1. Introduction
Interest in fish welfare is a matter of increasing importance in aquaculture. With the intensification of aquaculture practices, fish are submitted to daily stressful situations, including loss of water quality, handling, and transport, any of which may decrease growth performance due to the energy use toward stress mitigation, as well as increase their predisposition to diseases [
1,
2,
3]. Stress is the reaction of animals to a threat, altering their homeostasis, and consequently launching an endocrine stress response that releases cortisol into the bloodstream. To minimize stress, several options are being explored, including the use of specific nutrients or functional ingredients, such as probiotics, prebiotics, vitamins, minerals, nucleotides, and functional amino acids [
4,
5,
6]. Tryptophan (Trp) is an essential amino acid required for protein synthesis and is the only precursor of the neurotransmitter serotonin (5-HT), which itself is the precursor for melatonin, both having a vital role in stress mitigation. In addition, it is commercially available as a feed-grade product, making the supplementation of Trp a viable and economic way to optimize both production and animal welfare [
7]. Tryptophan may act as a functional amino acid due to its pivotal role in serotonin and melatonin synthesis, which are important in stress mitigation [
8,
9]. Previous studies reported that Trp-supplemented feeds affected fish behavior through 5-HT signaling, decreasing aggression, and attenuating stress-induced anorexia in several teleost species [
8,
10,
11,
12,
13]. Moreover, studies have shown that Trp is an important limiting factor in serotonin synthesis, causing a dose-dependent response. A deficiency in Trp can result in affective disorders, anxiety, aggression, stress, and eating disorders, among other conditions [
14,
15,
16]. Tryptophan-supplemented diets have been shown to lower plasma cortisol levels in
Oncorhynchus mykiss [
17]. Tryptophan is a widely studied functional amino acid. In meagre, the effects of a dietary supplementation of moderate levels (0.16 and 0.11% diet) have been studied in fish maintained under chronic or crowding stress [
3,
18]. Few reports have documented the effects of Trp-enriched diets on meagre, and the ones that have done so mainly tested on acute and chronic stressors.
Meagre (
Argyrosomus regius) is a Mediterranean carnivorous species with a high potential for marine aquaculture diversification [
19,
20,
21,
22,
23]. Meagre has several attractive qualities, such as a high growth rate and feed conversion efficiency, good nutritional value, and can grow in captivity, all of which are promising qualities for a future mass market species. Meagre feed requirements are similar to those of other Mediterranean marine species, such as gilthead seabream [
19,
20]. In 2020, European production of meagre reached almost 45,000 tones, representing an increase of 8.2% compared to the previous year [
24]. However, meagre production is highly impacted by stressful conditions, due to the susceptibility of this species to stress and its impact on growth performance and the overall welfare of animals. For this reason, the use of nutritional tools to reduce meagre stress vulnerability must be studied and optimized. Recently, studies in meagre showed that Trp-enriched diets could mitigate stress responses [
17,
25].
Thus, this study aimed to assess the potential role of both a non-supplement diet and dietary Trp supplementation on growth performance, feed utilization, and metabolic responses in meagre (Argyrosomus regius) juveniles.
2. Materials and Methods
The experiment was conducted at the experimental facilities of Marine Zoological Station, University of Porto, Portugal. Trained scientists (following FELASA category C recommendations) directed the trial, and all procedures were conducted according to the recommendations of the European Union Directive (2010/63/EU) while operating under the Portuguese Law (DL 113/2013) on the protection of animals for scientific purposes. The experimental protocol was approved by the Animal Welfare and Ethics Body committee of the Interdisciplinary Centre of Marine and Environmental Research (ORBEA-CIIMAR, reference ORBEA_CIIMAR_27_2019).
