
Two Probiotic Candidates of the Genus Psychrobacter Modulate the Immune Response and Disease Resistance after Experimental Infection in Turbot (Scophthalmus maximus, Linnaeus 1758)

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Probiotic Candidates
2.2. Probiotic Treatment
2.3. Determination of LD50 for Experimental Infection
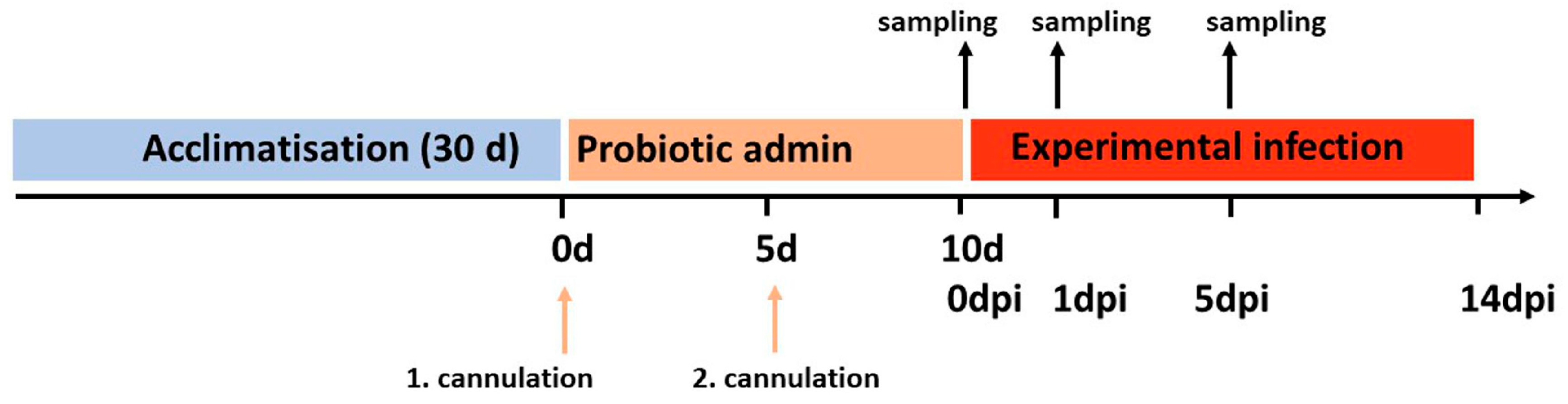
2.4. Experimental Infection and Sampling
2.5. Gene Expression
2.6. Statistical Analysis
3. Results
3.1. Mortality
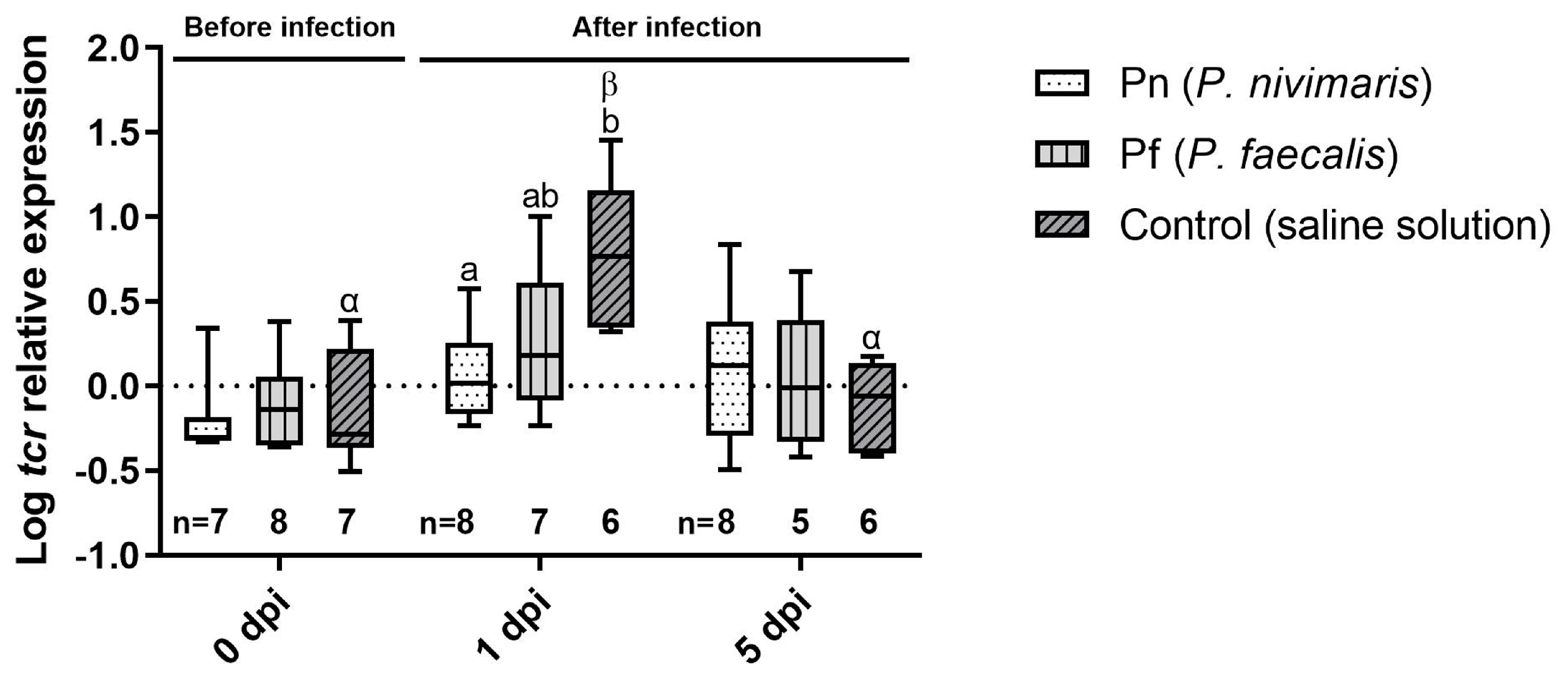
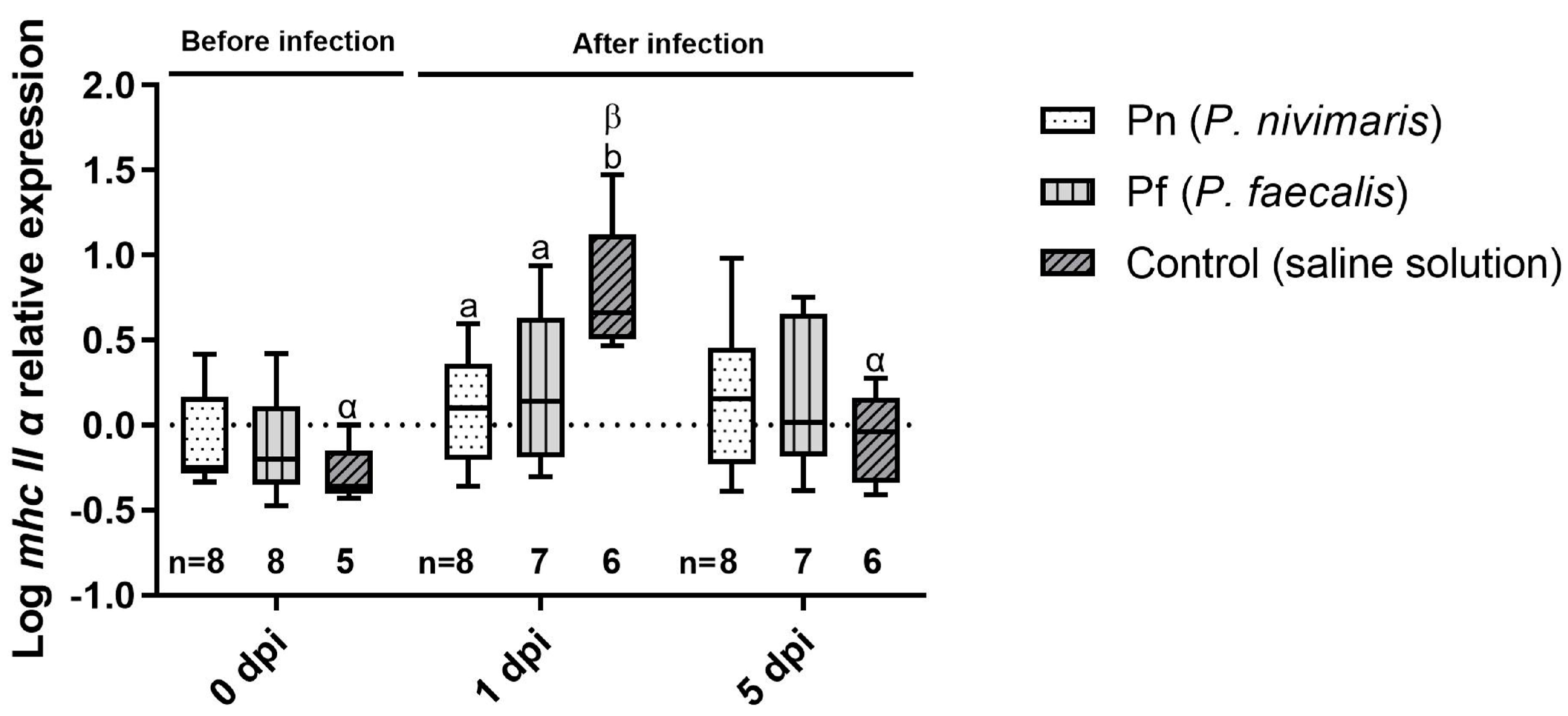
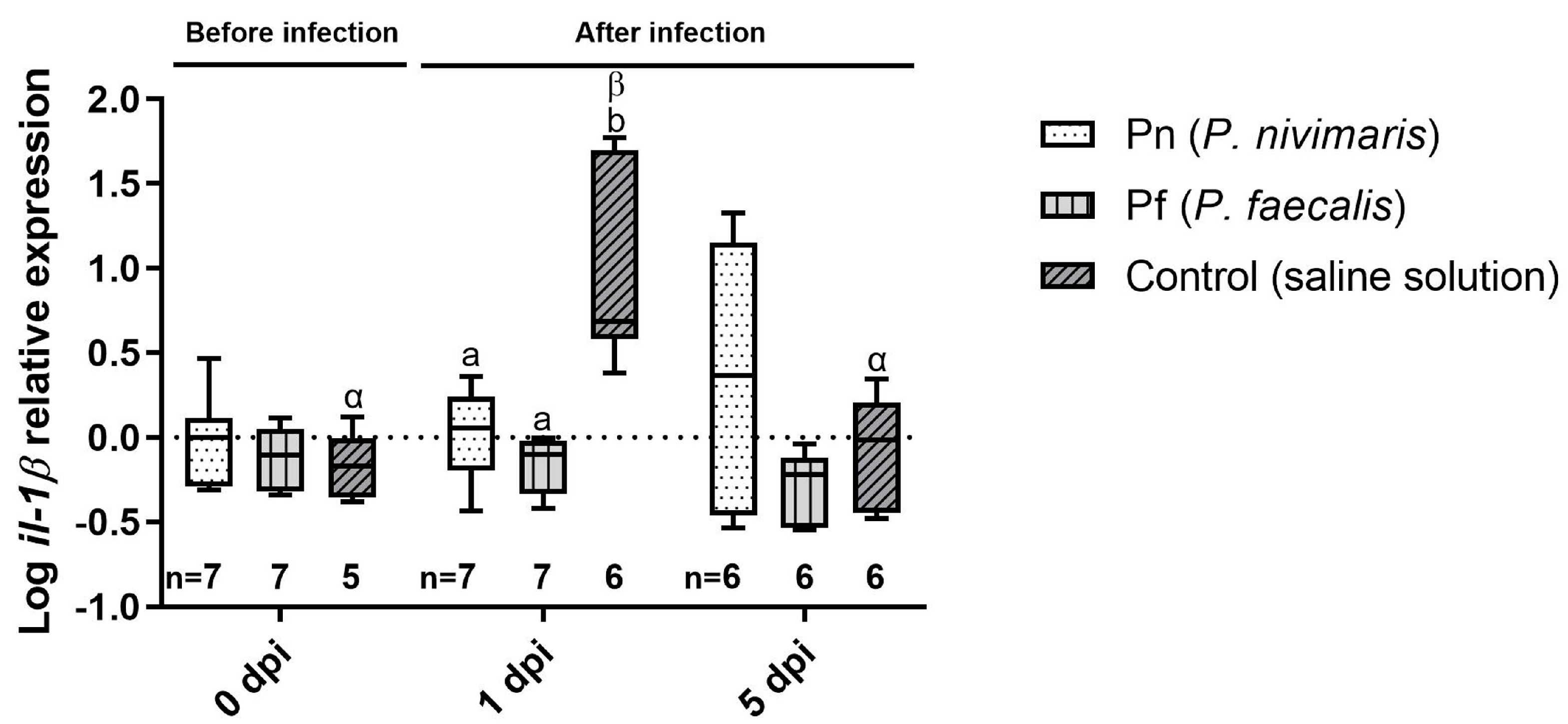
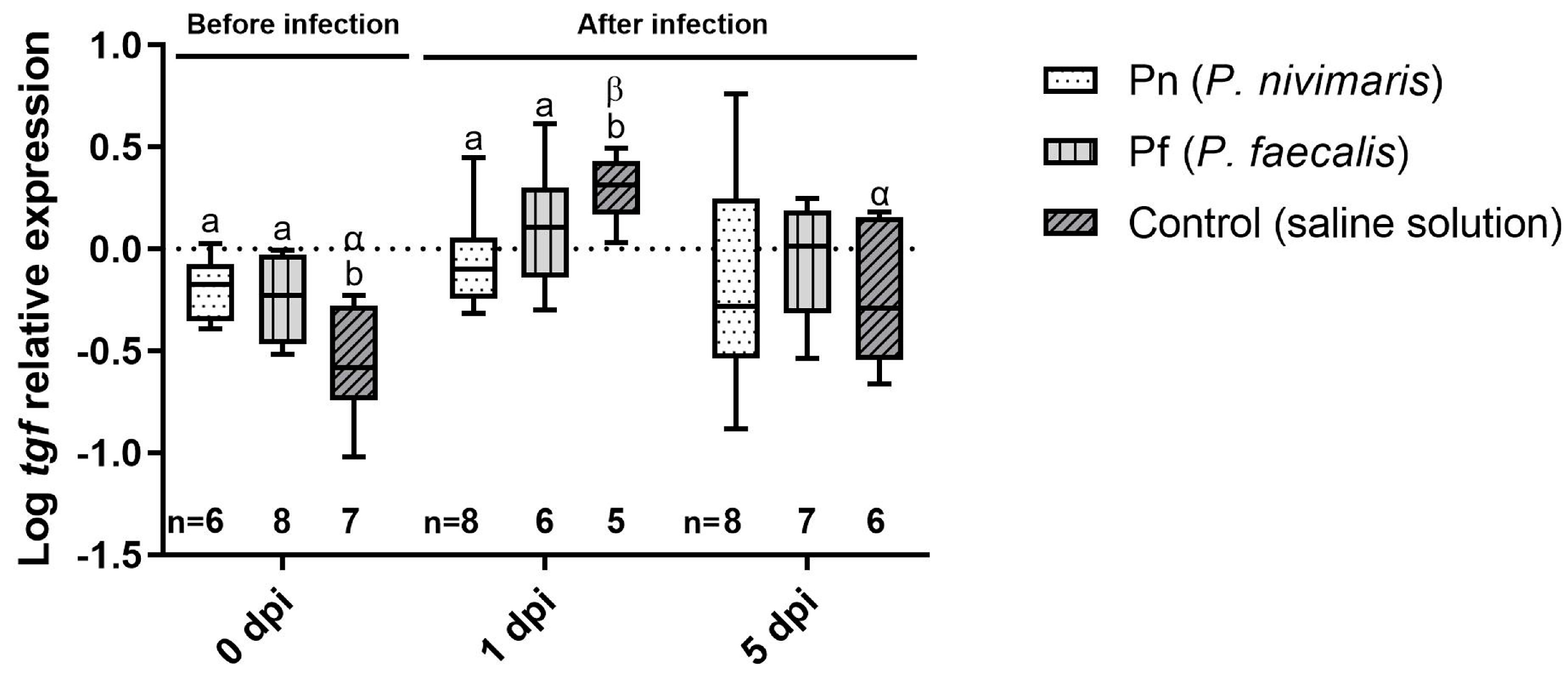
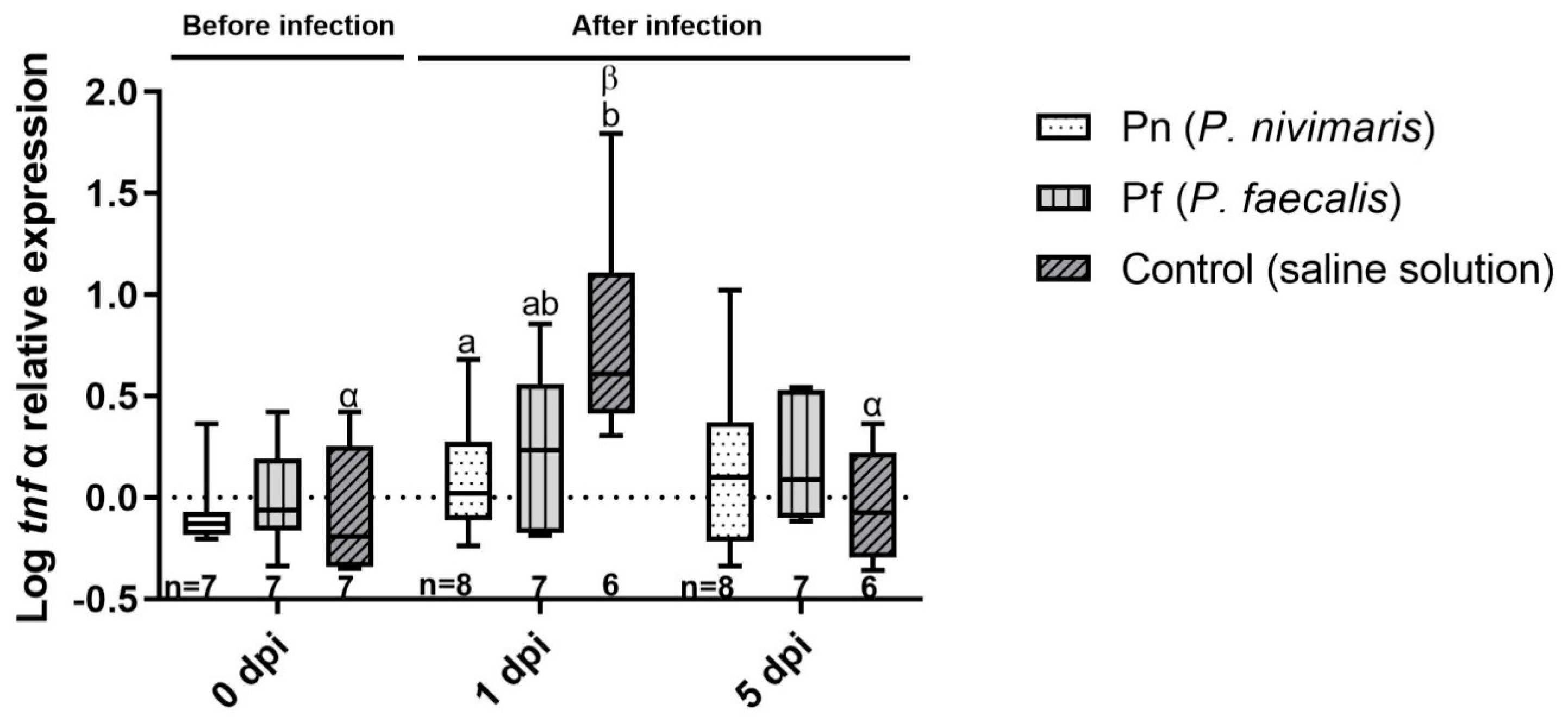
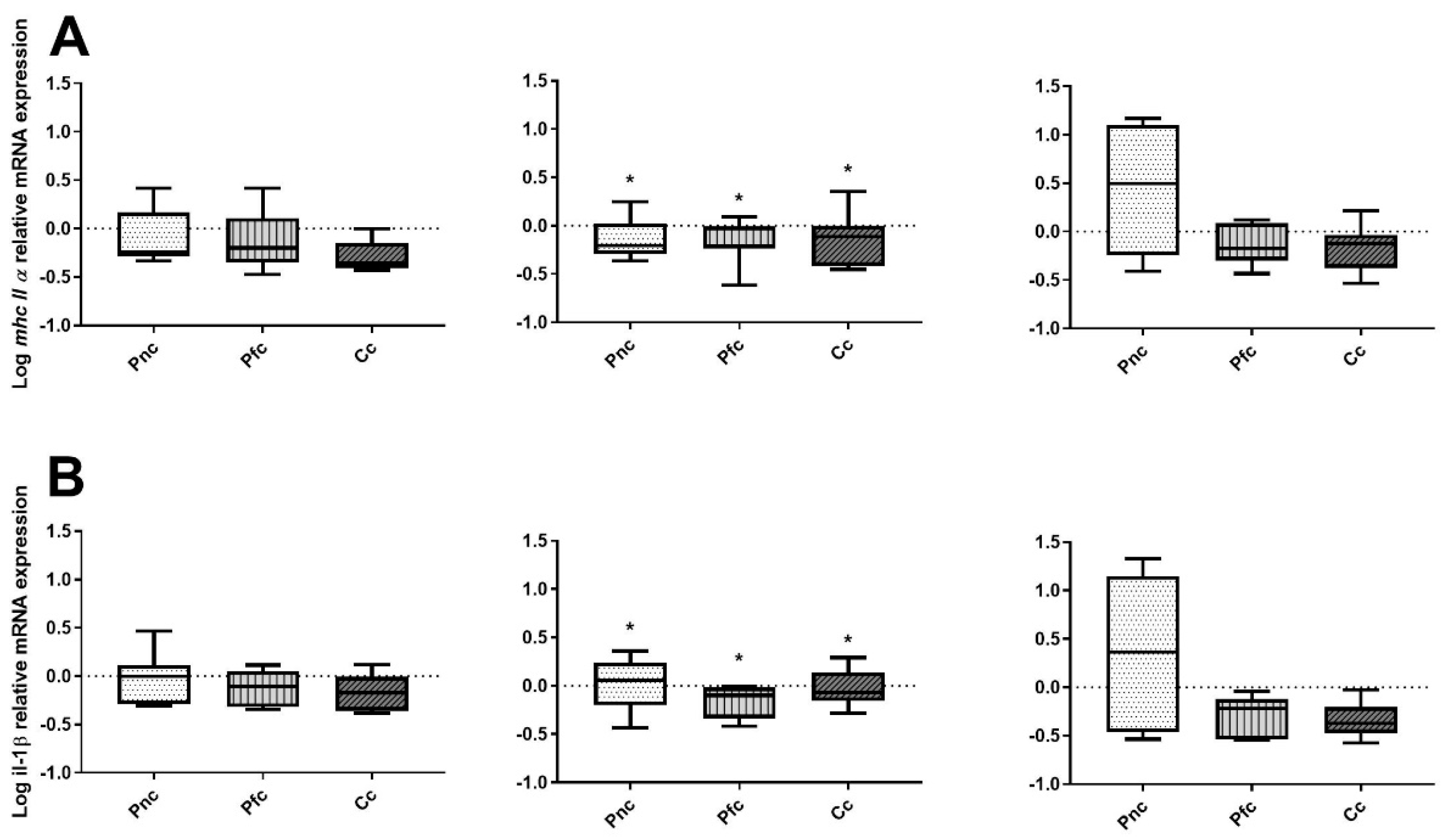
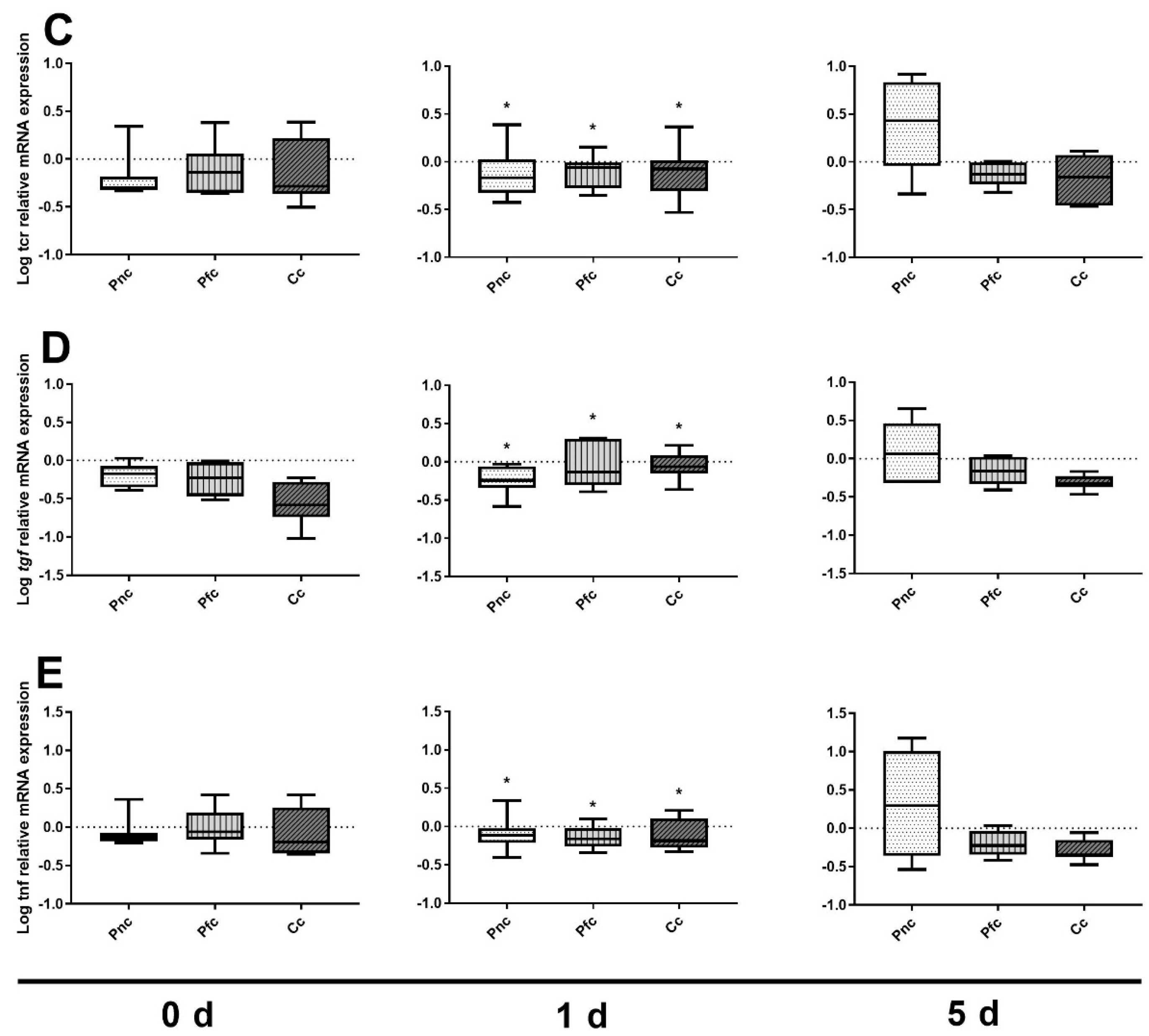
3.2. Immune Marker Gene Expression
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, N. Flatfish farming systems in the Atlantic region. Rev. Fish. Sci. 2002, 10, 403–419. [Google Scholar] [CrossRef]
- Reiser, S.; Schroeder, J.P.; Wuertz, S.; Kloas, W.; Hanel, R. Histological and physiological alterations in juvenile turbot (Psetta maxima, L.) exposed to sublethal concentrations of ozone-produced oxidants in ozonated seawater. Aquaculture 2010, 307, 157–164. [Google Scholar] [CrossRef]
- Imsland, A.K.; Folkvord, A.; Stefansson, S.O. Growth, oxygen consumption and activity of juvenile turbot (Scophthalmus maximus L.) reared under different temperatures and photoperiods. Neth. J. Sea Res. 1995, 34, 149–159. [Google Scholar] [CrossRef]
- Imsland, A.K.; Sunde, L.M.; Folkvord, A.; Stefansson, S.O. The interaction of temperature and fish size on growth of juvenile turbot. J. Fish Biol. 1996, 49, 926–940. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364, 345–352. [Google Scholar] [CrossRef]
- Wanka, K.M.; Schulz, C.; Kloas, W.; Wuertz, S. Administration of host-derived probiotics does not affect utilization of soybean meal enriched diets in juvenile turbot (Scophthalmus maximus). J. Appl. Ichthyol. 2019, 35, 1004–1015. [Google Scholar] [CrossRef]
- Waring, C.P.; Poxton, M.G.; Stagg, R.M. The physiological response of the turbot to multiple net confinements. Aquac. Int. 1997, 5, 1–12. [Google Scholar] [CrossRef]
- Waring, C.P.; Stagg, R.M.; Poxton, M.G. Physiological responses to handling in the turbot. J. Fish Biol. 1996, 48, 161–173. [Google Scholar] [CrossRef]
