Exposure of Hyalella bonariensis (Crustacea, Amphipoda) to Essential Oils: Effects on Anesthesia and Swimming Activity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Essential Oils and Major Constituents
2.3. Anesthesia Induction and Recovery
2.4. Locomotor Activity
2.5. Statistical Analysis
3. Results
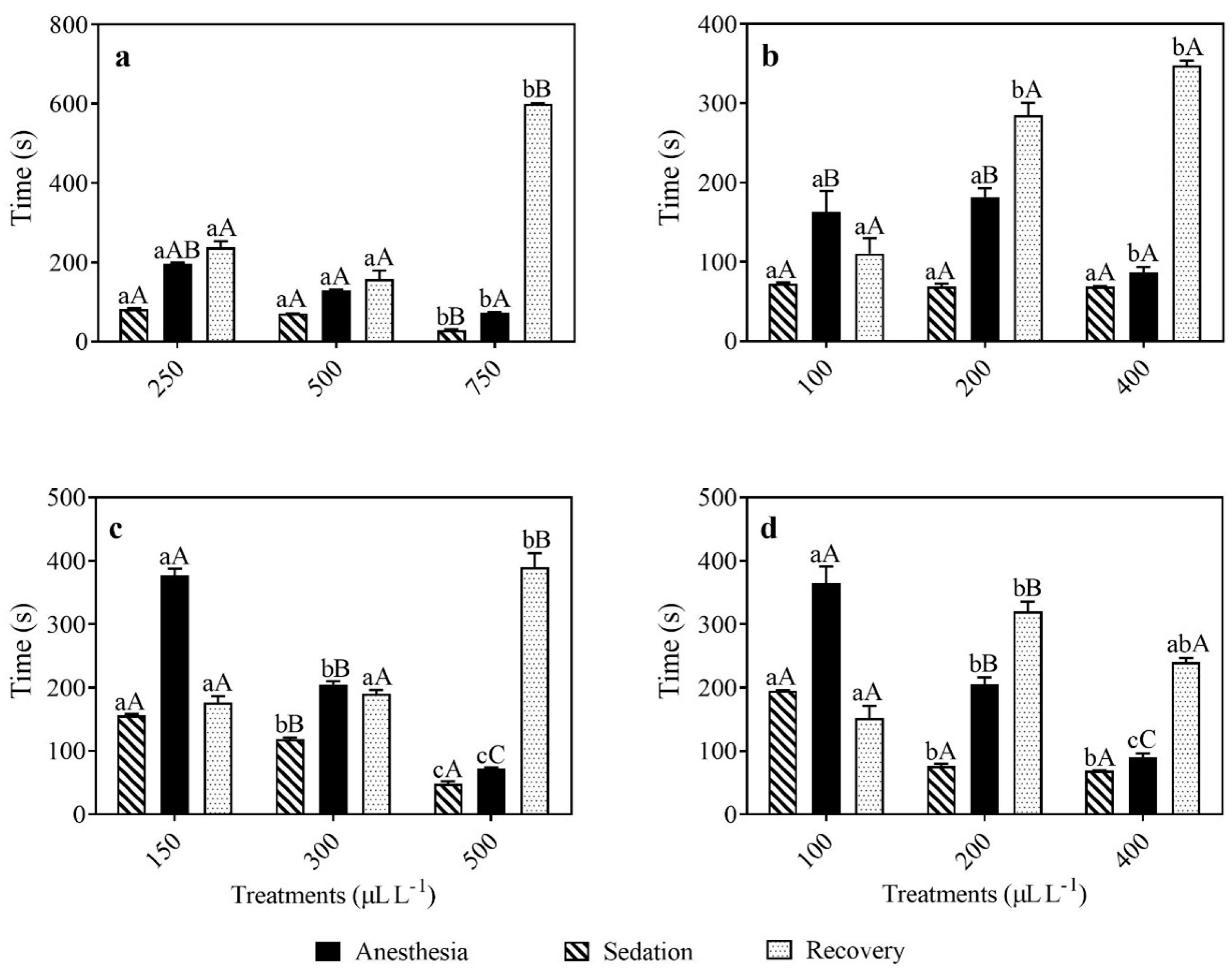
3.1. Anesthesia Induction and Recovery with EOLA and Linalool
3.2. Anesthesia Induction and Recovery with EOAT and Citral
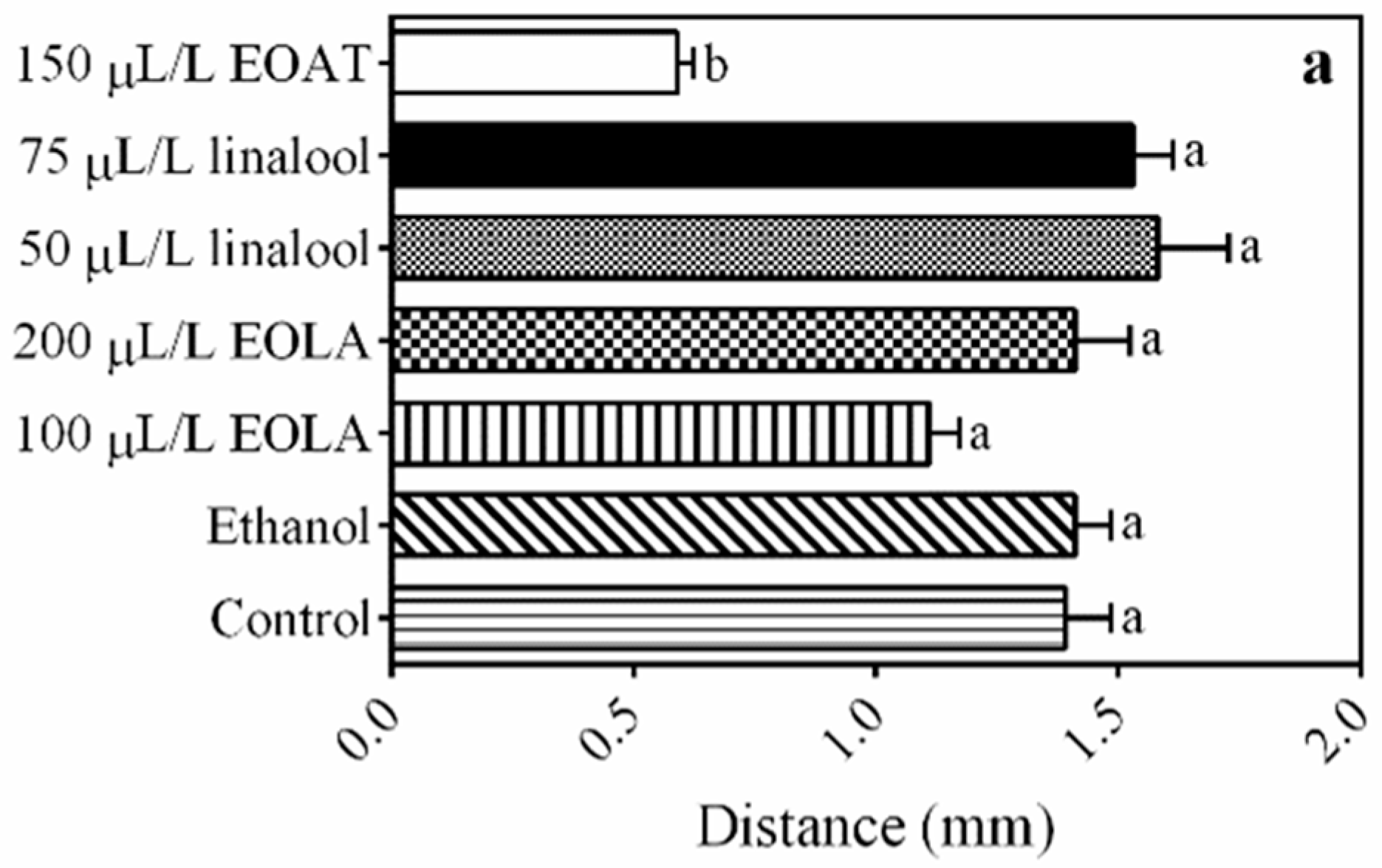
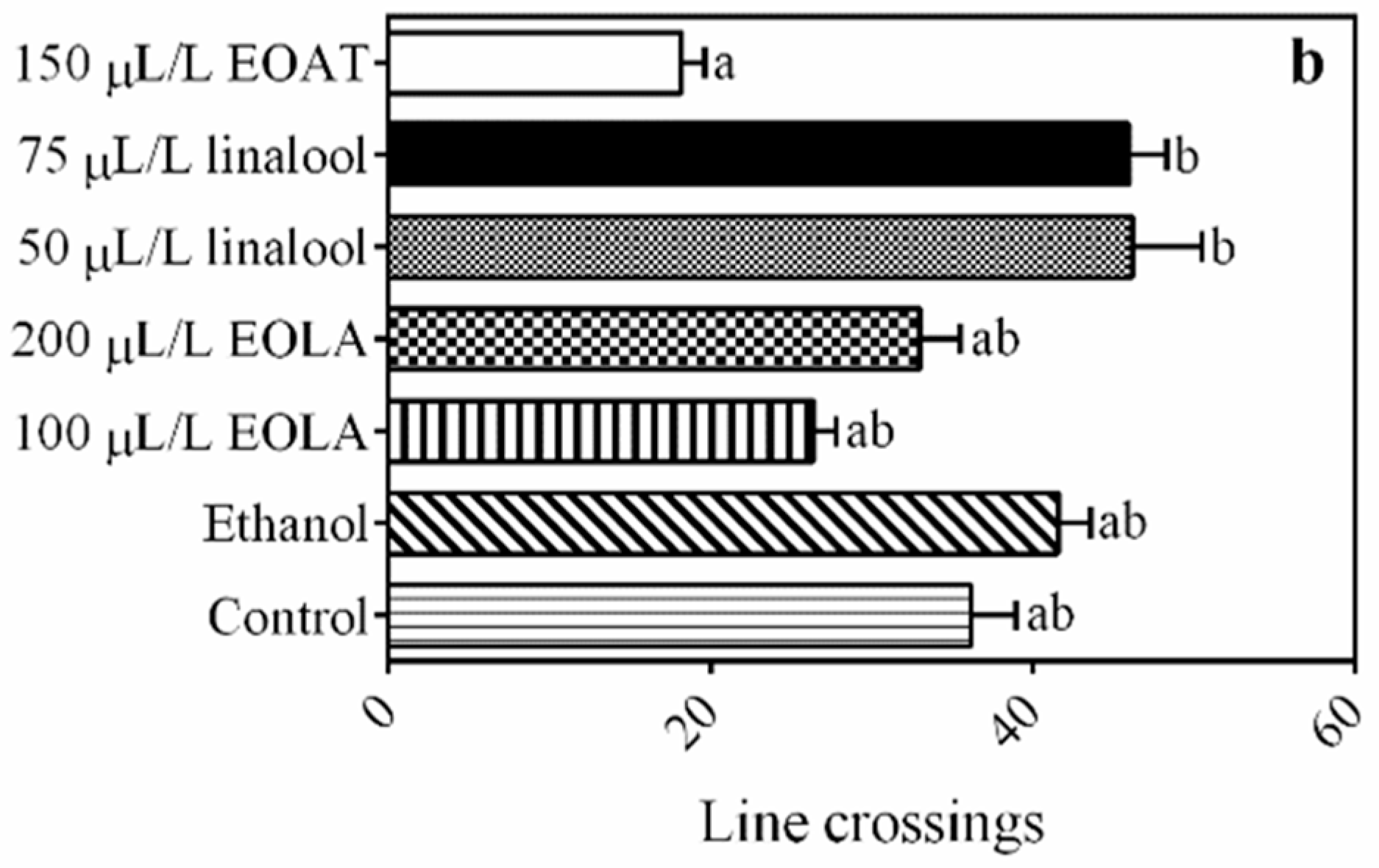
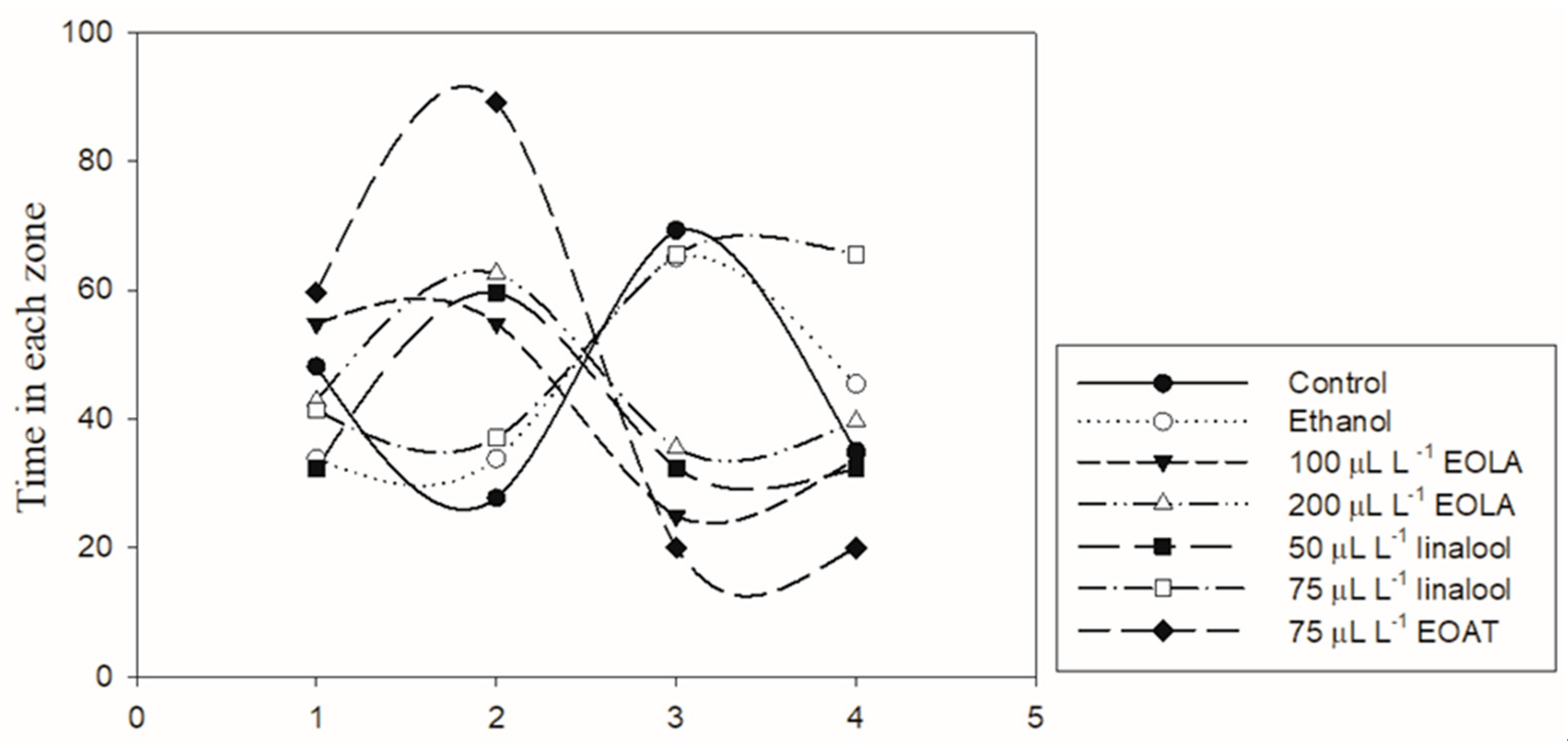
3.3. Locomotor Activity
4. Discussion
4.1. Anesthesia Induction and Recovery
4.2. Locomotor Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Limberger, M.; Santos, S.; Castiglioni, D.S. Hyalella luciae (Crustacea, Amphipoda, Hyalellidae)—A new species of freshwater amphipod from Southern Brazil. Zootaxa 2022, 5174, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Castiglioni, D.D.; Ozga, A.V.; Rodrigues, S.G.; Bueno, A.A.D. Population