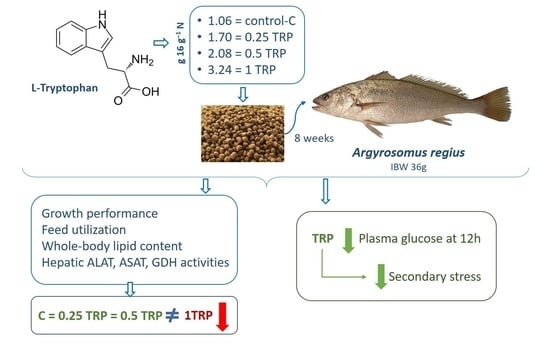
2.1. Experimental Diets
Four experimental diets were formulated to be isoproteic (45%DM) and isolipidic (16%DM), with fish meal and fish oil as main protein and lipid sources, respectively. One diet was not supplemented with L-Trp (control diet), and three other diets were supplemented with graded levels of L-tryptophan (diets 0.25Trp, 0.5Trp and 1Trp, respectively), corresponding to a total dietary tryptophan of 1.06, 1.70, 2.08 and 3.24 g 16 g
−1 N, respectively. L-Tryptophan was coated with agar before being mixed with the other ingredients to avoid leaching and delays in absorption in the digestive tract. Agar was dissolved in boiling distilled water and cooled to 40 °C before mixing. All ingredients were then finely ground, thoroughly mixed, and pelleted using a laboratory pellet mill (CPM: California Pellet, Mill, Crawfordsville, IN, USA) using a 2.5 mm die. Diets were dried in an oven at 35 °C for 24 h and stored at −20 °C until use. The formulation and proximate composition of the diets are presented in
Table 1.
2.2. Growth Trial
Meagre juveniles were obtained from IPMA’s Aquaculture Research Station in Olhão and were kept in quarantine for two weeks. Afterward, the fish were transported to the experimental systems and adapted to the experimental conditions for two weeks. During both periods, the fish were fed a commercial diet (48% protein and 17% lipids, Sorgal, S.A., Ovar, Portugal).
The growth trial was conducted in a thermoregulated semi-recirculating water system, using 12 fiberglass cylindrical tanks of 100 L water capacity, supplied with a continuous flow of filtered seawater. Throughout the trial, water quality parameters were monitored; temperature, salinity, and oxygen levels were checked daily, and nitrogen compounds were measured three times a week. The temperature averaged 22.0 ± 0.5 °C, salinity averaged 35‰, and oxygen levels were kept near saturation.
At the beginning of the trial, fish had an average body weight of 36 ± 2 g. A total of 180 meagre were randomly distributed into 12 homogenous groups of 15 fish. The experimental diets were randomly assigned to triplicate groups of fish. The fish were fed to apparent visual satiation twice a day, six days a week, for 8 weeks. Utmost care was taken to avoid feed losses.
2.3. Sampling
At the beginning and at the end of the trial (56 days later), following one day of feed deprivation the fish were bulk-weighed after being anesthetized with ethylene glycol monophenyl ether (0.3 mL L−1). From the initial stock population, eight fish were collected. At the end of the trial, 3 fish from each tank were also euthanized with lethal doses of anesthetic (10 mL L−1). These fish were then used for whole-body composition analysis and the weights of the whole fish, viscera, and liver were each recorded for hepatosomatic and visceral indices.
Five days after the end of the trial, a time course blood sample was performed. Two fish per tank (6 per dietary treatment) were randomly sampled at 0 h, 1 h, 3 h, 6 h, and 12 h after feeding. Following fish anesthesia, blood samples were taken from the caudal vein with heparinized syringes, and plasma was recovered using centrifugation (1000× g, 10 min) and kept at −20 °C to await further analysis. At the 6 h sampling point, fish were euthanized with a sharp blow to the head and immediately eviscerated in an ice-cold tray. The liver was excised, immediately frozen in liquid nitrogen, and then stored at −80 °C, until measurement of amino acid catabolism.
