
Zebrafish Sexual Behavior in Plain and Enriched Environments: Parameters in the Valproate Model of Autism

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Embryo Collection
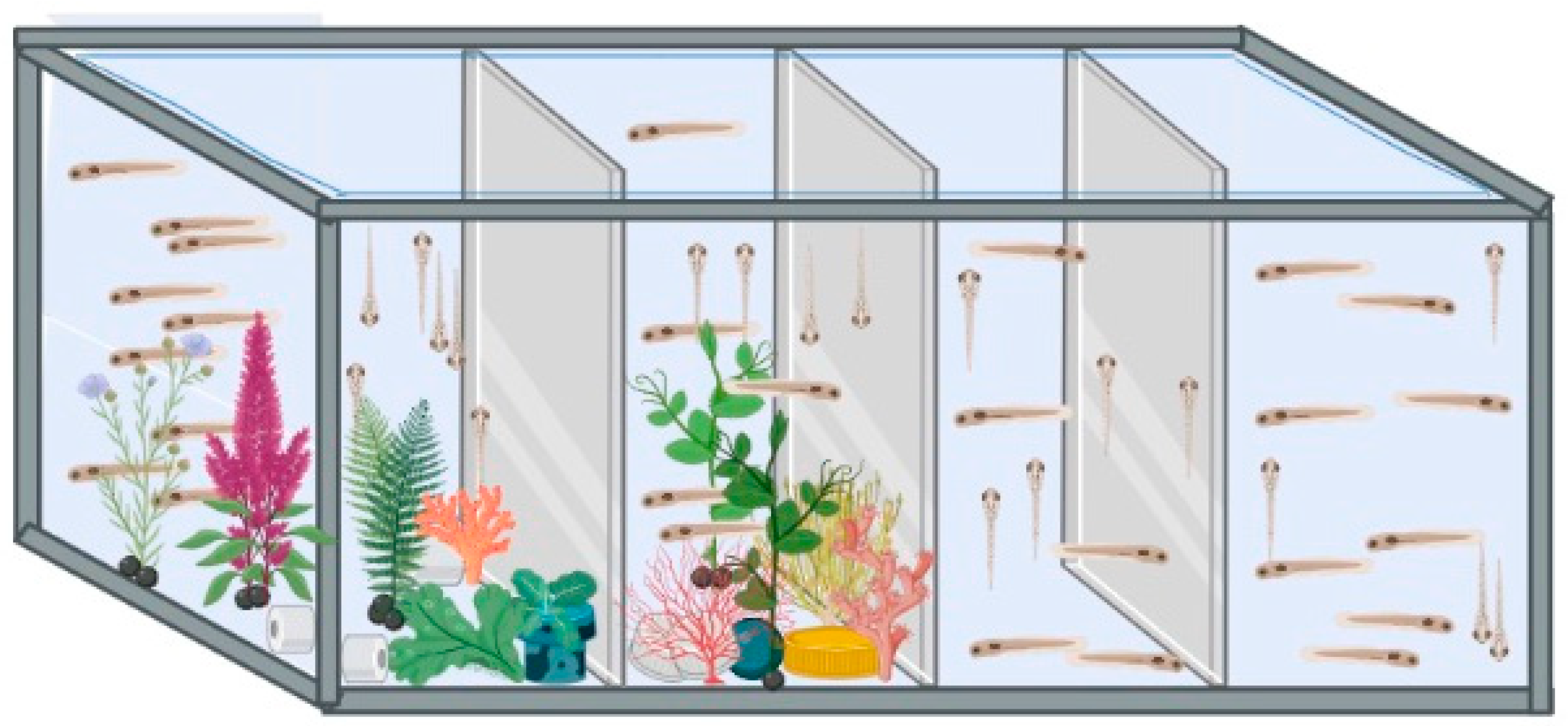
2.3. Larval Rearing and Environment Conditions
2.4. Sexual Behavior Recording
2.5. Statistics
3. Results
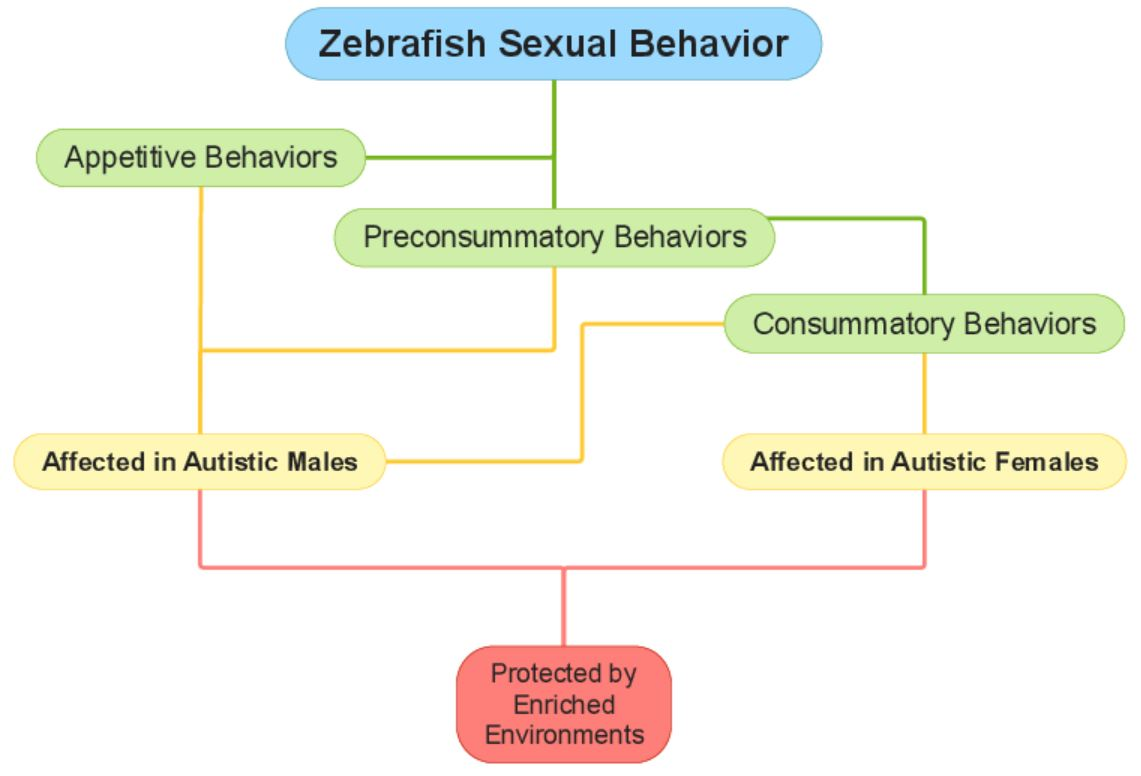
3.1. Sexual Behavior
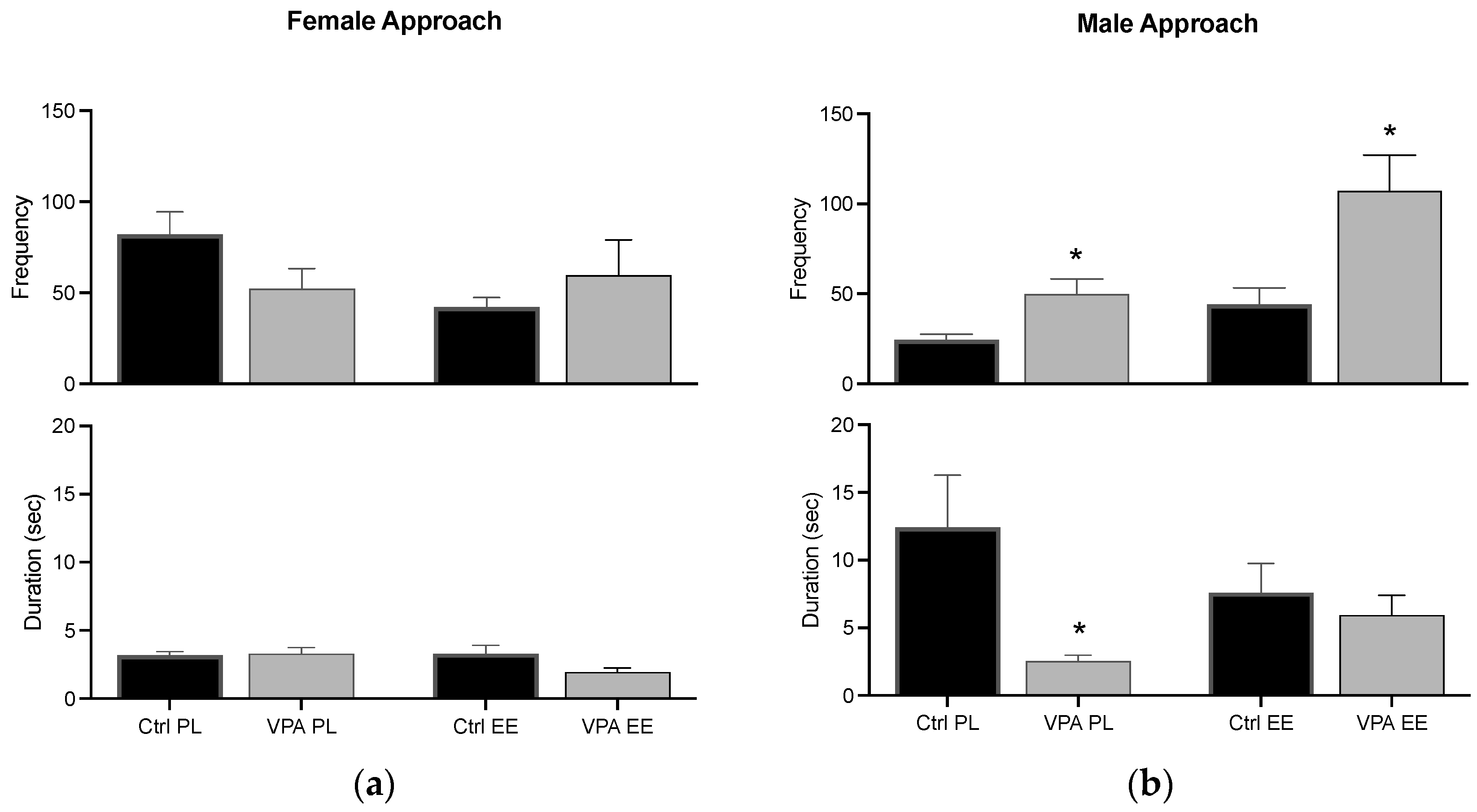
3.2. Appetitive Behavior
Approach
3.3. Preconsummatory Behaviors
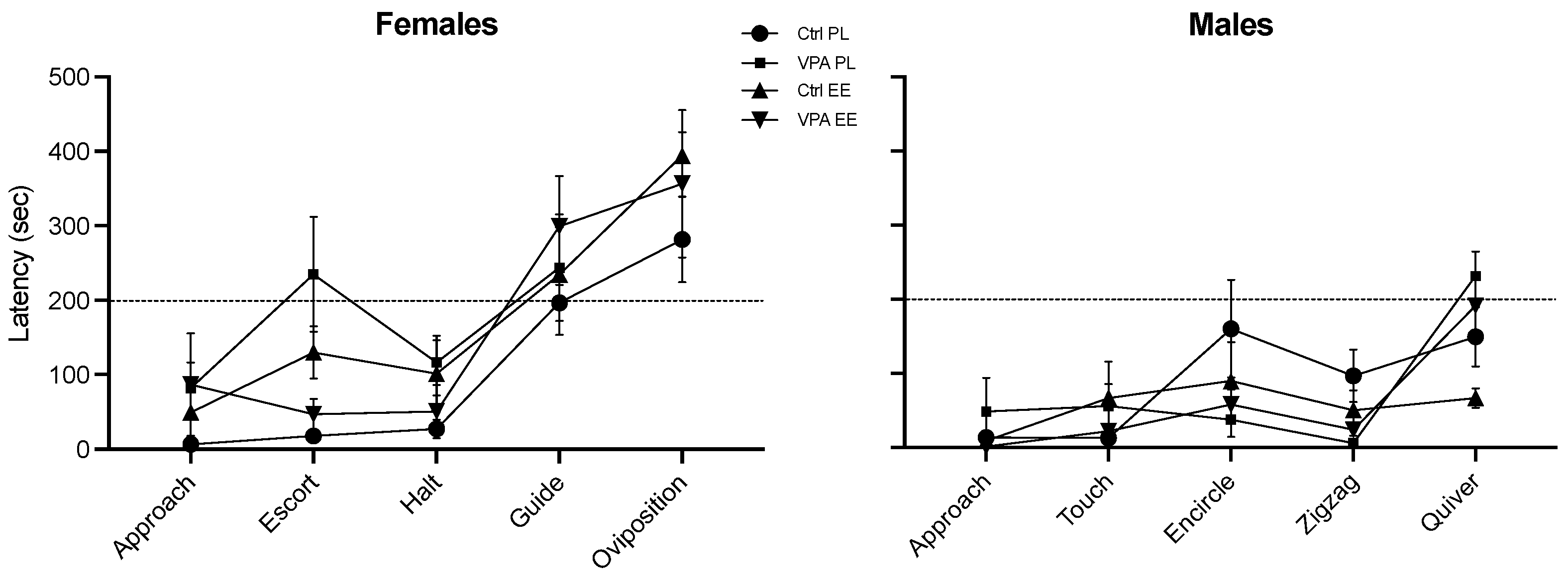
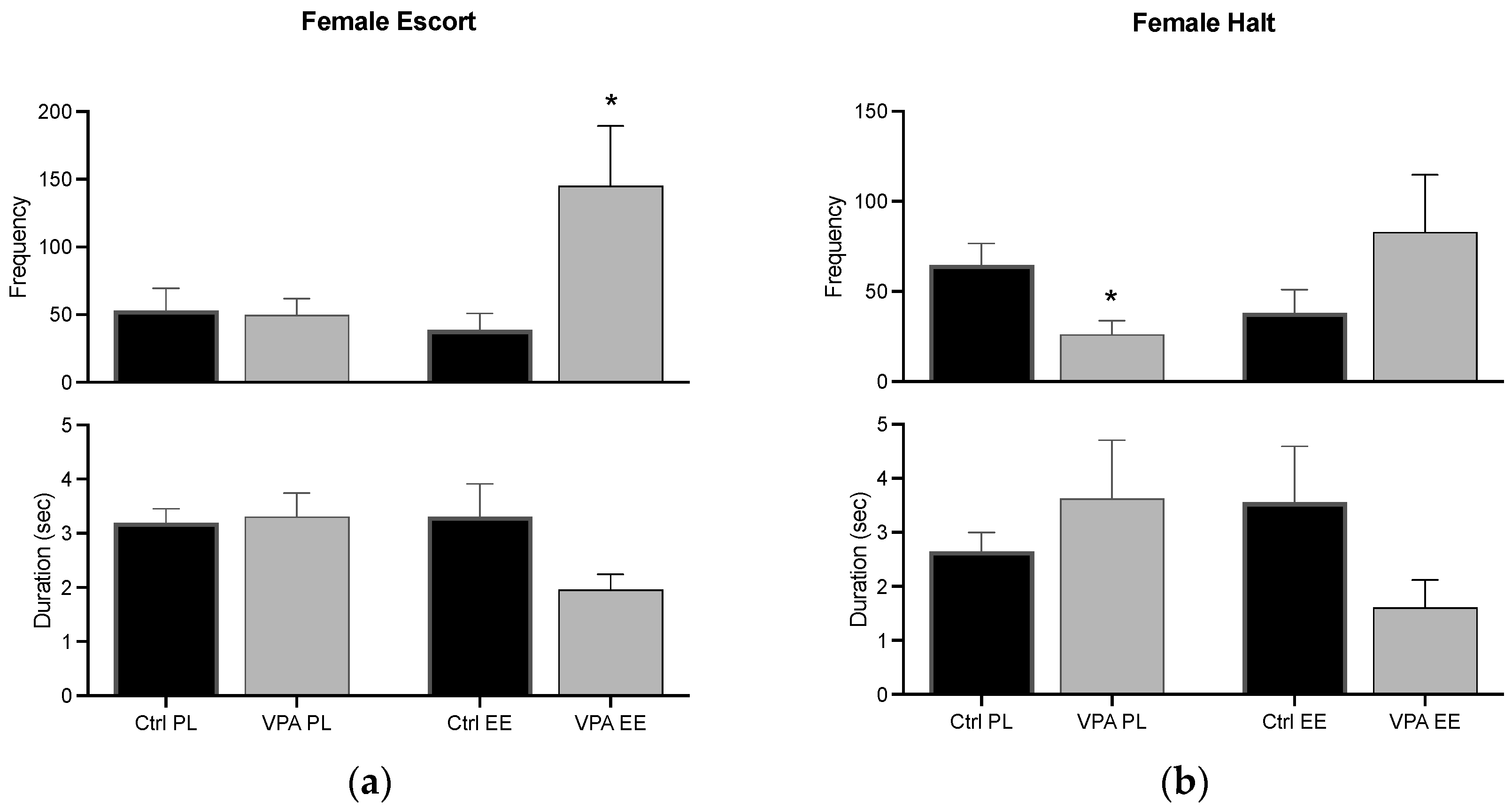
3.3.1. Female Escort and Halt
3.3.2. Male Touch and Encircle
3.4. Consummatory Behaviors
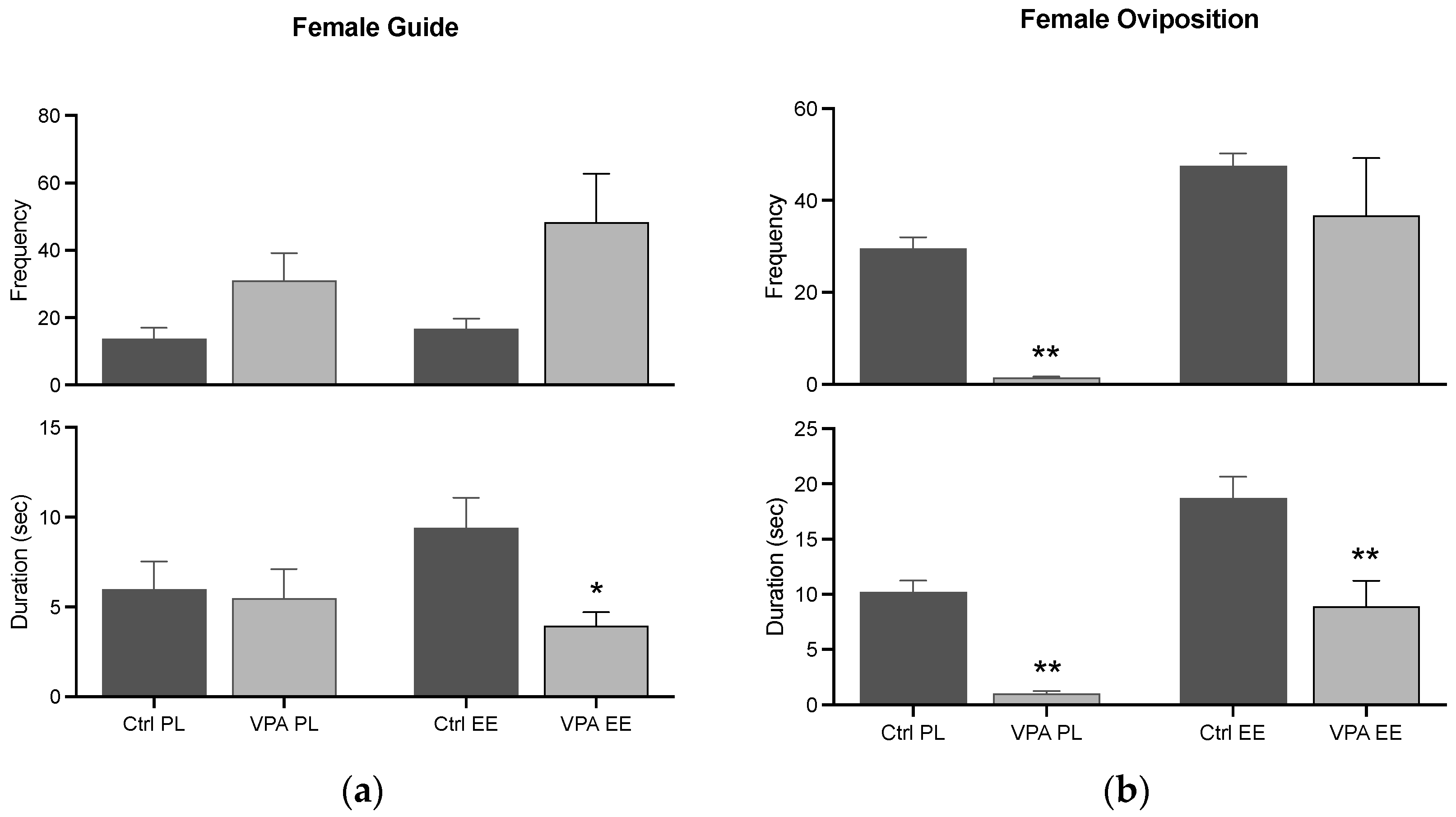
3.4.1. Female Guide and Oviposition
3.4.2. Male Zigzag and Quiver
4. Discussion
5. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Manzo, J. Un Segundo Espectro Del Autismo: De La Conducta a La Neurona. eNeurobiol 2019, 23, 1501. [Google Scholar] [CrossRef]
- Courchesne, E.; Carper, R.; Akshoomoff, N. Evidence of Brain Overgrowth in the First Year of Life in Autism. JAMA 2003, 290, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Lechuga, A.I.; Nuñez-Arcos, L.Y.; Carrillo, P.; García, L.I.; Coria-Ávila, G.A.; Toledo, R.; Hernández, M.E.; Manzo, J. Reduction of Cutaneous von Frey Thresholds in Boys with Autism Following a Year of Tactile and Emotional Stimulation. Rev. Mex. Neuroc. 2021, 22, 85–88. [Google Scholar] [CrossRef]
- Kanner, L. Autistic Disturbances of Affective Contact. Nervous Child 1943, 2, 217–250. [Google Scholar]
- Byers, E.S.; Nichols, S.; Voyer, S.D. Challenging Stereotypes: Sexual Functioning of Single Adults with High Functioning Autism Spectrum Disorder. J. Autism Dev. Disord. 2013, 43, 2617–2627. [Google Scholar] [CrossRef]
- Kellaher, D.C. Sexual Behavior and Autism Spectrum Disorders: An Update and Discussion. Curr. Psychiat. Rep. 2015, 17, 562. [Google Scholar] [CrossRef] [PubMed]
