
Established and Emerging Methods for Protecting Linear DNA in Cell-Free Expression Systems

Abstract
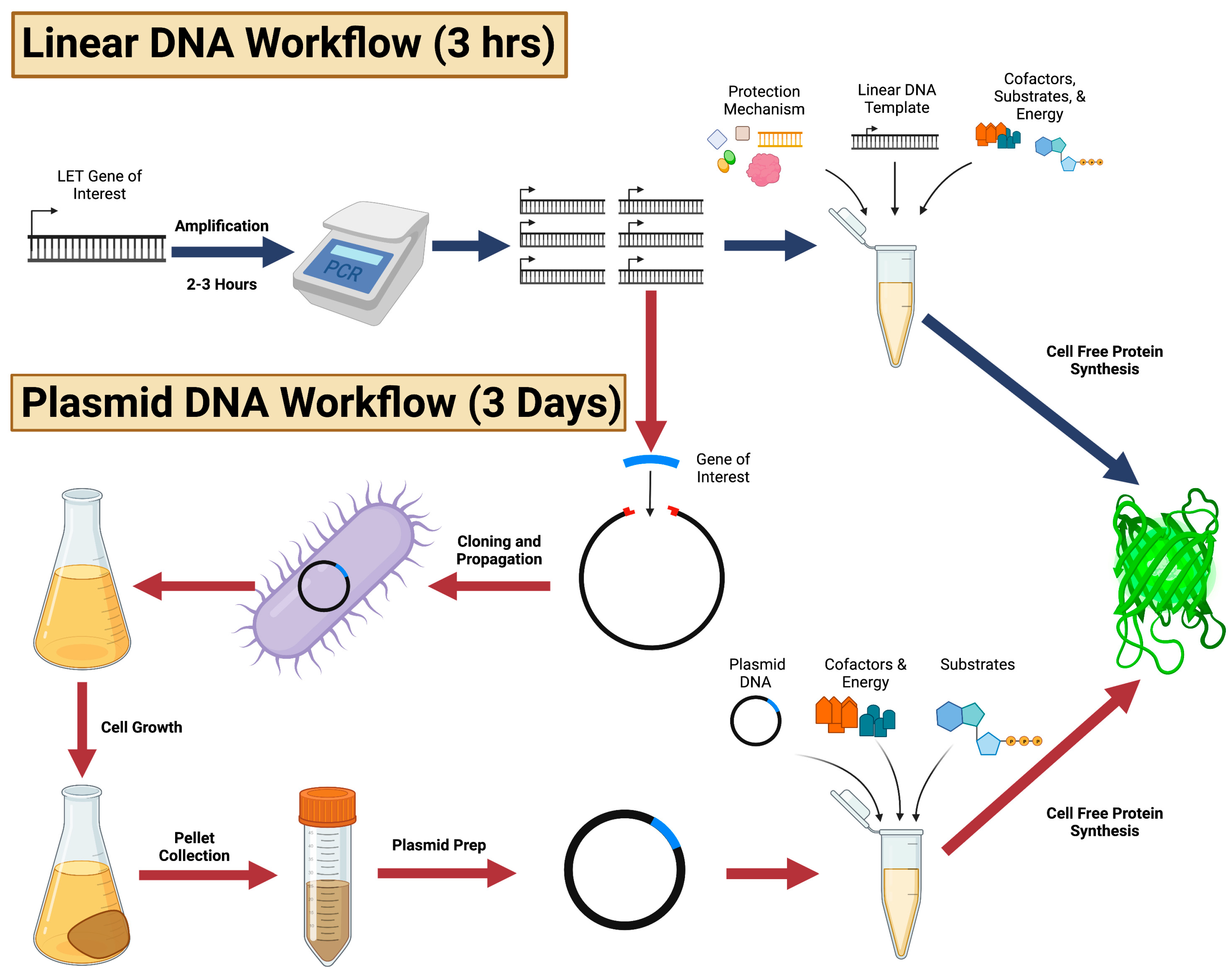
:1. Introduction
2. Established Methods
2.1. Nuclease Inhibition
2.1.1. GamS
2.1.2. Chi Sites
2.1.3. Ku
2.1.4. Small Molecule Inhibitors

{kind=link}
{kind=link}
| Inhibitor | Mechanism | Expression Data | Optimal Concentration | Sources |
|---|---|---|---|---|
| Control (Standard Linear Template) | N/A | Complete degradation of templates | N/A | [4] |
| GamS | Competitive Inhibitor | 650 AU of the Broccoli Aptamer -Produced 5 uM deGFP | 3.5–4 μM per reaction flask for a standard CFPS reaction | [4,13] |
| Chi Sites | Competitive DNA Binding Site | 600 AU of the Broccoli Aptamer 9 μM of deGFP a 14 h reaction produced 50 μM of GFP | The most ideal concentration of the Chi Site inhibitors is 0.5 uM of the Chi6 segment (85 bp) | [4,19] |
| Ku | Competitive Inhibitor | Ku barely reached 200 AU Highest effectiveness in V. Natriegens | 1–4 μM of protein is ideal | [4] |
| Tus-Ter | Initiation– Termination System | 52,000 RFU of deGFP 40,000 RFU of mCherry | 5 μM of Tus-Ter per CFPS reaction | [32] |
| CID 697851 | Small Molecule Inhibitor | Increased yields by 200% | 3.3 uM | [30,31] |
| CID 1517823 | Small Molecule Inhibitor | Increased yields by 300% | 5.1 uM | [30,31] |
2.2. Untranslated Regions
2.3. Tus-Ter Initiation/Termination System
2.4. Strain Engineering
| Alteration to Gene | Growth Effects | Degradation Time | Expression Yields | References |
|---|---|---|---|---|
| Wild Type Strain (No Modificaitons) | Normal Growth | Fully degraded 100% in 3 Hrs | 220 ug of protein | [33] |
| recBCD Deletion | Severely Limited Growth | N/A | N/A | [33] |
| recBCD Replacement | Normal Growth | Only degraded by 33% in 3 Hrs | 700 ug of protein | [33] |
3. Emerging Methods
3.1. Paranemic Crossover DNA
| Method | Type of Protection | Sequence | References |
|---|---|---|---|
| Chi Sites | Competitive Inhibitor | 5′-GCTGGTGG-3′ | [19] |
| Ter | Initiation Termination System | 5′ -AATTAGTATGTTGTAACTAAAGT-3′ | [32,35,36,37] |
| PX DNA | DNA Complex | PX1: 5′- GTGGTATCATCAATGCTATGTGTAGGCTTAGACCTGAG-3′ PX2: 5′- ACTAGGTCGCAACAGACACAATACTTGACCGAATCACT-3′ PX3: 5′- AGTGAGTCTAACAAGTCACATATCTGTGATGATCTAGT-3′ PX4: 5′- CTCAGTTCGGTGCCTAATTGTGGCATTTGCGACACCAC-3′ | [42,43,44] |
3.2. Chemical Modifications of DNA
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brookwell, A.; Oza, J.P.; Caschera, F. Biotechnology Applications of Cell-Free Expression Systems. Life 2021, 11, 1367. [Google Scholar] [CrossRef] [PubMed]
- Silverman, A.D.; Karim, A.S.; Jewett, M.C. Cell-free gene expression: An expanded repertoire of applications. Nat. Rev. Genet. 2020, 21, 151–170. [Google Scholar] [CrossRef]
- Gregorio, N.E.; Levine, M.Z.; Oza, J.P. A User’s Guide to Cell-Free Protein Synthesis. Methods Protoc. 2019, 2, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, S.S.; Johns, N.I.; Noireaux, V.; Wang, H.H. Protecting Linear DNA Templates in Cell-Free Expression Systems from Diverse Bacteria. ACS Synth. Biol. 2020, 9, 2851–2855. [Google Scholar] [CrossRef]