dynamics of a freshwater amphipod from South America (Crustacea, Amphipoda, Hyalellidae). Nauplius 2016, 24, e2016028. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Guttman, S.I.; Oris, J.T. Genetic differentiation among laboratory populations of Hyalella azteca: Implications for toxicology. Environ. Toxicol. Chem. 1997, 16, 691–695. [Google Scholar] [CrossRef]
- Neuparth, T.; Costa, F.O.; Costa, M.H. Effects of temperature and salinity on life history of the marine amphipod Gammarus locusta. Implications for ecotoxicological testing. Ecotoxicology 2002, 11, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Castiglioni, D.D.S.; Bond-Buckup, G. Reproductive strategies of two sympatric species of Hyalella Smith, 1874 (Amphipoda, Dogielinotidae) in laboratory conditions. J. Nat. Hist. 2007, 41, 1571–1584. [Google Scholar] [CrossRef]
- Ding, Y.; Weston, D.P.; You, J.; Rothert, A.K.; Lydy, M.J. Toxicity of sediment-associated pesticides to Chironomus dilutus and Hyalella azteca. Arch. Environ. Contam. Toxicol. 2011, 61, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Barr, S.; Laming, P.R.; Dick, J.T.A.; Elwood, R.W. Nociception or pain in a decapod crustacean? Anim. Behav. 2008, 75, 745–751. [Google Scholar] [CrossRef]
- Elwood, R.W. Pain and suffering in invertebrates? ILAR J. 2011, 52, 175–184. [Google Scholar] [CrossRef]
- De Lange, H.J.; Peeters, E.T.H.M.; Lürling, M. Changes in ventilation and locomotion of Gammarus pulex (Crustacea, Amphipoda) in response to low concentrations of pharmaceuticals. Hum. Ecol. Risk Assess. 2009, 15, 111–120. [Google Scholar] [CrossRef]
- Bownik, A. Clove essential oil from Eugenia caryophyllus induces anesthesia, alters swimming performance, heart functioning and decreases survival rate during recovery of Daphnia magna. Turk. J. Fish. Aquat. Sci. 2015, 15, 157–166. [Google Scholar] [CrossRef]