- Dewinter, J.; Vermeiren, R.; Vanwesenbeeck, I.; Lobbestael, J.; Nieuwenhuizen, C.V. Sexuality in Adolescent Boys with Autism Spectrum Disorder: Self-Reported Behaviours and Attitudes. J. Autism Dev. Disord. 2015, 45, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Brown-Lavoie, S.M.; Viecili, M.A.; Weiss, J.A. Sexual Knowledge and Victimization in Adults with Autism Spectrum Disorders. J. Autism Dev. Disord. 2014, 44, 2185–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ågmo, A. Male Rat Sexual Behavior. Brain Res. Protoc. 2019, 1, 203–209. [Google Scholar] [CrossRef]
- Meisel, R.L.; Sachs, B.D. The Physiology of Male Sexual Behavior. In The Physiology of Reproduction, 2nd ed.; Knobil, E., Neill, J.D., Eds.; Raven Press: New York, NY, USA, 1994; Volume 2, pp. 3–105. [Google Scholar]
- Manzo, J.; Carrillo, P.; Coria-Avila, G.A.; Garcia, L.I. The Sexual Cerebellum. In Behavioral Neuroendocrinology; Komisaruk, B.R., González-Mariscal, G., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 103–112. [Google Scholar]
- Manzo, J.; Coria-Avila, G.A.; Garcia, L.I.; Hernández-Aguilar, M.E.; Herrera-Covarrubias, D.; Toledo, R.; Monje-Reyna, D.; Santamaría, F. Male Sexual Behavior and Prostate Histology in a Rat Model of Autism. Eneurobiología 2019, 25, 280919. [Google Scholar] [CrossRef]
- Chen, J.; Lei, L.; Tian, L.; Hou, F.; Roper, C.; Ge, X.; Zhao, Y.; Chen, Y.; Dong, Q.; Tanguay, R.L.; et al. Developmental and Behavioral Alterations in Zebrafish Embryonically Exposed to Valproic Acid (VPA): An Aquatic Model for Autism. Neurotoxicol. Teratol. 2018, 66, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Darrow, K.O.; Harris, W.A. Characterization and Development of Courtship in Zebrafish, Danio rerio. Zebrafish 2004, 1, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Bloom, H.D.; Perlmutter, A. A Sexual Aggregating Pheromone System in the Zebrafish, Brachydanio rerio (Hamilton-buchanan). J. Exp. Zool. 1977, 199, 215–226. [Google Scholar] [CrossRef]