- Reiser, S.; Wuertz, S.; Schroeder, J.P.; Kloas, W.; Hanel, R. Risks of seawater ozonation in recirculation aquaculture—Effects of oxidative stress on animal welfare of juvenile turbot (Psetta maxima, L.). Aquat. Toxicol. 2011, 105, 508–517. [Google Scholar] [CrossRef]
- van Bussel, C.G.J.; Schroeder, J.P.; Wuertz, S.; Schulz, C. The chronic effect of nitrate on production performance and health status of juvenile turbot (Psetta maxima). Aquaculture 2012, 326, 163–167. [Google Scholar] [CrossRef]
- Hermann, B.T.; Wuertz, S.; Vanselow, K.H.; Schulz, C.; Stiller, K.T. Divergent gene expression in the gills of juvenile turbot (Psetta maxima) exposed to chronic severe hypercapnia indicates dose-dependent increase in intracellular oxidative stress and hypoxia. Aquat. Toxicol. 2019, 206, 72–80. [Google Scholar] [CrossRef]
- Zhang, X.J.; Zhou, H.H.; Liu, C.D.; Mai, K.S.; He, G.; Wang, X. Fishmeal substitution with low-gossypol cottonseed meal in the diet for juvenile turbot (Scophthalmus maximus L.): Effects on growth, nutrients utilization and haematological responses. Aquacult. Rep. 2022, 24, 101149. [Google Scholar] [CrossRef]
- Cao, S.H.; Li, P.Y.; Huang, B.S.; Song, Z.D.; Hao, T.T.; Wang, C.Q.; Wang, M.Q. Assessing feasibility of replacement of fishmeal with enzyme-treated feather meal in the diet of juvenile turbot (Scophthalmus maximus L.). Aquacult. Nutr. 2020, 26, 1340–1352. [Google Scholar] [CrossRef]
- Arnason, J.; Imsland, A.K.D.; Helmig, T.; Gunnarsson, S.; Kristjansson, G.O. Fishmeal replacement by mixed plant proteins and effect on growth and sensory attributes in on-growing turbot. Aquacult. Nutr. 2018, 24, 1041–1047. [Google Scholar] [CrossRef]
- Mulcahy, M.F. Diseases of flatfish. Bull.-Eur. Assoc. Fish Pathol. 2002, 22, 86–94. [Google Scholar]
- Sandlund, N.; Rodseth, O.M.; Knappskog, D.H.; Fiksdal, I.U.; Bergh, O. Comparative susceptibility of turbot, halibut, and cod yolk-sac larvae to challenge with Vibrio spp. Dis. Aquat. Org. 2010, 89, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Du, H.F.; Zhang, Y.H.; Zhang, M.; Liu, Q.A.; Zhu, H.J.; Cao, F. Marine fungal metabolites as a source of drug leads against aquatic pathogens. Appl. Microbiol. Biot. 2022, 106, 3337–3350. [Google Scholar] [CrossRef]