2.4. Analytical Methods
2.4.1. Proximate Analysis
Chemical analysis of the ingredients, diets, and whole body composition was conducted in duplicate following standard methodology [
26]: the dry matter obtained by drying the samples at 105 °C until a constant weight; protein content (N × 6.25) using the Kjeldahl method following acid digestion and distillation (Tecator System, Högamäs, Sweden; extraction unit model 1015 and 1026, respectively); lipid content extracted with petroleum ether using a Soxtec system (Tecator Systems, Höganäs, Sweden; extraction unit model 1043 and service until model 1046); ash acquired by means of incineration in a muffle furnace at 450 °C for 16 h.
The amino acid content of the experimental diets was analyzed in duplicate using high-performance liquid chromatography (HPLC) according to the Pico–Tag method. Samples were hydrolyzed for 23 h with 6N hydrochloric acid at 110 °C under an N
2 atmosphere, and derivatized with phenylisothiocyanate reagent (PITC, Ref. WAT088120, Waters™, Milford, MA, USA) before being separated using gradient exchange chromatography at 46 °C (Waters auto sample model 717 plus, Waters binary pump model 1525, Waters dual absorbance detector model 2487), according to the Pico–Tag method. Norleucine was used as an internal standard. Chromatographic peaks were identified, integrated, and quantified by comparing them to a known amino acid standard, using the Waters Breeze software package (Pierce NC10180, Waters™, Milford, MA, USA, 2001). Tryptophan was measured with a spectrophotometric method, as described by De Vries et al. [
27]. Amino acid proximate composition is shown in
Table 2.
2.4.2. Plasma Metabolites
Commercial kits from Spinreact, S.A. (Gerona, Spain) were used to determine plasma cholesterol (ref. 1001090), glucose (ref. 1001190), and triglycerides (ref. 41031). Plasma protein was determined according to Bradford [
28], using bovine serum albumin as standard.
2.4.3. Enzyme Activity
Alanine aminotransferase (ALAT; EC 2.6.1.2), aspartate aminotransferase (ASAT; EC 2.6.1.1.), and glutamate dehydrogenase (GDH; EC 1.4.1.2.) activities were assessed in the liver. The livers were homogenized (dilution1/100) in an ice-cold buffer (30 mM HEPES, 0.25 mM saccharose, 0.5 mM EDTA, 5 mM K
2HPO
4, 1 mM dithiothreitol, pH 7.4). After 900×
g for 10 min centrifugation, the resultant supernatant was sonicated for 1 min (pulse 1 s, amplitude 50) and centrifuged again at 15,000×
g for 20 min. The resultant supernatant was separate for ALAT, ASAT, and GDH activity measurements. The GDH activity was measured using 10 mM of L-glutamic acid, as described previously by Bergmeyer (1974), at 37 °C. ALAT and ASAT were assayed with kits from Enzyline (ALAT/GPT, ref. 63313; ASAT/GOT, ref. 63213, bioMérieux, S.A., France) at 37 °C and followed at 340 nm. Enzyme activity was expressed as nmol of substrate transformed per min and per mg of protein under the assay conditions. Soluble protein concentration was determined using the Bradford method [
27], with bovine serum albumin used as a standard.
2.5. Statistical Analysis
Data were checked for normal distribution and homogeneity of variances, and normalized, if necessary. Statistical data analysis was conducted using a one-way analysis of variance (one-way ANOVA) at a probability level of p ≤ 0.05. Significant differences among mean values were determined using the Tukey multiple comparison test. To analyze plasma metabolites, a two-way analysis of variance was used (two-way ANOVA), with diets and time as fixed factors. When the interaction was significant, diet and time effects were analyzed by performing a one-way ANOVA. All statistical analyses were performed using SPSS version 28.0 for Windows software package (IBM, Armonk, NY, USA, 2022).