- Gerlach, G. Pheromonal Regulation of Reproductive Success in Female Zebrafish: Female Suppression and Male Enhancement. Anim. Behav. 2006, 72, 1119–1124. [Google Scholar] [CrossRef]
- Spence, R.; Gerlach, G.; Lawrence, C.; Smith, C. The Behaviour and Ecology of the Zebrafish, Danio rerio. Biol. Rev. 2008, 83, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Hutter; Magee; Zala; Penn Reproductive Behaviour of Wild Zebrafish (Danio rerio) in Large Tanks. Behaviour 2010, 147, 641–660. [CrossRef]
- Ghoshal, A.; Daniel, D.K.; Bhat, A. Temporal Patterns and Sex Differences in Dyadic Interactions in a Wild Zebrafish Population. Behav. Process 2019, 166, 103896. [Google Scholar] [CrossRef] [PubMed]
- DePasquale, C.; Neuberger, T.; Hirrlinger, A.M.; Braithwaite, V.A. The Influence of Complex and Threatening Environments in Early Life on Brain Size and Behaviour. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152564. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, F.F.; Gaspary, K.V.; Leite, C.E.; Cognato, G.D.P.; Bonan, C.D. Embryological Exposure to Valproic Acid Induces Social Interaction Deficits in Zebrafish (Danio rerio): A Developmental Behavior Analysis. Neurotoxicol. Teratol. 2015, 52, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [Green Version]
- Pfaus, J.G.; Kippin, T.E.; Coria-Avila, G. What Can Animal Models Tell Us about Human Sexual Response? Annu. Rev. Sex Res. 2003, 14, 1–63. [Google Scholar] [PubMed]
- Dwivedi, S.; Medishetti, R.; Rani, R.; Sevilimedu, A.; Kulkarni, P.; Yogeeswari, P. Larval Zebrafish Model for Studying the Effects of Valproic Acid on Neurodevelopment: An Approach towards Modeling Autism. J. Pharmacol. Toxicol. 2019, 95, 56–65. [Google Scholar] [CrossRef]
