
There’s Plenty of Room Right Here: Biological Systems as Evolved, Overloaded, Multi-Scale Machines

Abstract
:1. Introduction
1.1. What Constitutes a Computer?
1.2. Observer Dependency
1.3. What Things Are Alive?
1.4. From a Philosophy to a Science of How Life (Poly)Computes
1.5. Why “Life as Computation” Matters
2. Current Debates: Dissolving Dichotomous Thinking
2.1. Structure Function Mapping and Polycomputing

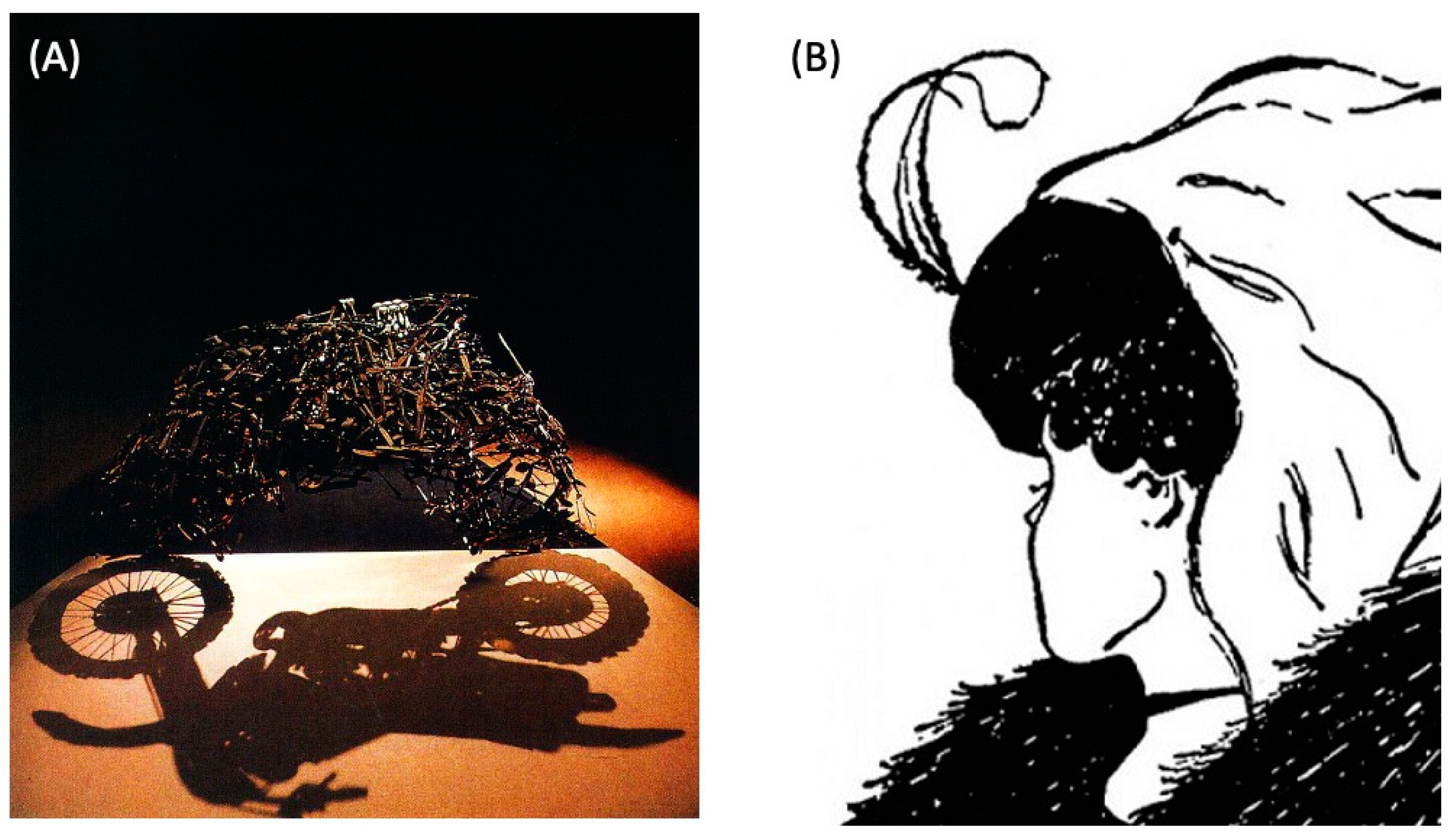{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assumed Distinction | Counterexamples |
|---|---|
| Software/Hardware | Physical materials that compute [65] and learn [77]. |
| Tape/Machine | Tape-less von Neumann self replicators [78] (assuming a Turing machine architecture) |
| Digital/Analog | Evolved digital circuits can exploit electromagnetic properties of the circuit’s substrate [11]. |