- Marks, I.M.; Nesse, R.M. Fear and fitness: An evolutionary analysis of anxiety disorders. Ethol. Sociobiol. 1994, 15, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Fossat, P.; Bacqué-Cazenave, J.; De Deurwaerdère, P.; Delbecque, J.P.; Cattaert, D. Anxiety-like behavior in crayfish is controlled by serotonin. Science 2014, 344, 1293–1297. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Minnot, M.J.; Banchetry, L.; Cézilly, F. Anxiety-like behaviour increases safety from fish predation in an amphipod crustacea. R. Soc. Open Sci. 2017, 4, 171558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacqué-Cazenave, J.; Cattaert, D.; Delbecque, J.P.; Fossat, P. Social harassment induces anxiety-like behaviour in crayfish. Sci. Rep. 2017, 7, 39935. [Google Scholar] [CrossRef] [Green Version]
- Boyd, W.A.; Brewer, S.K.; Williams, P.L. Altered Behaviour of Invertebrates Living in Polluted Environments. In Behavioural Ecotoxicology; Dell’Omo, G., Ed.; John Wiley & Sons: Chichester, UK, 2002; pp. 93–336. [Google Scholar]
- Valente, C.D. Anaesthesia of decapod crustaceans. Vet. Anim. Sci. 2022, 16, 100252. [Google Scholar] [CrossRef] [PubMed]
- Coyle, S.D.; Dasgupta, S.; Tidwell, J.H.; Beavers, T.; Bright, L.A.; Yasharian, D.K. Comparative efficacy of anesthetics for the freshwater prawn Macrobrachium rosenbergii. J. World Aquac. Soc. 2005, 36, 282–290. [Google Scholar] [CrossRef]
- Jensen, M.A.; Fitzgibbon, Q.P.; Carter, C.G.; Adams, L.R. Recovery periods of cultured spiny lobster, Sagmariasus verreauxi juveniles: Effects of handling, force feeding, exercising to exhaustion and anaesthesia on oxygen consumption and ammonia-N excretion rates. Aquaculture 2013, 410, 114–121. [Google Scholar] [CrossRef]
- Guzmán-Sáenz, F.M.; González-Alanís, P.; Martínez, J.G.S.; Salazar, G.G.; Guzmán, G.A.; Perez-Castañeda, R. Uso de diferentes fármacos para anestesiar camarones Litopenaeus vannamei Boone en prácticas de acuacultura. REDVET R. Eléctron. Vet. 2010, 11, 1–9. Available online: http://www.veterinaria.org/revistas/redvet/n030310/031009.pdf (accessed on 10 February 2023).