- Meyer, F.P. Aquaculture disease and health management. J. Anim. Sci. 1991, 69, 4201–4208. [Google Scholar] [CrossRef]
- Bondad-Reantaso, M.G.; Subasinghe, R.P.; Arthur, J.R.; Ogawa, K.; Chinabut, S.; Adlard, R.; Tan, Z.; Shariff, M. Disease and health management in Asian aquaculture. Vet. Parasitol. 2005, 132, 249–272. [Google Scholar] [CrossRef]
- Dey, M.M.; Briones, R.M.; Garcia, Y.T.; Nissapa, A.; Rodriguez, U.P.; Talukder, R.K.; Senaratne, A.; Omar, I.H.; Koeshendrajana, S.; Khiem, N.T.; et al. Strategies and Options for Increasing and Sustaining Fisheries and Aquaculture Production to Benefit Poorer Households in Asia; WorldFish: Penang, Malaysia, 2008. [Google Scholar]
- Zhang, Z.; Wang, Y.G.; Yang, G.P.; Li, Q.F. The present status of research on bacterial diseases of turbots Scophthalmus maximus. Trans. Oceanol. Limnol. 2004, 3, 83–89. [Google Scholar]
- Coscelli, G.A.; Bermudez, R.; Silva, A.R.S.; de Ocenda, M.V.R.; Quiroga, M.I. Granulomatous dermatitis in turbot (Scophthalmus maximus L.) associated with natural Aeromonas salmonicida subsp salmonicida infection. Aquaculture 2014, 428, 111–116. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magarinos, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Y.H.; Sun, Q.L.; Li, X.P.; Sun, L. An inactivated bivalent vaccine effectively protects turbot (Scophthalmus maximus) against Vibrio anguillarum and Vibrio harveyi infection. Aquaculture 2021, 544, 737158. [Google Scholar] [CrossRef]
- Lillehaug, A.; Lunestad, B.T.; Grave, K. Epidemiology of bacterial diseases in Norwegian aquaculture—A description based on antibiotic prescription data for the ten-year period 1991 to 2000. Dis. Aquat. Org. 2003, 53, 115–125. [Google Scholar] [CrossRef]
- Noga, E.J.; Smith, S.A.; Ottesen, O.H. Bacterial diseases. In Practical Flatfish Culture and Stock Enhancement; Daniels, H.V., Watanabe, W., Eds.; Wiley: Hoboken, NJ, USA, 2010; pp. 265–268. [Google Scholar]
- Wallace, I.S.; McKay, P.; Murray, A.G. A historical review of the key bacterial and viral pathogens of Scottish wild fish. J. Fish Dis. 2017, 40, 1741–1756. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, Y.G.; Zhang, Z.; Liao, M.J.; Li, B.; Rong, X.J.; Chen, G.P. Responses of microbial community structure in turbot (Scophthalmus maximus) larval intestine to the regulation of probiotic introduced through live feed. PLoS ONE 2019, 14, e0216590. [Google Scholar] [CrossRef] [Green Version]