| Machine/Life form | AI-designed organisms [78,79]. |
| Automaton/Free agent | The intentional stance [26]. |
| Brain/Body | Computational metamaterials [8]. |
| Body/Environment | Other cells are the environment for a cell in a multicellular body. |
| Intelligent/Faking it | AI technologies that seem to pass verbal [80], visual [81], or physical [82] Turing tests. |
| Made/Evolved | Artefacts designed by human-created evolutionary algorithms. |
2.2. Dichotomous Thinking in the Life Sciences
2.3. Dichotomous Thinking in Computer Science
2.4. Polycomputing in Bodies and Brains
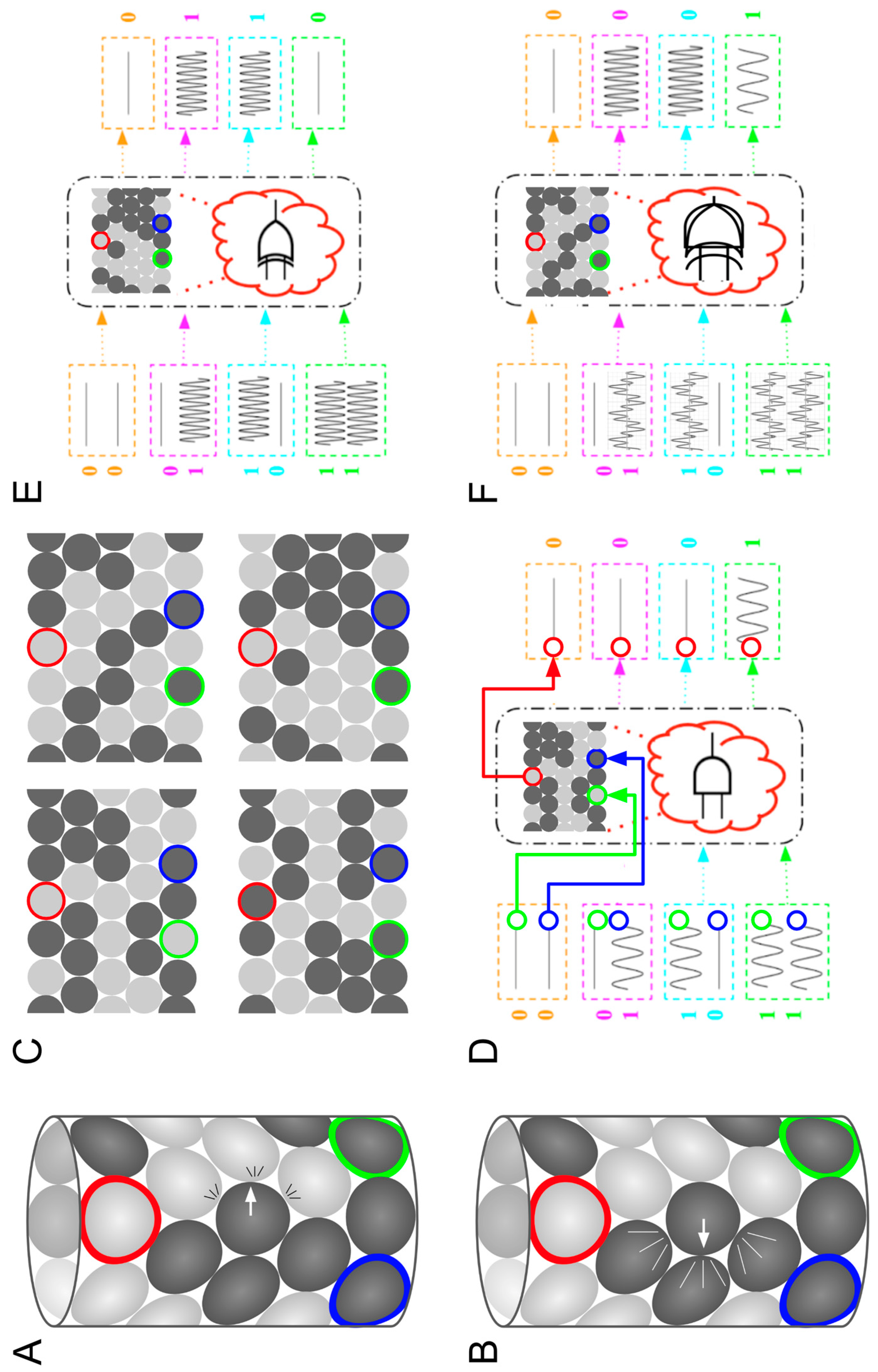
3. Learning from Superposed Systems in Engineering
4. Biology Is Massively Overloaded: Polycomputing