- Sato, W.; Sharon, J.; Deich, C.; Gaut, N.; Cash, B.; Engelhart, A.E.; Adamala, K.P. Akaby—Cell-free protein expression system for linear templates. PLoS ONE 2022, 17, e0266272. [Google Scholar] [CrossRef]
- Romantseva, E.; Alperovich, N.; Ross, D.; Lund, S.P.; A Strychalski, E. Effects of DNA Template Preparation on Variability in Cell-Free Protein Production. Synth. Biol. 2022, 7, ysac015. [Google Scholar] [CrossRef] [PubMed]
- Schinn, S.-M.; Broadbent, A.; Bradley, W.T.; Bundy, B.C. Protein synthesis directly from PCR: Progress and applications of cell-free protein synthesis with linear DNA. New Biotechnol. 2016, 33, 480–487. [Google Scholar] [CrossRef] [Green Version]
- McSweeney, M.A.; Styczynski, M.P. Effective Use of Linear DNA in Cell-Free Expression Systems. Front. Bioeng. Biotechnol. 2021, 9, 715328. [Google Scholar] [CrossRef]
- Stein, V.; Hollfelder, F. An efficient method to assemble linear DNA templates for in vitro screening and selection systems. Nucleic Acids Res. 2009, 37, e122. [Google Scholar] [CrossRef]
- Sitaraman, K.; Esposito, D.; Klarmann, G.; Le Grice, S.F.; Hartley, J.L.; Chatterjee, D.K. A novel cell-free protein synthesis system. J. Biotechnol. 2004, 110, 257–263. [Google Scholar] [CrossRef]
- Fujiwara, K.; Doi, N. Biochemical Preparation of Cell Extract for Cell-Free Protein Synthesis without Physical Disruption. PLoS ONE 2016, 11, e0154614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederholtmeyer, H.; Sun, Z.Z.; Hori, Y.; Yeung, E.; Verpoorte, A.; Murray, R.M.; Maerkl, S.J. Rapid cell-free forward engineering of novel genetic ring oscillators. eLife 2015, 4, e09771. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Z.; Yeung, E.; Hayes, C.A.; Noireaux, V.; Murray, R.M. Linear DNA for Rapid Prototyping of Synthetic Biological Circuits in an Escherichia coli Based TX-TL Cell-Free System. ACS Synth. Biol. 2014, 3, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Khambhati, K.; Bhattacharjee, G.; Gohil, N.; Braddick, D.; Kulkarni, V.; Singh, V. Exploring the Potential of Cell-Free Protein Synthesis for Extending the Abilities of Biological Systems. Front. Bioeng. Biotechnol. 2019, 7, 248. [Google Scholar] [CrossRef]
- Montagud-Martinez, R.; Marquez-Costa, R.; Rodrigo, G. Programmable regulation of translation by harnessing the CRISPR-Cas13 system. Chem. Commun. 2023, 59, 2616–2619. [Google Scholar] [CrossRef]
- Casjens, S.R.; Hendrix, R.W. Bacteriophage lambda: Early pioneer and still relevant. Virology 2015, 479, 310–330. [Google Scholar] [CrossRef] [Green Version]
- Salaj-Smic, E.; Marsić, N.; Trgovcević, Z.; Lloyd, R.G. Modulation of EcoKI restriction in vivo: Role of the lambda Gam protein and plasmid metabolism. J. Bacteriol. 1997, 179, 1852–1856. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, M.; Troman, L.; Ismah, W.A.W.N.; Chaban, Y.; Avison, M.B.; Dillingham, M.S.; Wigley, D.B. Structural basis for the inhibition of RecBCD by Gam and its synergistic antibacterial effect with quinolones. eLife 2016, 5, e22963. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.; Maxwell, C.S.; Collins, S.P.; Beisel, C.L.; Noireaux, V. Short DNA containing χ sites enhances DNA stability and gene expression in E. coli cell-free transcription–translation systems. Biotechnol. Bioeng. 2017, 114, 2137–2141. [Google Scholar] [CrossRef]