- Masumura, K.; Wakabayashi, H. An outbreak of gliding bacterial disease in hatchery-born red seabream (Pagrus major) and gilthead (Acanthopagrus schlegeli) fry in Hiroshima. Fish Pathol. 1977, 12, 171–177. [Google Scholar] [CrossRef]
- Avendano-Herrera, R.; Toranzo, A.E.; Magarinos, B. Tenacibaculosis infection in marine fish caused by Tenacibaculum maritimum: A review. Dis. Aquat. Org. 2006, 71, 255–266. [Google Scholar] [CrossRef]
- Nowlan, J.P.; Lumsden, J.S.; Russell, S. Advancements in characterizing Tenacibaculum infections in Canada. Pathogens 2020, 9, 1029. [Google Scholar] [CrossRef]
- Mitchell, S.O.; Rodger, H.D. A review of infectious gill disease in marine salmonid fish. J. Fish Dis. 2011, 34, 411–432. [Google Scholar] [CrossRef]
- Avendano-Herrera, R.; Magarinos, B.; Morinigo, M.A.; Romalde, J.L.; Toranzo, A.E. A novel O-serotype in Tenacibaculum maritimum strains isolated from cultured sole (Solea senegalensis). Bull. Eur. Assoc. Fish Pathol. 2005, 25, 70–74. [Google Scholar]
- Bernardet, J.F.; Campbell, A.C.; Buswell, J.A. Flexibacter maritimus is the agent of Black Patch Necrosis in Dover sole in Scotland. Dis. Aquat. Org. 1990, 8, 233–237. [Google Scholar] [CrossRef]
- Gourzioti, E.; Kolygas, M.N.; Athanassopoulou, F.; Babili, V. Tenacibaculosis in aquaculture farmed marine fish. J. Hell. Vet. Med. Soc. 2016, 67, 21–32. [Google Scholar] [CrossRef]
- Delannoy, C.M.J.; Houghton, J.D.R.; Fleming, N.E.C.; Ferguson, H.W. Mauve stingers (Pelagia noctiluca) as carriers of the bacterial fish pathogen Tenacibaculum maritimum. Aquaculture 2011, 311, 255–257. [Google Scholar] [CrossRef]
- Rico, A.; Satapornvanit, K.; Haque, M.M.; Min, J.; Nguyen, P.T.; Telfer, T.C.; van den Brink, P.J. Use of chemicals and biological products in Asian aquaculture and their potential environmental risks: A critical review. Rev. Aquacult. 2012, 4, 75–93. [Google Scholar] [CrossRef]
- Sorum, H. Antimicrobial drug resistance in fish pathogens. In Antimicrobial Resistance in Bacteria of Animal Origin; American Society of Microbiology: Washington, DC, USA, 2006. [Google Scholar]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dolz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial use in aquaculture re-examined: Its relevance to antimicrobial resistance and to animal and human health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, J.; Xiao, P.; Li, G.Y.; Yue, S.; Huang, J.; Zhu, W.Y.; Mo, Z.L. Isolation and characterization of Bacillus spp. M001 for potential application in turbot (Scophthalmus maximus L.) against Vibrio anguillarum. Aquacult. Nutr. 2016, 22, 374–381. [Google Scholar] [CrossRef]