4. Discussion
Many practices deemed necessary to aquaculture unavoidably submit fish to several stressors. In response to a stressor, fish undergo several biochemical and physiological changes to compensate for the imposed challenge, which can impair the fish’s nutritional and health status [
29]. Tryptophan (Trp) has been pointed out as a possible nutritional strategy to mitigate stress through use as a feed additive, due to being the precursor of serotonin and melatonin [
10,
30,
31,
32,
33,
34]. Previous studies in meagre showed that dietary supplementation with Trp (total dietary level of 0.11% diet) modulated their immune parameters following acute stress [
3]. Moreover, dietary supplementation with 0.25% L-Trp did not affect meagre growth performance and enhanced their observed mucus antibacterial activity [
18] Correspondingly, this study evaluated the effect of a higher Trp dietary supplementation (0.25, 0.5 and 1% of the diet) on meagre performance and feed utilization, as well as on the plasma metabolites profile and the key liver enzymes involved in amino acid catabolism.
Tryptophan requirements differ for each fish species and have been reported to range from 0.3 to 1.3% of dietary protein [
34,
35]. Moreover, these requirements may increase with stress [
34]. To our knowledge, the nutritional requirements of Trp for meagre have yet to be established. In some species, an excess of dietary Trp (i.e., above their requirement levels) may increase serotonin and melatonin production [
30,
32], which can lead to a decrease in voluntary feed intake [
36,
37]. Murthy and Vanghese [
38] determined that for
Labeo rohita the optimum Trp level was 1.13%, and fish fed a diet comprising 1.38% Trp presented a decreased level of growth; the authors attributed this to a possible toxic effect, which will be further discussed. Hseu et al. [
8] also showed that supplementation of Trp altered feeding behavior, with a decrease in feed intake observed with supplementation level of Trp higher than 0.5% of the diet. Recent studies have also suggested that growth performance may be reduced by higher dietary Trp levels [
39,
40,
41]. Herrera et al. [
18] showed that supplementation up to 0.25% of the total diet did not affect growth performance nor feed efficiency in meagre. In this study, dietary supplementation levels of Trp up to 0.5% (diet 0.5Trp; 2.08 g 16 g
−1 N total dietary Trp level) did not affect growth performance and feed utilization. However, 1% Trp supplementation (diet 1Trp; 3.24 g 16 g
−1 N total dietary Trp level) decreased both these parameters, suggesting that a total Trp dietary level of 3.24 g 16 g
−1 N may be harmful to this species. As aforementioned, high levels of some amino acids could be considered toxic. This may be due to excessive levels exerting antagonistic actions against other amino acids, i.e., may impair the absorption and utilization of other amino acids, resulting in a deficient absorption rate of this amino acid. Trp is a large neutral amino acid (LNAA), competing for the same transporter as other LNAAs, such as valine, leucine, isoleucine, tyrosine, and phenylalanine [
42]. Supplementation of Trp can affect the Trp/LNAA ratio, meaning that Trp brain concentrations may increase at the expense of other LNAAs [
20,
32,
43,
44].
The whole-body lipid composition of meagre decreased in the fish group feeding on 1Trp, while whole-body protein levels were maintained regardless of dietary treatment. The supplied diets from diet 0Trp to 0.5Trp were enough both for energy expenditure and lipid deposition. However, fish fed the 1Trp diet had lower ingestion, so less energy was available for energetic metabolism, decreasing the amount allocated to lipid deposition. Similar results were reported for rats, with high supplementation of Trp reducing fat accumulation in the body, which could be to higher oxidation of fatty acids [
45].