- Li, X.; Heyer, W.-D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008, 18, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Lovett, S.T. The DNA Exonucleases of Escherichia coli. EcoSal Plus 2011, 4. [Google Scholar] [CrossRef] [Green Version]
- Spies, M.; Amitani, I.; Baskin, R.J.; Kowalczykowski, S.C. RecBCD Enzyme Switches Lead Motor Subunits in Response to χ Recognition. Cell 2007, 131, 694–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.H.Y.; Pannunzio, N.R.; Adachi, N.; Lieber, M.R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nat. Rev. Mol. Cell Biol. 2017, 18, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Mansilla-Soto, J.; Cortes, P. VDJ Recombination. J. Exp. Med. 2003, 197, 543–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Guo, H.; Wang, D.; Tu, R.; Qing, G.; Liu, H. Genome-Wide Analysis Identifies Rag1 and Rag2 as Novel Notch1 Transcriptional Targets in Thymocytes. Front. Cell Dev. Biol. 2021, 9, 703338. [Google Scholar] [CrossRef] [PubMed]
- Fell, V.L.; Schild-Poulter, C. The Ku heterodimer: Function in DNA repair and beyond. Mutat. Res. Rev. Mutat. Res. 2015, 763, 15–29. [Google Scholar] [CrossRef]
- Smith, P.E.J.; Slouka, T.; Dabbas, M.; Oza, J.P. From Cells to Cell-Free Protein Synthesis within 24 Hours Using Cell-Free Autoinduction Workflow. J. Vis. Exp. 2021, 23, e62866. [Google Scholar] [CrossRef]
- Levine, M.Z.; So, B.; Mullin, A.C.; Fanter, R.; Dillard, K.; Watts, K.R.; La Frano, M.R.; Oza, J.P. Activation of Energy Metabolism through Growth Media Reformulation Enables a 24-Hour Workflow for Cell-Free Expression. ACS Synth. Biol. 2020, 9, 2765–2774. [Google Scholar] [CrossRef]
- Burrington, L.R.; Watts, K.R.; Oza, J.P. Characterizing and Improving Reaction Times for E. coli-Based Cell-Free Protein Synthesis. ACS Synth. Biol. 2021, 10, 1821–1829. [Google Scholar] [CrossRef]
- Amundsen, S.K.; Spicer, T.; Karabulut, A.C.; Londoño, L.M.; Eberhart, C.; Vega, V.F.; Bannister, T.D.; Hodder, P.; Smith, G.R. Small-Molecule Inhibitors of Bacterial AddAB and RecBCD Helicase-Nuclease DNA Repair Enzymes. ACS Chem. Biol. 2012, 7, 879–891. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, P.; Smith, M.T.; Bundy, B.C. Cell-free unnatural amino acid incorporation with alternative energy systems and linear expression templates. New Biotechnol. 2014, 31, 28–34. [Google Scholar] [CrossRef]
- Norouzi, M.; Panfilov, S.; Pardee, K. High-Efficiency Protection of Linear DNA in Cell-Free Extracts from Escherichia coli and Vibrio natriegens. ACS Synth. Biol. 2021, 10, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, Y. In silico Design of Linear DNA for Robust Cell-Free Gene Expression. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, D.B.; Decornez, H.; Furr, J.R.; Bajorath, J. Docking and scoring in virtual screening for drug discovery: Methods and applications. Nat. Rev. Drug Discov. 2004, 3, 935–949. [Google Scholar] [CrossRef] [PubMed]