- Wuertz, S.; Schroeder, A.; Wanka, K.M. Probiotics in fish nutrition—Long-standing household remedy or native nutraceuticals? Water 2021, 13, 1348. [Google Scholar] [CrossRef]
- Wanka, K.M.; Damerau, T.; Costas, B.; Krueger, A.; Schulz, C.; Wuertz, S. Isolation and characterization of native probiotics for fish farming. BMC Microbiol. 2018, 18, 119. [Google Scholar] [CrossRef] [Green Version]
- Caipang, C.M.A.; Brinchmann, M.F.; Kiron, V. Antagonistic activity of bacterial isolates from intestinal microbiota of Atlantic cod, Gadus morhua, and an investigation of their immunomodulatory capabilities. Aquac. Res. 2010, 41, 249–256. [Google Scholar] [CrossRef]
- Lazado, C.C.; Caipang, C.M.A.; Rajan, B.; Brinchmann, M.F.; Kiron, V. Characterization of GP21 and GP12: Two potential probiotic bacteria isolated from the gastrointestinal tract of Atlantic cod. Probiot. Antimicrob. Proteins 2010, 2, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Chu, T.W.; Chen, C.N.; Pan, C.Y. Antimicrobial status of tilapia (Oreochromis niloticus) fed Enterococcus avium originally isolated from goldfish intestine. Aquac. Rep. 2020, 17, 100397. [Google Scholar] [CrossRef]
- Zhang, D.X.; Kang, Y.H.; Zhan, S.; Zhao, Z.L.; Jin, S.N.; Chen, C.; Zhang, L.; Shen, J.Y.; Wang, C.F.; Wang, G.Q.; et al. Effect of Bacillus velezensis on Aeromonas veronii-induced intestinal mucosal barrier function damage and inflammation in Crucian carp (Carassius auratus). Front. Microbiol. 2019, 10, 2663. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.C.; Chang, X.L.; Zhang, Y.R.; Yan, X.; Zhang, J.X.; Nie, G.X. Effects of Lactococcus lactis from Cyprinus carpio L. as probiotics on growth performance, innate immune response and disease resistance against Aeromonas hydrophila. Fish Shellfish Immunol. 2019, 93, 73–81. [Google Scholar] [CrossRef]
- Chauhan, A.; Singh, R. Isolation and evaluation of putative probiotic strains from different teleost to prevent Pseudomonas aeruginosa infection in Cyprinus carpio. Aquac. Res. 2019, 50, 3616–3627. [Google Scholar] [CrossRef]