- Venarsky, M.P.; Vilhelm, F.M. Use of clove oil to anaesthetize freshwater amphipods. Hydrobiologia 2006, 568, 425–432. [Google Scholar] [CrossRef]
- Parodi, T.V.; Cunha, M.A.; Heldwein, C.G.; Souza, D.M.; Martins, A.C.; Garcia, L.O.; Junior, W.W.; Monserrat, J.M.; Schmidt, D.; Caron, B.O.; et al. The anesthetic efficacy of eugenol and the essential oils of Lippia alba and Aloysia triphylla in post-larvae and subadults of Litopenaeus vannamei (Crustacea, Penaeidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Saydmohammed, M.; Pal, A.K. Anesthetic effect of eugenol and menthol on handling stress in Macrobrachium rosenbergii. Aquaculture 2009, 298, 162–167. [Google Scholar] [CrossRef]
- Souza, C.F.; Lima, T.; Baldissera, M.D.; Geihs, M.A.; Maciel, F.E.; Nery, L.E.; Santos, R.C.V.; Raffin, R.P.; Heinzmann, B.M.; Caron, B.O.; et al. Nanoencapsulated Melaleuca alternifolia essential oil exerts anesthetic effects in the brachyuran crab using Neohelice granulata. An. Acad. Bras. Ciências 2018, 90, 2855–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.L.F.; Araujo, P.B.; Bond-Buckup, G. New species and new reports of Hyalella (Crustacea, Amphipoda, Dogielinotidae) from Argentina. Zootaxa 2008, 1760, 24–36. [Google Scholar] [CrossRef]
- Gobbo-Neto, L.; Lopes, N.P. Plantas medicinais: Fatores de influência no conteúdo de metabólitos secundários. Quim. Nova 2007, 30, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Becker, A.J.; Vaz, L.J.; Garcia, L.O.; Wasielesky, W.; Heinzmann, B.M.; Baldisserotto, B. Anesthetic potential of different essential oils for two shrimp species, Farfantepenaeus paulensis and Litopenaeus vannamei (Decapoda, Crustacea). Cienc. Rural 2021, 51, e20200793. [Google Scholar] [CrossRef]
- Li, Y.; Liang, S.; She, Q.; Han, Z.; Li, Y.; Li, X. Influence of temperature and size on menthol anaesthesia in Chinese grass shrimp Palaemonetes sinensis (Sollaud, 1911). Aquac. Res. 2018, 49, 2091–2098. [Google Scholar] [CrossRef]
- Sugawara, Y.; Hara, C.; Tamura, K.; Fujii, T.; Nakamura, K.; Masujima, T.; Aoki, T. Sedative effect on humans of inhalation of essential oil of linalool: Sensory evaluation and physiological measurements using optically active linalools. Anal. Chim. Acta 1998, 365, 293–299. [Google Scholar] [CrossRef]
- Sousa, D.P.; Nóbrega, F.F.; Santos, C.C.M.P.; Almeida, R.N. Anticonvulsant activity of the linalool enantiomers and racemate: Investigation of chiral influence. Nat. Prod. Commun. 2010, 5, 1847–1851. [Google Scholar] [CrossRef] [Green Version]
- Hutt, A.G.; O’Grady, J. Drug chirality: A consideration of the significance of the stereochemistry of antimicrobial agents. J. Antimicrob. Chemother. 1996, 37, 7–32. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Chopra, P. Chirality and anaesthetic drugs: A review and an update. Indian J. Anaesth. 2011, 55, 556–562. [Google Scholar] [CrossRef]
- Silva, L.; Balconi, L.S.; Gressler, L.T.; Garlet, Q.I.; Sutili, F.J.; Vargas, A.P.; Baldisserotto, B.; Morel, A.F.; Heinzmann, B.M. S-(+)-and R-(−)-linalool: A comparison of the in vitro anti-Aeromonas hydrophila activity and anesthetic properties in fish. An. Acad. Bras. Ciênc. 2017, 89, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, A.J.; Ramos, P.B.; Monserrat, J.M.; Wasielesky, W.; Baldisserotto, B. Behavioural and biochemical responses in adult Pacific white shrimp, Litopenaeus vannamei, exposed to the essential oil of Cymbopogon citratus. Aquac. Res. 2021, 52, 6205–6217. [Google Scholar] [CrossRef]