- Berghuis, B.A.; Raducanu, V.-S.; Elshenawy, M.; Jergic, S.; Depken, M.; Dixon, N.E.; Hamdan, S.M.; Dekker, N.H. What is all this fuss about Tus? Comparison of recent findings from biophysical and biochemical experiments. Crit. Rev. Biochem. Mol. Biol. 2017, 53, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Guajardo, R.; Sousa, R. Characterization of the effects of Escherichia coli replication terminator protein (Tus) on transcription reveals dynamic nature of the Tus block to transcription complex progression. Nucleic Acids Res. 1999, 27, 2814–2824. [Google Scholar] [CrossRef] [Green Version]
- Neylon, C.; Kralicek, A.V.; Hill, T.M.; Dixon, N.E. Replication Termination in Escherichia coli: Structure and Antihelicase Activity of the Tus-Ter Complex. Microbiol. Mol. Biol. Rev. 2005, 69, 501–526. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Ellis, H.M.; Lee, E.-C.; Jenkins, N.A.; Copeland, N.G.; Court, D.L. An efficient recombination system for chromosome engineering in Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 5978–5983. [Google Scholar] [CrossRef] [Green Version]
- Michel-Reydellet, N.; Woodrow, K.; Swartz, J. Increasing PCR Fragment Stability and Protein Yields in a Cell-Free System with Genetically Modified Escherichia coli Extracts. Microb. Physiol. 2005, 9, 26–34. [Google Scholar] [CrossRef]
- Murphy, K.C. Lambda Gam protein inhibits the helicase and chi-stimulated recombination activities of Escherichia coli RecBCD enzyme. J. Bacteriol. 1991, 173, 5808–5821. [Google Scholar] [CrossRef] [Green Version]
- Batista, A.C.; Levrier, A.; Soudier, P.; Voyvodic, P.L.; Achmedov, T.; Reif-Trauttmansdorff, T.; DeVisch, A.; Cohen-Gonsaud, M.; Faulon, J.-L.; Beisel, C.L.; et al. Differentially Optimized Cell-Free Buffer Enables Robust Expression from Unprotected Linear DNA in Exonuclease-Deficient Extracts. ACS Synth. Biol. 2022, 11, 732–746. [Google Scholar] [CrossRef] [PubMed]
- Kizer, M.; Huntress, I.D.; Walcott, B.D.; Fraser, K.; Bystroff, C.; Wang, X. Complex between a Multicrossover DNA Nanostructure, PX-DNA, and T7 Endonuclease I. Biochemistry 2019, 58, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Jiang, H.; Sha, R.; Canary, J.W.; Seeman, N.C. The unusual and dynamic character of PX-DNA. Nucleic Acids Res. 2015, 43, 7201–7206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Yan, H.; Wang, T.; Seeman, N.C. Paranemic Crossover DNA: A Generalized Holliday Structure with Applications in Nanotechnology. J. Am. Chem. Soc. 2004, 126, 1666–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraniak, D.; Boryski, J. Triazole-Modified Nucleic Acids for the Application in Bioorganic and Medicinal Chemistry. Biomedicines 2021, 9, 628. [Google Scholar] [CrossRef]