- Ghosh, B.; Cain, K.D.; Nowak, B.F.; Bridle, A.R. Microencapsulation of a putative probiotic Enterobacter species, C6-6, to protect rainbow trout, Oncorhynchus mykiss (Walbaum), against bacterial coldwater disease. J. Fish Dis. 2016, 39, 1–11. [Google Scholar] [CrossRef]
- Makled, S.O.; Hamdan, A.M.; El-Sayed, A.F.M.; Hafez, E.E. Evaluation of marine psychrophile, Psychrobacter namhaensis SO89, as a probiotic in Nile tilapia (Oreochromis niloticus) diets. Fish Shellfish Immunol. 2017, 61, 194–200. [Google Scholar] [CrossRef]
- Sun, Y.Z.; Yang, H.L.; Ma, R.L.; Zhang, C.X.; Lin, W.Y. Effect of dietary administration of Psychrobacter sp. on the growth, feed utilization, digestive enzymes and immune responses of grouper Epinephelus coioides. Aquac. Nutr. 2011, 17, E733–E740. [Google Scholar] [CrossRef]
- Herias, M.V.; Hessle, C.; Telemo, E.; Midtvedt, T.; Hanson, L.A.; Wold, A.E. Immunomodulatory effects of Lactobacillus plantarum colonizing the intestine of gnotobiotic rats. Clin. Exp. Immunol. 1999, 116, 283–290. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Satoh, S.; Hirono, I.; Kobayshi, T.; Sugita, H.; Puangkaew, J.; Aoki, T. Immune modulation and expression of cytokine genes in rainbow trout Oncorhynchus mykiss upon probiotic feeding. Dev. Comp. Immunol. 2007, 31, 372–382. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Foysal, M.J.; Fotedar, R.; Francis, D.S.; Gupta, S.K. Probiotic yeast Saccharomyces cerevisiae coupled with Lactobacillus casei modulates physiological performance and promotes gut microbiota in juvenile barramundi, Lates calcarifer. Aquaculture 2022, 546, 737346. [Google Scholar] [CrossRef]
- Shadrack, R.S.; Manabu, I.; Koshio, S.; Yokoyama, S.; Zhang, Y.K.; Mzengereza, K.; El Basuini, M.F.; Dawood, M.A.O. Effects of single and mixture probiotic supplements on growth, digestive activity, antioxidative status, immune and growth-related genes, and stress response of juvenile Red Sea Bream (Pagrus major). Aquac. Nutr. 2022, 2022, 8968494. [Google Scholar] [CrossRef]
- Bowman, J.P. The Genus Psychrobacter. In The Prokaryotes, Vol. 6: Proteobacteria Gamma Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 920–930. [Google Scholar]