- Heldwein, C.G.; Silva, L.L.; Gai, E.Z.; Roman, C.; Parodi, T.V.; Bürger, M.E.; Baldisserotto, B.; Flores, E.M.M.; Heinzmann, B.M. S-(+)-Linalool from Lippia alba: Sedative and anesthetic for silver catfish (Rhamdia quelen). Vet. Anaesth. Analg. 2014, 41, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.F.; Descovi, S.; Baldissera, M.D.; Bertolin, K.; Bianchini, A.E.; Mourão, R.H.V.; Schmidt, D.; Heinzmann, B.M.; Antoniazzi, A.; Baldisserotto, B.; et al. Involvement of HPI-axis in anesthesia with Lippia alba essential oil citral and linalool chemotypes: Gene expression in the secondary responses in silver catfish. Fish Physiol. Biochem. 2019, 45, 155–166. [Google Scholar] [CrossRef]
- Ozeki, M. The effects of eugenol on the nerve and muscle in crayfish. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1975, 50, 183–191. [Google Scholar]
- Bownik, A. Protective effects of ectoine on physiological parameters of Daphnia magna subjected to clove oil-induced anaesthesia. Turk. J. Fish. Aquat. Sci. 2016, 16, 691–701. [Google Scholar] [CrossRef]
- Cowing, D.; Powell, A.; Johnson, M. Evaluation of different concentration doses of eugenol on the behaviour of Nephrops norvegicus. Aquaculture 2015, 442, 78–85. [Google Scholar] [CrossRef]
- Heldwein, C.G.; Silva, L.L.; Reckziegel, P.; Barros, F.M.C.; Bürger, M.E.; Baldisserotto, B.; Mallmann, C.A.; Schmidt, D.; Caron, B.O.; Heinzmann, B.M. Participation of the GABAergic system in the anesthetic effect of Lippia alba (Mill.) NE Brown essential oil. Braz. J. Med. Biol. Res. 2012, 45, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Swanson, C.J.; Bures, M.; Johnson, M.P.; Linden, A.M.; Monn, J.A.; Schoepp, D.D. Metabotropic glutamate receptors as novel targets for anxiety and stress disorders. Nat. Rev. Drug Discov. 2005, 4, 131–144. [Google Scholar] [CrossRef]
- Kruschwitz, L.G. Environmental factors controlling reproduction of the amphipod Hyalella azteca. Proc. Okla. Acad. Sci. 1978, 21, 16–21. [Google Scholar]
- Muskó, I.B. Qualitative and quantitative relationships of Amphipoda (Crustacea) living on macrophytes in Lake Balaton (Hungary). In Trophic Relationships in Inland Waters; Biró, P., Talling, J.F., Eds.; Developments in Hydrobiology; Springer: Dordrecht, The Netherlands, 1990. [Google Scholar] [CrossRef]
- Wellborn, G.A. Determinants of reproductive success in freshwater amphipod species that experience different mortality regimes. Anim. Behav. 1995, 50, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Stom, D.I.; Zhdanova, G.O.; Saksonov, M.N.; Balayan, A.E.; Tolstoy, M.Y. Light avoidance in Baikalian amphipods as a test response to toxicants. Contemp. Probl. Ecol. 2017, 10, 77–83. [Google Scholar] [CrossRef]
- Fossat, P.; Bacqué-Cazenave, J.; De Deurwaerdère, P.; Cattaert, D.; Delbecque, J.P. Serotonin, but not dopamine, controls the stress response and anxiety-like behavior in the crayfish Procambarus clarkii. J. Exp. Biol. 2015, 218, 2745–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crook, R.J.; Dickson, K.; Hanlon, R.T.; Walters, E.T. Nociceptive sensitization reduces predation risk. Curr. Biol. 2014, 24, 1121–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, T.J.; Kwan, G.T.; Gallup, J.; Tresguerres, M. Acute fluoxetine exposure alters crab anxiety-like behaviour, but not aggressiveness. Sci. Rep. 2016, 6, 19850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dianne, L.; Perrot-Minnot, M.J.; Bauer, A.; Guvenatam, A.; Rigaud, T. Parasite-induced alteration of plastic response to predation threat: Increased refuge use but lower food intake in Gammarus pulex infected with the acanthocephalan Pomphorhynchus laevis. Int. J. Parasitol. 2014, 44, 211–216. [Google Scholar] [CrossRef]
- Dunn, A.M.; Dick, J.T.A.; Hatcher, M.J. The less amorous Gammarus: Predation risk affects mating decisions in Gammarus duebeni (Amphipoda). Anim. Behav. 2008, 76, 1289–1295. [Google Scholar] [CrossRef]