- Korsching, S.I.; Argo, S.; Campenhausen, H.; Friedrich, R.W.; Rummrich, A.; Weth, F. Olfaction in Zebrafish: What Does a Tiny Teleost Tell Us? Semin. Cell Dev. Biol. 1997, 8, 181–187. [Google Scholar] [CrossRef]
- Santacà, M.; Dadda, M.; Bisazza, A. The Role of Visual and Olfactory Cues in Social Decisions of Guppies and Zebrafish. Anim. Behav. 2021, 180, 209–217. [Google Scholar] [CrossRef]
- Pfaus, J.G. Neurobiology of Sexual Behavior. Curr. Opinion. Neurobiol. 1999, 9, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Paredes, R.G.; Vazquez, B. What Do Female Rats like about Sex? Paced Mating. Behav. Brain Res. 1999, 105, 117–127. [Google Scholar] [CrossRef]
- Welberg, L. Autism: The Importance of Getting the Dose Right. Nat. Rev. Neurosci. 2011, 12, 429. [Google Scholar] [CrossRef]
- Grissom, N.M.; McKee, S.E.; Schoch, H.; Bowman, N.; Havekes, R.; O’Brien, W.T.; Mahrt, E.; Siegel, S.; Commons, K.; Portfors, C.; et al. Male-Specific Deficits in Natural Reward Learning in a Mouse Model of Neurodevelopmental Disorders. Mol. Psychiatr. 2018, 23, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, E.J.; Almada, V.C.; Oliveira, R.F.; Santos, A.J. Female Mimicry as a Mating Tactic in Males of the Blenniid Fish Salaria Pavo. J. Mar. Biol. Assoc. UK 1996, 76, 529–538. [Google Scholar] [CrossRef]
- Gonçalves, D.; Alpedrinha, J.; Teles, M.; Oliveira, R.F. Endocrine Control of Sexual Behavior in Sneaker Males of the Peacock Blenny Salaria Pavo: Effects of Castration, Aromatase Inhibition, Testosterone and Estradiol. Horm. Behav. 2007, 51, 534–541. [Google Scholar] [CrossRef]
- Todd, E.V.; Liu, H.; Lamm, M.S.; Thomas, J.T.; Rutherford, K.; Thompson, K.C.; Godwin, J.R.; Gemmell, N.J. Female Mimicry by Sneaker Males Has a Transcriptomic Signature in Both the Brain and the Gonad in a Sex-Changing Fish. Mol. Biol. Evol. 2017, 35, 225–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Guo, J.