- Jammal, A.; Bariche, M.; zu Dohna, H.; Kambris, Z. Characterization of the cultivable gut microflora in wild-caught Mediterranean fish species. Curr. Nutr. Food Sci. 2017, 13, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, K.G.; Hsu, C.K.; Pettengill, J.B.; Ihrie, J.; Karathia, H.; Hasan, N.A.; Grim, C.J. Microbiome population dynamics of cold-smoked Sockeye salmon during refrigerated storage and after culture enrichment. J. Food Prot. 2022, 85, 238–253. [Google Scholar] [CrossRef]
- Lazado, C.C.; Caipang, C.M.A.; Brinchmann, M.F.; Kiron, V. In vitro adherence of two candidate probiotics from Atlantic cod and their interference with the adhesion of two pathogenic bacteria. Vet. Microbiol. 2011, 148, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Lazado, C.C.; Caipang, C.M.A.; Gallage, S.; Brinchmann, M.F.; Kiron, V. Expression profiles of genes associated with immune response and oxidative stress in Atlantic cod, Gadus morhua head kidney leukocytes modulated by live and heat-inactivated intestinal bacteria. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 155, 249–255. [Google Scholar] [CrossRef]
- Winkelbach, A.; Schade, R.; Schulz, C.; Wuertz, S. Comparison of oral, rectal and intraperitoneal administration of IgY antibodies in passive immunization of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2015, 23, 427–438. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Yu, Y.; Li, H.R.; Luo, W. Psychrobacter fjordensis sp nov., a psychrotolerant bacterium isolated from an Arctic fjord in Svalbard. Antonie Van Leeuwenhoek 2015, 108, 1283–1292. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Yu, Y.; Liu, Y.; Li, H.R. Psychrobacter glaciei sp nov., isolated from the ice core of an Arctic glacier. J. Syst. Evol. Microbiol. 2016, 66, 1792–1798. [Google Scholar] [CrossRef]
- Avendano-Herrera, R.; Toranzo, A.E.; Magarinos, B. A challenge model for Tenacibaculum maritimum infection in turbot, Scophthalmus maximus (L.). J. Fish Dis. 2006, 29, 371–374. [Google Scholar] [CrossRef]
- Mabrok, M.; Machado, M.; Serra, C.R.; Afonso, A.; Valente, L.M.P.; Costas, B. Tenacibaculosis induction in the Senegalese sole (Solea senegalensis) and studies of Tenacibaculum maritimum survival against host mucus and plasma. J. Fish Dis. 2016, 39, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method for estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 23 February 2023).