| Multiple Computations in the Same Biological Hardware | Reference |
|---|---|
| Mitochondria also act as micro-lenses in photoreceptors | [164] |
| Proteins acting in multiple (fluctuating) conformations | [165] |
| Pathways and transcriptional networks regulating real-time physiology and performing learning at the same time | [161,166,167,168,169,170,171,172] |
| Gene regulatory networks with multiple memories/behaviors | [170,172,173,174] |
| Chemical networks performing neural network tasks | [171,175] |
| RNA encoding enzyme and protein functions | [176,177,178,179] |
| ATP as an energy source and neurotransmitter | [180] |
| DNA with more than one active reading frame (overlapping/dual-coding genes) | [181,182] |
| Ion channels that are also transcription factors | [183] |
| DNA transcription factors working in DNA replication machinery | [184] |
| Polysemanticity and superposition of meaning in neural networks and language understanding | [185,186,187] |
| Cytoskeleton performing computations via simultaneous biomechanical, bioelectrical, and quantum-mechanical dynamics | [188,189,190,191,192,193,194,195,196,197] |
| Electrophysiological networks performing memory functions while regulating heartbeat | [198,199,200] |
| Bioelectric networks performing physiological functions while also regulating morphogenesis | [117] |
| Spider webs as auditory sensors and structural elements | [3] |
| Pleiotropy: most genes have multiple functions | [74] |
| Holographic memory in the brain | [201] |
| Multiple behaviors in the same neuronal circuit | [202] |
| Multiple personalities in the same brain (dissociative identity disorder and split brain studies) | [203,204] |