- Mohammad, F.; Aryal, S.; Ho, J.; Stewart, J.C.; Norman, N.A.; Tan, T.L.; Eisaka, A.; Claridge-Chang, A. Ancient anxiety pathways influence Drosophila defense behaviors. Curr. Biol. 2016, 26, 981–986. [Google Scholar] [CrossRef] [Green Version]
- MacNeil, C.; Dick, J.T.; Elwood, R.W. The dynamics of predation on Gammarus spp. (Crustacea: Amphipoda). Biol. Rev. 1999, 74, 375–395. [Google Scholar] [CrossRef]
- Bossus, M.C.; Guler, Y.Z.; Short, S.J.; Morrison, E.R.; Ford, A.T. Behavioural and transcriptional changes in the amphipod Echinogammarus marinus exposed to two antidepressants, fluoxetine and sertraline. Aquat. Toxicol. 2014, 151, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Worischka, S.; Schmidt, S.I.; Hellmann, C.; Winkelmann, C. Selective predation by benthivorous fish on stream macroinvertebrates—The role of prey traits and prey abundance. Limnologica 2015, 52, 41–50. [Google Scholar] [CrossRef]
- Junior, G.B.; Abreu, M.S.; Rosa, J.G.S.; Pinheiro, C.G.; Heinzmann, B.M.; Caron, B.O.; Baldisserotto, B.; Barcellos, L.J.G. Lippia alba and Aloysia triphylla essential oils are anxiolytic without inducing aversiveness in fish. Aquaculture 2018, 482, 49–56. [Google Scholar] [CrossRef]
- Capatina, L.; Boiangiu, R.S.; Dumitru, G.; Napoli, E.M.; Ruberto, G.; Hritcu, L.; Todirascu-Ciornea, E. Rosmarinus officinalis essential oil improves scopolamine-induced neurobehavioral changes via restoration of cholinergic function and brain antioxidant status in zebrafish (Danio rerio). Antioxidants 2020, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Capparucci, F.; De Benedetto, G.; Natale, S.; Pecoraro, R.; Iaria, C.; Marino, F. Evaluation of anaesthetic effect of commercial basil Ocimum basilicum on zebrafish (Danio rerio) embryos. Fishes 2022, 7, 318. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Category | Behavior | Description |
|---|---|---|
| Speed | Mean speed | Average animal speed as a function of the distance traveled in the aquarium. |
| Maximum speed | Maximum speed of the animal in the aquarium. | |
| Maneuvering | Absolute turning angle | The sum of the absolute angle between each movement vector of the animal. |
| Immobility | Freezing episodes | Number of times the animal froze during the test. |
| Time freezing | Total amount of time during the test for which the animal was freezing. | |
| Tank exploration | Total distance | Sum of the total distance that the animal travelled between each point during the test. |
| Crossings between the tank zones | Numbers of crossings of the animal between the different zones of the aquarium. | |
| Entries in each virtual zone | Counts the number of times the animal entered in each zone. | |
| Dwelling time in each zone | Total amount of time the animal spent in the zone. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, A.J.; Santos, S.; Heinzmann, B.M.; Caron, B.O.; Baldisserotto, B. Exposure of Hyalella bonariensis (Crustacea, Amphipoda) to Essential Oils: Effects on Anesthesia and Swimming Activity. Fishes 2023, 8, 149. https://doi.org/10.3390/fishes8030149
Becker AJ, Santos S, Heinzmann BM, Caron BO, Baldisserotto B. Exposure of Hyalella bonariensis (Crustacea, Amphipoda) to Essential Oils: Effects on Anesthesia and Swimming Activity. Fishes. 2023; 8(3):149. https://doi.org/10.3390/fishes8030149
Chicago/Turabian StyleBecker, Alessandra Janaína, Sandro Santos, Berta Maria Heinzmann, Braulio Otomar Caron, and Bernardo Baldisserotto. 2023. "Exposure of Hyalella bonariensis (Crustacea, Amphipoda) to Essential Oils: Effects on Anesthesia and Swimming Activity" Fishes 8, no. 3: 149. https://doi.org/10.3390/fishes8030149