-Y.; Li, X.; Zhou, H.-J.; Zhang, S.-H.; Liu, X.-D.; Chen, D.-Y.; Fang, Y.-C.; Feng, X.-Z. Behavioural Effect of Low-Dose BPA on Male Zebrafish: Tuning of Male Mating Competition and Female Mating Preference during Courtship Process. Chemosphere 2017, 169, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Fan, Z.; Yang, M.; Zhang, S.; Li, M.; Fang, Y.; Li, J.; Feng, X. Low Concentrations of the Antidepressant Venlafaxine Affect Courtship Behaviour and Alter Serotonin and Dopamine Systems in Zebrafish (Danio rerio). Aquat. Toxicol. 2022, 244, 106082. [Google Scholar] [CrossRef]
- Beach, F.A. Sexual Attractivity, Proceptivity, and Receptivity in Female Mammals. Horm. Behav. 1976, 7, 105–138. [Google Scholar] [CrossRef]
- McClintock, M.K.; Adler, N.T. The Role of the Female During Copulation in Wild and Domestic Norway Rats (Rattus Norvegicus). Behaviour 1978, 67, 67–95. [Google Scholar] [CrossRef]
- Chu, X.; Ågmo, A. Studies of Sociosexual Interactions in Rats in an Externally Valid Procedure: Are They Relevant for Understanding Human Sexual Behavior? Int. J. Psychol. Res. 2016, 9, 76–95. [Google Scholar] [CrossRef] [Green Version]
- Zempo, B.; Tanaka, N.; Daikoku, E.; Ono, F. High-Speed Camera Recordings Uncover Previously Unidentified Elements of Zebrafish Mating Behaviors Integral to Successful Fertilization. Sci. Rep.-UK 2021, 11, 20228. [Google Scholar] [CrossRef] [PubMed]
- Converse, A.; Thomas, P. The Zinc Transporter ZIP9 (Slc39a9) Regulates Zinc Dynamics Essential to Egg Activation in Zebrafish. Sci. Rep.-UK 2020, 10, 15673. [Google Scholar] [CrossRef]
- Ploeger, A.; Galis, F. Evolutionary Approaches to Autism- an Overview and Integration. Mcgill J. Med. 2020, 13, 38. [Google Scholar] [CrossRef]
- Sachs, B.D.; Barfield, R.J. Temporal Patterning of Sexual Behavior in the Male Rat. J. Comp. Physiol. Psych. 1970, 73, 359. [Google Scholar] [CrossRef]