- Lin, C.; Ke, Y.; Li, Z.; Wang, J.H.; Liu, Y.; Yan, H. Mirror Image DNA Nanostructures for Chiral Supramolecular Assemblies. Nano Lett. 2009, 9, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Hauser, N.C.; Martinez, R.; Jacob, A.; Rupp, S.; Hoheisel, J.D.; Matysiak, S. Utilising the left-helical conformation of L-DNA for analysing different marker types on a single universal microarray platform. Nucleic Acids Res. 2006, 34, 5101–5111. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.; Sczepanski, J.T. Direct Comparison of d-DNA and l-DNA Strand-Displacement Reactions in Living Mammalian Cells. ACS Synth. Biol. 2021, 10, 209–212. [Google Scholar] [CrossRef]
- Kurreck, J. Design of antisense oligonucleotides stabilized by locked nucleic acids. Nucleic Acids Res. 2002, 30, 1911–1918. [Google Scholar] [CrossRef] [Green Version]
- Marušič, M.; Veedu, R.N.; Wengel, J.; Plavec, J. G-rich VEGF aptamer with locked and unlocked nucleic acid modifications exhibits a unique G-quadruplex fold. Nucleic Acids Res. 2013, 41, 9524–9536. [Google Scholar] [CrossRef]
- Oliynyk, R.T.; Church, G.M. Efficient modification and preparation of circular DNA for expression in cell culture. Commun. Biol. 2022, 5, 1393. Available online: https://www.nature.com/articles/s42003-022-04363-z#citeas (accessed on 16 March 2023). [CrossRef] [PubMed]
- Chen, X.; Lu, Y. Circular RNA: Biosynthesis in vitro. Front. Bioeng. Biotechnol. 2021, 9, 787881. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, S.K.; Belk, J.A.; Amaya, L.; Li, Z.; Cardenas, A.; Abe, B.T.; Chen, C.-K.; Wender, P.A.; Chang, H.Y. Engineering circular RNA for enhanced protein production. Nat. Biotechnol. 2022, 41, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Wesselhoeft, R.A.; Kowalski, P.S.; Anderson, D.G. Engineering circular RNA for potent and stable translation in eukaryotic cells. Nat. Commun. 2018, 9, 2629. [Google Scholar] [CrossRef] [Green Version]
- BioSpace. Cell-Free Protein Expression Market Size, Share, Report 2022 to 2030. Available online: https://www.biospace.com/article/cell-free-protein-expression-market-size-share-report-2022-to-2030/?keywords=agriculture (accessed on 16 March 2023).
- The White House. Executive Order on Advancing Biotechnology and Biomanufacturing Innovation for a Sustainable, Safe, and Secure American Bioeconomy. Available online: https://www.whitehouse.gov/briefing-room/presidential-actions/2022/09/12/executive-order-on-advancing-biotechnology-and-biomanufacturing-innovation-for-a-sustainable-safe-and-secure-american-bioeconomy/ (accessed on 16 March 2023).


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fochtman, T.J.; Oza, J.P. Established and Emerging Methods for Protecting Linear DNA in Cell-Free Expression Systems. Methods Protoc. 2023, 6, 36. https://doi.org/10.3390/mps6020036
Fochtman TJ, Oza JP. Established and Emerging Methods for Protecting Linear DNA in Cell-Free Expression Systems. Methods and Protocols. 2023; 6(2):36. https://doi.org/10.3390/mps6020036
Chicago/Turabian StyleFochtman, Trevor J., and Javin P. Oza. 2023. "Established and Emerging Methods for Protecting Linear DNA in Cell-Free Expression Systems" Methods and Protocols 6, no. 2: 36. https://doi.org/10.3390/mps6020036