop on Age Determination of Oceanic Pelagic Fishes: Tunas, Billfishes, Sharks, Prince, E.D.; Pulos, L.M., Ed.; NOAA Technical Report NMFS: Seattle, DC, USA, 1983; Volume 8, pp. 189–191. [Google Scholar]
- Cailliet, G.M.; Smith, W.D.; Mollet, H.F.; Goldman, J. Age and growth studies of chondrichthyan fishes: The need for consistency in terminology, verification, validation, and growth function fitting. Environ. Biol. Fish. 2006, 77, 211–228. [Google Scholar] [CrossRef]
- Cailliet, G.M.; Martin, L.K.; Harvey, J.T.; Kusher, D.; Welden, B.A. Preliminary studies on the age and growth of blue (Prionace glauca), common thresher (Alopias vulpinus), and shortfin mako (Isurus oxyrinchus) sharks from California waters. In Proceedings International Workshop on Age Determination of Oceanic Pelagic Fishes: Tunas, Billfishes, Sharks, Prince, E.D.; Pulos, L.M., Ed.; NOAA Technical Report NMFS: Seattle, DC, USA, 1983; Volume 8, pp. 179–188. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice-Hall: Upper Saddle River, NJ, USA, 1996; 944p. [Google Scholar]
- Goldman, K.J. Aspects of Age, Growth, Demographics and Thermal Biology of Two Lamniform Shark Species. Ph.D. Thesis, College of William and Mary, Virginia Institute of Marine Science, Williamsburg, VA, USA, 2002. [Google Scholar]
- Beamish, R.J.; Fournier, D.A. A method for comparing the precision of a set of age determinations. Can. J. Fish. Aquat. Sci. 1981, 38, 982–983. [Google Scholar] [CrossRef]
- Natanson, L.J.; Casey, J.G.; Kohler, N.E. Age and growth estimates for the dusky shark, Carcharhinus obscurus, in the western North Atlantic Ocean. Fish. Bull. 1995, 93, 116–126. [Google Scholar]
- Okamura, H.; Semba, Y. A novel statistical method for validating the periodicity of vertebral growth band formation in elasmobranch fishes. Can. J. Fish. Aquat. Sci. 2009, 66, 771–780. [Google Scholar] [CrossRef]
- Akaike, H. Information theory as an extension of the maximum likelihood principle. In Second International Symposium on Information Theory; Petrov, B.N., Csaki, F., Eds.; Akademiai Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Beverton, R.J.H. Notes on the Use of Theoretical Models in the Study of the Dynamics of Exploited Fish Populations; Miscellaneous Contributions; United States Fishery Laboratory: Beaufort, NC, USA, 1954; 159p. [Google Scholar]
- Fabens, A.J. Properties and fitting of the von Bertalanffy growth curve. Growth 1965, 29, 265–289. [Google Scholar]
- Simpfendorfer, C.A. Growth rates of juvenile dusky sharks, Carcharhinus obscurus (Lesueur, 1818), from southwestern Australia estimated from tag-recapture data. Fish. Bull. 2000, 98, 811–822. [Google Scholar]
- Araya, M.; Cubillos, L.A. Evidence of two phase growth in elasmobranchs. Environ. Biol. Fish. 2006, 77, 293–300. [Google Scholar] [CrossRef]
- Campana, S.E.; Jones, C.M. Analysis of otolith microstructure data. pp. 73–100. In Otolith Microstructure Examination and Analysis; Stevenson, D.K., Campana, S.E., Eds.; Canadian Special Publication of Fisheries and Aquatic Sciences; Fisheries and Oceans Canada: Ottawa, ON, USA, 1992; 117p. [Google Scholar]
- Schnute, J.A. versatile growth model with statistically stable parameters. Can. J. Fish. Aquat. Sci. 1981, 38, 1128–1140. [Google Scholar] [CrossRef]
- Harry, A.V.; Simpfendorfer, C.A.; Tobin, A.J. Improving age, growth, and maturity estimates for aseasonally reproducing chondrichthyans. Fish. Res. 2010, 106, 393–403. [Google Scholar] [CrossRef]
- Hood, G.M. PopTools Version 3.2.5. 2010. Available online: http://www.poptools.org (accessed on 10 June 2014).