In addition to measuring cortisol and catecholamine levels, secondary stress response parameters (such as plasma glucose levels) have been used [
46,
47,
48] to evaluate the stress response. Under stress, the release of cortisol, catecholamines, and glucagon hormones increases the metabolic state, and glucose functions as the main energy source to cope with this increased energy demand [
49]. In fish, increased plasma glucose levels are often associated with increased plasma cortisol levels [
50]. This relationship between Trp and cortisol regulation has been attributed to the interaction between serotonergic activity and the hypothalamic–pituitary–interrenal axis. Therefore, glucose levels are expected to increase upon stress [
49]. In fact, reports of Trp-enriched diets fed to fish showed that stress markers such as cortisol and glucose showed similar tendencies, with increased levels of Trp in the diets ultimately decreasing both serum cortisol and blood glucose in data for diets supplemented with Trp [
31,
51]. In the present study, the supplementation of Trp lowered glucose levels 3 h after feeding compared to 0 h after feeding. These results evidence a probable reduction of the stress condition of fish that were fed the Trp-supplemented diets, reflecting a reduction in body energy mobilization. Similarly, feeding
Cyprinus carpio a diet supplemented with 5% Trp has been shown to result in lower cortisol and glucose levels compared to a control group after copper exposure, yet levels of cortisol and glucose did not reach pre-exposure levels [
9]. In
Cirrhinus mrigala, plasma cortisol and blood levels were affected by dietary treatment up to 2.72% DM Trp supplementation, in a similar observable trend; the supplementation of L-Trp gradually decreasing both cortisol and glucose levels [
31]. In rohu, cortisol and glucose levels decreased with the increase of dietary Trp level from 0% up to 1.42% of their total diet [
51]. In gilthead seabream, supplemented diets of 1.5 and 2% DM (total dietary Trp of 0.54 and 0.78% DM) decreased glucose levels in line with increasing Trp levels, a result observed in both stressed and unstressed fish [
52]. In rainbow trout exposed to stress, the Trp supplemented diet (at 3.57% of the total dietary level) reduced cortisol levels compared to the control diet. Interestingly, in the same study, fish not submitted to stress showed higher cortisol levels when fed Trp supplemented diets [
17]. The authors attributed this evaluation to other underlying mechanisms involved in mediating the roles of Trp on stress response [
16]. In addition, the rainbow trout showed a decrease in post-stress plasma cortisol when fed a Trp supplemented diet (total dietary Trp level of 3.57%) for 7 days [
53]. In Atlantic salmon, increasing dietary Trp levels by 3-fold (1.2% Trp) resulted in decreased plasma cortisol levels compared with those fed a 2-fold and one-fold increase [
54].
Stress conditions induce a hypermetabolic status which may induce the mobilization of amino acids as a coping mechanism for the increased energy demand. Normally, increased activity of GDH, ASAT, and ALAT are used as indicators of increased catabolic utilization of dietary protein by fish, as the amino acids which are not used for protein synthesis are deaminated [
55]. In the present study, hepatic amino acid catabolism enzyme activity was affected by dietary treatment, decreasing with the increased level of Trp inclusion in the diets. These results are in line with those reported in meagre by Herrera et al. [
17], who observed a decreased activity of amino acid catabolic enzymes with the augmentation of Trp levels. However, other authors have reported the opposite. For example, in Atlantic cod (
Gadus morhua) and Senegalese sole (
Solea senegalensis), it has been shown that the dietary supplementation of Trp increased the activity of these enzymes [
56,
57], as the dietary excess of amino acids induced the activity of protein-metabolizing enzymes, particularly GDH and ALAT [
58].
As aforementioned, nitrogen retention was lower in fish fed diets of 1% Trp, while the level of protein catabolism activity decreased. Thus, it can be hypothesized that the reduction of nitrogen retention in fish fed the 1Trp diet was not due to increased use of protein for energetic purposes but rather to a deficient protein intake associated with a reduced feed intake. Our results correlate well with the reduction of amino acid catabolism enzymes, which may indicate a reduction in the meagres’ metabolic rate, reducing energy expenditure and with it the use of amino acids for energetic purposes. The discrepancies between these results and those of the different studies found in the literature could be due to several aspects, such as species, the nutritional status of the animals, levels of supplementation, developmental stage, and stress condition.

 ,
, 

{kind=link}