- Manzo, J.; Miquel, M.; Toledo, R.; Mayor-Mar, J.A.; Garcia, L.I.; Aranda-Abreu, G.E.; Caba, M.; Hernandez, M.E. Fos Expression at the Cerebellum Following Non-Contact Arousal and Mating Behavior in Male Rats. Physiol. Behav. 2008, 93, 357–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.E.; Soto-Cid, A.; Aranda-Abreu, G.E.; Díaz, R.; Rojas, F.; Garcia, L.I.; Toledo, R.; Manzo, J. A Study of the Prostate, Androgens and Sexual Activity of Male Rats. Reprod. Biol. Endocrin. 2007, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucio, R.A.; Manzo, J.; Martínez-Gómez, M.; Sachs, B.D.; Pacheco, P. Participation of Pelvic Nerve Branches in Male Rat Copulatory Behavior. Physiol. Behav. 1994, 55, 241–246. [Google Scholar] [CrossRef]
- Manzo, J.; Vazquez, M.I.; Cruz, M.R.; Hernandez, M.E.; Carrillo, P.; Pacheco, P. Fertility Ratio in Male Rats Effects after Denervation of Two Pelvic Floor Muscles. Physiol. Behav. 2000, 68, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, J.; Frombonne, E.; Scorah, J.; Ibrahim, A.; Durkin, M.S.; Saxena, S.; Yusuf, A.; Shih, A.; Elsabbagh, M. Global Prevalence of Autism: A Systematic Review Update. Autism Res. 2022, 15, 778–790. [Google Scholar] [CrossRef]
- Ferri, S.L.; Abel, T.; Brodkin, E.S. Sex Differences in Autism Spectrum Disorder: A Review. Curr. Psych. Rep. 2018, 20, 2–17. [Google Scholar] [CrossRef]
- Carlstead, K.; Shepherdson, D. Effects of Environmental Enrichment on Reproduction. Zoo Biol. 1994, 13, 447–458. [Google Scholar] [CrossRef]
- Wafer, L.N.; Jensen, V.B.; Whitney, J.C.; Gomez, T.H.; Flores, R.; Goodwin, B.S. Effects of Environmental Enrichment on the Fertility and Fecundity of Zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 291–294. [Google Scholar]
- Sale, A.; Berardi, N.; Maffei, L. Enrich of the Environment to Empower the Brain. Trends Neurosci. 2009, 32, 233–239. [Google Scholar] [CrossRef]