- Kimura, D.K. Likelihood methods for the von Bertalanffy growth curve. Fish. Bull. 1980, 77, 765–776. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; 488p. [Google Scholar]
- Sparre, P.E.; Venema, S.C. Introduction to Tropical Fish Stock; Part 1. Manual. FAO Fisheries Technical Paper 306; FAO: Rome, Italy, 1997; p. 337. [Google Scholar]
- Lovejoy, N. Systematics of myliobatoid elasmobranchs: With emphasis on the phylogeny and historical biogeography of neotropical freshwater stingrays (Potamotrygonidae: Rajiformes). Zool. J. Linn. Soc. 1996, 117, 207–257. [Google Scholar] [CrossRef]
- Thorson, J.T.; Simpfendorfer, C.A. Gear selectivity and sample size effects on growth curve selection in shark age and growth studies. Fish. Res. 2009, 98, 75–84. [Google Scholar] [CrossRef]
- Brothers, E.B. Sumary of Round Table Discussions on Age Validation; NOAA Technical Report NMFS: Seattle, DC, USA, 1983; Volume 8, pp. 35–44. [Google Scholar]
- Beamish, R.J.; McFarlane, G.A. The forgotten requirement for validation in fisheries biology. Trans. Am. Fish. Soc. 1983, 112, 735–743. [Google Scholar] [CrossRef]
- Cailliet, G.M. Perspectives on elasmobranch life–history studies: A focus on age validation and relevance to fishery management. J. Fish Biol. 2015, 87, 1271–1292. [Google Scholar] [CrossRef] [PubMed]
- Lessa, R.; Santana, F.M.; Hazin, F.H. Age and growth of the blue shark Prionace glauca (Linnaeus, 1758) off northeastern Brazil. Fish. Res. 2004, 66, 19–30. [Google Scholar] [CrossRef]
- Santander-Neto, J.; Barreto, R.; Santana, F.M.; Lessa, R.P. Age, growth and demography of the silky shark Carcharhinus falciformis from the southwestern Atlantic. Endanger Species Res. 2021, 45, 237–249. [Google Scholar] [CrossRef]
- Wourms, J.P. Reproduction and development in chondrichthyan fishes. Am. Zool. 1977, 17, 379–410. [Google Scholar] [CrossRef]
- Santana, F.M.; Lessa, R. Age determination and growth of the night shark (Carcharhinus signatus) off the northeastern Brazilian coast. Fish. Bull. 2004, 102, 156–167. [Google Scholar]
- Katsanevakis, S. Modelling fish growth: Model selection, multi-model inference and model selection uncertainty. Fish. Res. 2006, 81, 229–235. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Maravelias, C.D. Modelling fish growth: Multi-model inference as a better alternative to a priori using von Bertalanffy equation. Fish Fish. 2008, 9, 178–187. [Google Scholar] [CrossRef]
- Barreto, R.R.; Lessa, R.P.; Hazin, F.H.; Santana, F.M. Age and growth of the blacknose shark, Carcharhinus acronotus (Poey, 1860) off the northeastern Brazilian Coast. Fish. Res. 2011, 110, 170–176. [Google Scholar] [CrossRef]
- Pardo, S.A.; Cooper, A.B.; Dulvy, N.K. Avoiding fishy growth curves. Methods Ecol. Evol. 2013, 4, 353–360. [Google Scholar] [CrossRef]
- Babel, J.S. Reproduction, life history and ecology of the round stingray Urolophus halleri Cooper. Fish. Bull. 1967, 137, 1–104. [Google Scholar]