- Murall, C.L.; Abbate, J.L.; Touzel, M.P.; Allen-Vercoe, E.; Alizon, S.; Froissart, R.; McCann, K. Chapter five—Invasions of host-associated microbiome networks. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Massol, F., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 57, p. 80. [Google Scholar]
- Ibrahem, M.D. Evolution of probiotics in aquatic world: Potential effects, the current status in Egypt and recent prospectives. J. Adv. Res. 2015, 6, 765–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.Z.; Yang, H.L.; Ling, Z.C.; Chang, J.B.; Ye, J.D. Gut microbiota of fast and slow growing grouper Epinephelus coioides. Afr. J. Microbiol. Res. 2009, 3, 713–720. [Google Scholar]
- Kakizaki, E.; Takahama, K.; Seo, Y.; Kozawa, S.; Sakai, M.; Yukawa, N. Marine bacteria comprise a possible indicator of drowning in seawater. Forensic Sci. Int. 2008, 176, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Shang, D.D.; Yang, G.J.; Chen, G.J.; Du, Z.J. Psychrobacter halodurans sp. nov. and Psychrobacter coccoides sp. nov., two new slightly halophilic bacteria isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 2022, 72, 005173. [Google Scholar] [CrossRef] [PubMed]
- Cantas, L.; Fraser, T.W.K.; Fjelldal, P.G.; Mayer, I.; Sorum, H. The culturable intestinal microbiota of triploid and diploid juvenile Atlantic salmon (Salmo salar)—A comparison of composition and drug resistance. BMC Vet. Res. 2011, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Datta, A.A.; Sharma, A.K.; Kundu, R.; Singh, S.P. Diversity and enzymatic profile of bacterial flora in the gut of an estuarine fish, Mugil jerdoni. Indian J. Geo-Mar. Sci. 2017, 46, 1116–1127. [Google Scholar]
- Etyemez, M.; Balcazar, J.L. Bacterial community structure in the intestinal ecosystem of rainbow trout (Oncorhynchus mykiss) as revealed by pyrosequencing-based analysis of 16S rRNA genes. Res. Vet. Sci. 2015, 100, 8–11. [Google Scholar] [CrossRef]
- Svanevik, C.S.; Lunestad, B.T. Characterisation of the microbiota of Atlantic mackerel (Scomber scombrus). Int. J. Food Microbiol. 2011, 151, 164–170. [Google Scholar] [CrossRef]
- Klemetsen, T.; Willassen, N.P.; Karlsen, C.R. Full-length 16S rRNA gene classification of Atlantic salmon bacteria and effects of using different 16S variable regions on community structure analysis. Microbiologyopen 2019, 8, e898. [Google Scholar] [CrossRef]
- Tennant, S.M.; Hartland, E.L.; Phumoonna, T.; Lyras, D.; Rood, J.I.; Robins-Browne, R.M.; van Driel, I.R. Influence of gastric acid on susceptibility to infection with ingested bacterial pathogens. Infect. Immun. 2008, 76, 639–645. [Google Scholar] [CrossRef] [Green Version]
- Koelz, H.R. Gastric-acid in vertebrates. Scand. J. Gastroenterol. 1992, 27, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Howden, C.W.; Hunt, R.H. Relationship between gastric secretion and infection. Gut 1987, 28, 96–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisch, K.; Smage, S.B.; Johansen, R.; Duesund, H.; Brevik, O.J.; Nylund, A. Pathology of experimentally induced mouthrot caused by Tenacibaculum maritimum in Atlantic salmon smolts. PLoS ONE 2018, 13, e0206951. [Google Scholar] [CrossRef] [PubMed]
- Costas, B.; Simoes, I.; Castro-Cunha, M.; Afonso, A. Non-specific immune responses of Senegalese sole, Solea senegalensis (Kaup), head-kidney leucocytes against Tenacibaculum maritimum. J. Fish Dis. 2014, 37, 765–769. [Google Scholar] [CrossRef]
- Millan, A.; Gomez-Tato, A.; Pardo, B.G.; Fernandez, C.; Bouza, C.; Vera, M.; Alvarez-Dios, J.A.; Cabaleiro, S.; Lamas, J.; Lemos, M.L.; et al. Gene expression profiles of the spleen, liver, and head kidney in turbot (Scophthalmus maximus) Along the infection process with Aeromonas salmonicida using an immune-enriched oligo-microarray. Mar. Biotechnol. 2011, 13, 1099–1114. [Google Scholar] [CrossRef]
- Massague, J. The transforming growth-factor-beta family. Annu. Rev. Cell Biol. 1990, 6, 597–641. [Google Scholar] [CrossRef]
- Liu, X.H.; Zhang, H.; Jiao, C.L.; Liu, Q.; Zhang, Y.X.; Xiao, J.F. Flagellin enhances the immunoprotection of formalin-inactivated Edwardsiella tarda vaccine in turbot. Vaccine 2017, 35, 369–374. [Google Scholar] [CrossRef]
- Wuertz, S.; Beça, F.; Kreuz, E.; Wanka, K.M.; Azeredo, R.; Machado, M.; Costas, B. Effects of Native Probiotic Candidates on the Immune Status, Inflammatory Response and Disease Resistance of Turbot (Scophthalmus Maximus, Linnaeus 1758); Research Square: Durham, NC, USA, 2022. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | 5’–3’ Sequence | Ta [°C] | Length [bp] | Eff. [%] | GeneBank |
|---|---|---|---|---|---|---|
| rpl8 * | F | CTCCGCCACATTGACTTC | 64 | 197 | 94 | DQ848874 |
| R | GCCTTCTTGCCACAGTAG | |||||
| Tcr | F | ACGCCAATCACACGGTCA | 63 | 129 | 116 | AY303762 |
| R | ATCCGAACTGCTCTCGTGG | |||||
| tgf β | F | GCTACCATGCCAACTACTGC | 64 | 101 | 109 | AJ276709 |
| R | TCCCGGGTTGTGATGCTT | |||||
| tnf α | F | ATCCCCACTCCACGCTGA | 65 | 224 | 95 | FJ654645 |
| R | CGTCCTTGCTGTCATCGTC | |||||
| mhc II α | F | GATCCTCCTTCCAGTCCGAT | 63 | 140 | 105 | DQ094170 |
| R | AATGTTGAGACTCGCTCCC | |||||
| il-1β | F | CAGAAATCGCACCATGTCG | 62 | 191 | 98 | AJ295836 |
| R | GACAACCGCAAAGTTAACCTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wuertz, S.; Beça, F.; Kreuz, E.; Wanka, K.M.; Azeredo, R.; Machado, M.; Costas, B. Two Probiotic Candidates of the Genus Psychrobacter Modulate the Immune Response and Disease Resistance after Experimental Infection in Turbot (Scophthalmus maximus, Linnaeus 1758). Fishes 2023, 8, 144. https://doi.org/10.3390/fishes8030144
Wuertz S, Beça F, Kreuz E, Wanka KM, Azeredo R, Machado M, Costas B. Two Probiotic Candidates of the Genus Psychrobacter Modulate the Immune Response and Disease Resistance after Experimental Infection in Turbot (Scophthalmus maximus, Linnaeus 1758). Fishes. 2023; 8(3):144. https://doi.org/10.3390/fishes8030144
Chicago/Turabian StyleWuertz, Sven, Filipa Beça, Eva Kreuz, Konrad M. Wanka, Rita Azeredo, Marina Machado, and Benjamin Costas. 2023. "Two Probiotic Candidates of the Genus Psychrobacter Modulate the Immune Response and Disease Resistance after Experimental Infection in Turbot (Scophthalmus maximus, Linnaeus 1758)" Fishes 8, no. 3: 144. https://doi.org/10.3390/fishes8030144