- Antón-Galindo, E.; Vecchia, E.D.; Orlandi, J.G.; Castro, G.; Gualda, E.J.; Young, A.M.J.; Guasch-Piqueras, M.; Arenas, C.; Herrera-Ubeda, C.; Garcia-Fernandez, J.; et al. Deficiency of the ywhaz Gene, Involved in Neurodevelopmental Disorders, Alters Brain aAtivity and Behaviour in Zebrafish. Mol. Psych. 2022, 27, 3739–3748. [Google Scholar] [CrossRef]
- Elsen, G.E.; Choi, L.Y.; Prince, V.E.; Ho, R.K. The Autism Susceptibility Gen met Regulates Zebrafish Cerebellar Development and Facial Motor Neuron Migration. Dev. Biol. 2009, 335, 78–92. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pouchoulen, M.; Miquel, M.; Saft, P.; Brug, B.; Toledo, R.; Hernandez, M.E.; Manzo, J. Prenatal Exposure to Sodium Valproate Alters Androgen Receptor Expression in the Developing Cerebellum in a Region and Age Specific Manner in Male and Female Rats. Int. J. Dev. Neurosci. 2016, 53, 46–52. [Google Scholar] [CrossRef]
- Monje-Reyna, D.; Manzo, J.; Santamaria, F. Effects of Environmental Enrichment and Sexual Dimorphism on the Expression of Cerebellar Receptors in C57BL/6 and BTBR + ltpr3tf/J mice. BMC Res. Notes 2022, 15, 175. [Google Scholar] [CrossRef]
- Perez-Pouchoulen, M.; Toledo, R.; Garcia, L.I.; Perez-Estudillo, C.A.; Coria-Avila, G.A.; Hernandez, M.E.; Carrillo, P.; Manzo, J. Androgen Receptors in Purkinje Neurons are Modulated by Systemic Testosterone and Sexual Training in a Region-Specific Manner in the Male Rat. Physiol. Behav. 2016, 156, 191–198. [Google Scholar] [CrossRef]
- Baronio, D.; Puttonen, H.A.; Sundvik, M.; Semenova, S.; Lehtonen, E.; Panula, P. Embryonic Exposure to Valproic Acid Affects the Histaminergic System and the Social Behaviour of Adult Zebrafish (Danio rerio). Br. J. Pharmacol. 2018, 175, 797–809. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.H.; Reed, B.T.; Hawkins, P. Enrichment for Laboratory Zebrafish—A Review of the Evidence and the Challenges. Animals 2021, 11, 698. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Appetitive Behaviors |