- Fairbairn, D.J. Allometry for sexual size dimorphism: Pattern and process in the coevolution of body size in males and females. Annu. Rev. Ecol. Syst. 1997, 28, 659–687. [Google Scholar] [CrossRef]
- Blanckenhorn, W.U. Behavioral causes and consequences of sexual size dimorphism. Ethology 2005, 111, 977–1016. [Google Scholar] [CrossRef]
- Aversa, M.I.; Dans, S.L.; García, N.A.; Crespo, E.A. Growth models fitted to Dipturus chilensis length-at-age data support a two phase growth. Rev. Chil. Hist. Nat. 2011, 84, 33–49. [Google Scholar] [CrossRef]
- Klimley, A.P. Diet and growth. In The Biology of Sharks and Rays; University of Chicago Press: Chicago, IL, USA, 2013; pp. 339–372. [Google Scholar]
- Holden, M.J. Problems in the rational exploitation of elasmobranch populations and some suggested solutions. In Sea Fisheries Research; Harden-Jones, F.R., Ed.; John Wiley & Sons: New York, NY, USA, 1974; pp. 117–137. [Google Scholar]
- Campana, S.E. Accuracy, precision and quality control in age determination, incluing a review of the use and abuse of age validation methods. J. Fish. Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Ricker, W.E. Growth rates and models. In Fish Physiology; Hoar, W.S., Randall, D.J., Brett, J.R., Eds.; Bioenergetics and Growth; Academic Press: Cambridge, MA, USA, 1979; Volume VIII, pp. 677–743. [Google Scholar]
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Equation |
|---|---|
| VBGM | |
| VBGM L0 | |
| VBGM TP | |
| Gompertz | |
| Logistic |
| Dataset | Model | L∞ | k | t0 | h | th | AICi | Δi | wi |
|---|---|---|---|---|---|---|---|---|---|
| Female—1 ring | VBGM-L0 | 282.55 (272.63/292.47) | 0.37 (0.32/0.42) | 1575.68 | 0.00 | 0.28 | |||
| Gompertz | 277.03 (268.11/285.95) | 1577.45 | 1.76 | 0.12 | |||||
| VBGM-TP | 286.74 (270.55/302.93) | 0.34 (0.24/0.45) | −1.31 (−2.06/−0.57) | −0.06 (−0.36/0.24) | 0.55 (−2.82/3.92) | 1579.70 | 4.01 | 0.04 | |
| Logistic | 271.70 (264.47/278.94) | 0.62 (0.53/0.70 | 0.47 (0.33/0.61) | 1580.58 | 4.90 | 0.02 | |||
| VBGM | 286.01 (273.79/298.23) | 0.35 (0.28/0.41) | −1.41 (−1.70/−1.13) | 1582.61 | 6.93 | 0.01 | |||
| Female—2 rings | VBGM-L0 | 304.59 (287.78/321.39) | 0.50 (0.41/0.58) | 1575.68 | 0.00 | 0.28 | |||
| Logistic | 276.62 (268.23/285.04) | 1.07 (0.93/1.22) | 0.43 (0.34/0.51) | 1577.17 | 1.48 | 0.13 | |||
| Gompertz | 285.54 (274.10/296.60) | 1578.31 | 2.63 | 0.08 | |||||
| VBGM-TP | 294.08 (254.41/333.73) | 0.50 (0.01/1.00) | −1.42 (−3.99/1.14) | 0.37 (−0.20/0.94) | 0.18 (−0.50/0.86) | 1579.93 | 4.24 | 0.03 | |
| VBGM | 302.45 (284.42/320.47) | 0.51 (0.41/0.62) | −0.80 (−0.98/−0.62) | 1582.61 | 6.93 | 0.01 | |||
| Male—1 ring | VBGM-TP | 230.35 (220.48/240.21) | 1.00 (0.56/1.44) | −0.76 (−1.03/−0.48) | 0.34 (0.11/0.56) | 1.16 (0.86/1.47) | 1343.66 | 0.00 | 0.33 |
| Logistic | 237.33 (228.59/246.06) | 0.95 (0.79/1.11) | 0.14 (0.02/0.26) | 1345.15 | 1.49 | 0.15 | |||
| Gompertz | 241.87 (231.05/252.68) | 1346.60 | 2.94 | 0.07 | |||||