|---|
| Female |
| Approach |
| Swimming toward the male |
| Male |
| Approach |
| Swimming toward the female and alongside the female |
| Preconsummatory Behaviors |
| Female |
| Escort |
| Swimming alongside the male and staying still while courted |
| Halt |
| Vertical swimming in front of the male with stay-still periods |
| Male |
| Touch |
| Physical contact with the female |
| Encircle |
| Swimming around the female |
| Consummatory Behaviors |
| Female |
| Guide |
| Recurrent swimming to the same spot in the tank |
| Oviposition |
| Laying eggs into the water |
| Male |
| Zigzag |
| Whipping the tail alongside the female while swimming in sharp turns |
| Quiver |
| Vibration of the tail close to the female side |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velázquez-Landa, X.; Carrillo, P.; Coria-Avila, G.A.; Herrera-Covarrubias, D.; García, L.I.; Toledo-Cárdenas, M.R.; Hernández-Aguilar, M.E.; Manzo, J. Zebrafish Sexual Behavior in Plain and Enriched Environments: Parameters in the Valproate Model of Autism. Fishes 2023, 8, 156. https://doi.org/10.3390/fishes8030156
Velázquez-Landa X, Carrillo P, Coria-Avila GA, Herrera-Covarrubias D, García LI, Toledo-Cárdenas MR, Hernández-Aguilar ME, Manzo J. Zebrafish Sexual Behavior in Plain and Enriched Environments: Parameters in the Valproate Model of Autism. Fishes. 2023; 8(3):156. https://doi.org/10.3390/fishes8030156
Chicago/Turabian StyleVelázquez-Landa, Xiomara, Porfirio Carrillo, Genaro A. Coria-Avila, Deissy Herrera-Covarrubias, Luis I. García, María Rebeca Toledo-Cárdenas, María Elena Hernández-Aguilar, and Jorge Manzo. 2023. "Zebrafish Sexual Behavior in Plain and Enriched Environments: Parameters in the Valproate Model of Autism" Fishes 8, no. 3: 156. https://doi.org/10.3390/fishes8030156