| VBGM-L0 | 247.42 (234.85/259.99) | 0.57 (0.46/0.68) | 1347.25 | 3.59 | 0.05 | ||||
| VBGM | 249.18 (234.54/263.82) | 0.55 (0.41/0.68) | −1.04 (−1.30/−0.77) | 1348.81 | 5.15 | 0.02 | |||
| Male—2 rings | VBGM-TP | 288.85 (220.33/237.38) | 2.33 (0.95/3.70) | −0.47 (−0.69/−0.24) | 0.59 (0.42/0.76) | 0.68 (0.52/0.84) | 1343.78 | 0.12 | 0.31 |
| VBGM-L0 | 278.19 (249.98/306.40) | 0.63 (0.46/0.81) | 1347.25 | 3.59 | 0.05 | ||||
| Logistic | 246.82 (234.42/259.22) | 1.44 (1.18/1.70) | 0.22 (0.14/0.31) | 1352.19 | 8.54 | 0.00 | |||
| Gompertz | 256.86 (239.42/274.31) | 1356.78 | 13.12 | 0.00 | |||||
| VBGM | 275.87 (246.37/305.36) | 0.65 (0.44/0.87) | −0.71 (−0.91/−0.51) | 1362.23 | 18.58 | 0.00 |
| Species | Model | Sex | DW∞ | k | tmax | DWmax | tmat | n | Study Area | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Urotrygon microphthalmum | VBGM L0 | F | 147 * | 0.37 | 8.5 | 154 | 2 | 186 | SW Atlantic, Brazil | Present study |
| VBGM TP | M | 127 * | 1.00 | 5.5 | 128 | 1.5 | 161 | |||
| Urotrygon aspidura | VBGM TP | F | 249 | 0.47 | 7.5 | 265 | 2.3 | 125 | E Pacific, Colômbia | [4] |
| VBGM TP | M | 160 | 1.63 | 5.5 | 185 | - | 184 | |||
| Urotrygon rogersi | VBGM TP | F | 200 | 0.22 | 8 | 199 | 1 | 234 | E Pacific, Colômbia | [26] |
| VBGM TP | M | 155 | 0.64 | 6 | 170 | 1 | 232 | |||
| Urobatis halleri | VBGM | F | 224 | 0.15 | 14 | 213 ** | 3.8 | 96 | NE Pacific, USA | [25] |
| VBGM | M | 286 | 0.09 | 14 | 239 ** | 3.8 | 84 | |||
| Urolophus lobatus | VBGM | F | 249 | 0.37 | 15 | 277 | 3.1 | 388 | SE Indian, Australia | [22] |
| VBGM | M | 210 | 0.51 | 13 | 237 | 1.7 | 428 | |||
| Urolophus paucimaculatus | VBGM | F | 261 | 0.26 | 14 | 272 | 5 | 330 | SE Indian, Australia | [24] |
| VBGM | M | 243 | 0.36 | 11 | 256 | 3.5 | 437 | |||
| Trygonoptera personata | VBGM | F | 303 | 0.14 | 16 | 311 | 4 | 352 | SE Indian, Australia | [23] |
| VBGM | M | 269 | 0.20 | 10 | 269 | 4 | 303 | |||
| Trygonoptera mucosa | VBGM | F | 308 | 0.24 | 17 | 369 | 5 | 324 | SE Indian, Australia | [23] |
| VBGM | M | 261 | 0.49 | 12 | 283 | 2 | 400 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santander-Neto, J.; Santana, F.M.; Vasconcelos-Filho, J.E.; Lessa, R. Age and Growth of the Threatened Smalleye Round Ray, Urotrygon microphthalmum, Delsman, 1941, from Northeastern Brazil. Fishes 2023, 8, 160. https://doi.org/10.3390/fishes8030160
Santander-Neto J, Santana FM, Vasconcelos-Filho JE, Lessa R. Age and Growth of the Threatened Smalleye Round Ray, Urotrygon microphthalmum, Delsman, 1941, from Northeastern Brazil. Fishes. 2023; 8(3):160. https://doi.org/10.3390/fishes8030160
Chicago/Turabian StyleSantander-Neto, Jones, Francisco Marcante Santana, Jonas Eloi Vasconcelos-Filho, and Rosângela Lessa. 2023. "Age and Growth of the Threatened Smalleye Round Ray, Urotrygon microphthalmum, Delsman, 1941, from Northeastern Brazil" Fishes 8, no. 3: 160. https://doi.org/10.3390/fishes8030160