| Calcium dynamics performing as a hub in a huge bowtie network of diverse simultaneous processes | [205,206] |
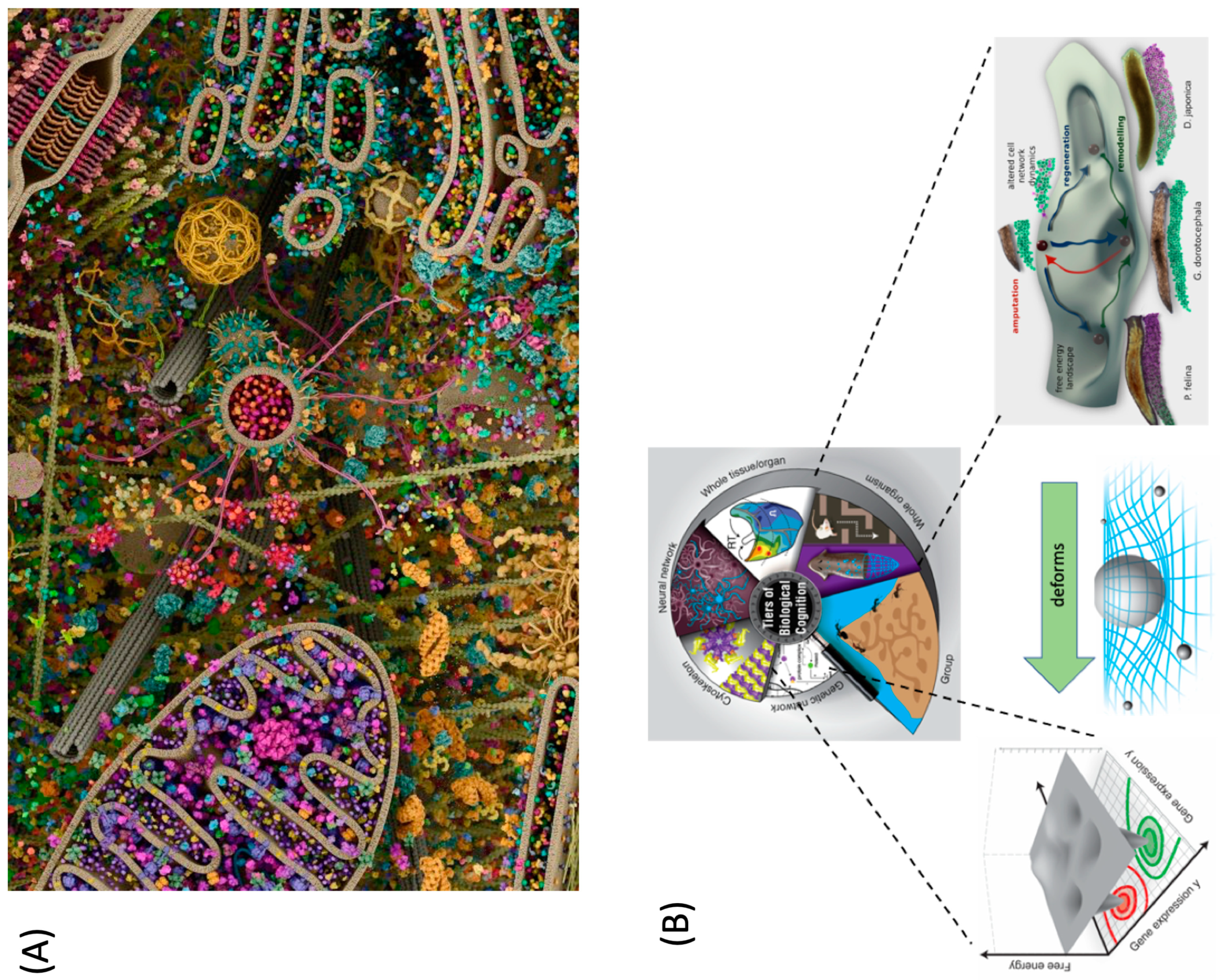
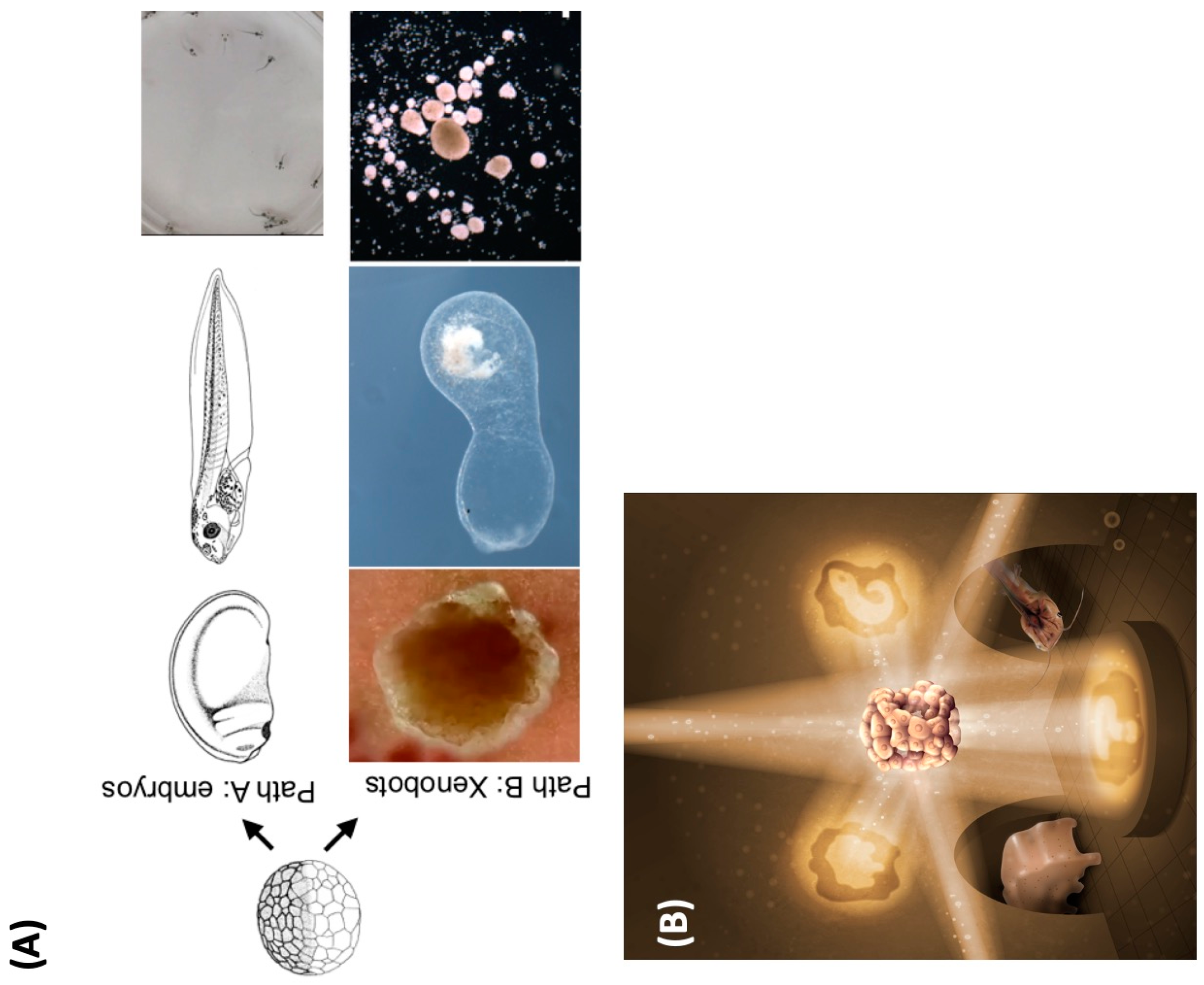
4.1. Evolutionary Pivots: Origins of Polycomputing?
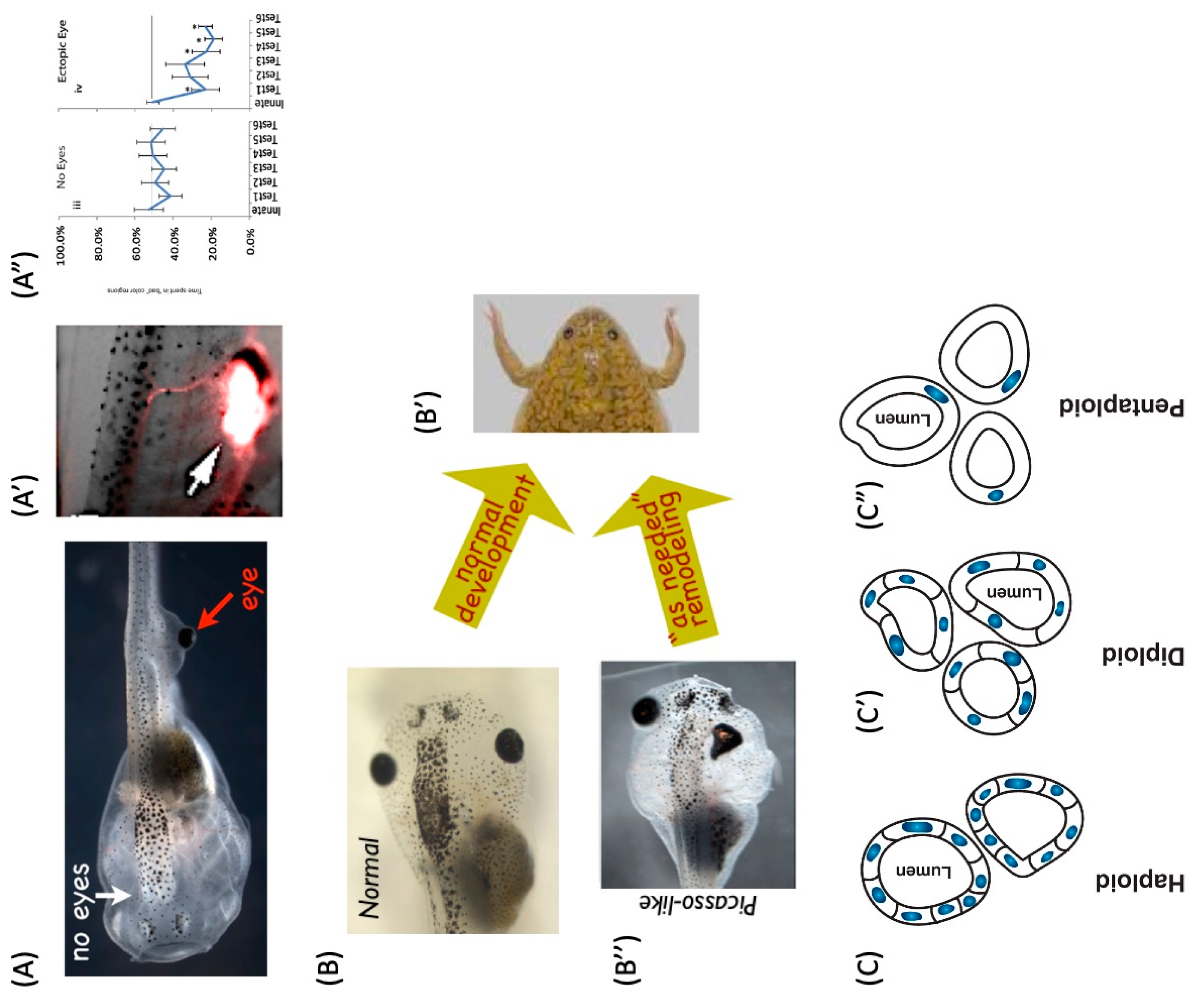
4.2. Polycomputing and the Range of Phenotypic Possibility

4.3. Evolving Polycomputing
4.4. A New Approach to Identifying and Harnessing Computational Capabilities In Vivo and In Silico
5. Conceptual Transitions
5.1. Directions of Conceptual Travel
5.2. Practical Implications for AI/Robotics
6. Gradual Computing in Biology: When Does the (Digital) Soul Enter the (Analog) Body?
6.1. Agency and Persuadability: Implication for Polycomputing
6.2. The Impact of Observer Frames
6.3. Becoming a Computer
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Feynman, R. There’s plenty of room at the bottom. Eng. Sci. 1960, 23, 22–36. [Google Scholar]
- Levin, M. Technological Approach to Mind Everywhere: An Experimentally-Grounded Framework for Understanding Diverse Bodies and Minds. Front. Syst. Neurosci. 2022, 16, 768201. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lai, J.; Menda, G.; Stafstrom, J.A.; Miles, C.I.; Hoy, R.R.; Miles, R.N. Outsourced hearing in an orb-weaving spider that uses its web as an auditory sensor. Proc. Natl. Acad. Sci. USA 2022, 119, e2122789119. [Google Scholar] [CrossRef]
- Cariani, P.; Baker, J.M. Time Is of the Essence: Neural Codes, Synchronies, Oscillations, Architectures. Front. Comput. Neurosci. 2022, 16, 898829. [Google Scholar] [CrossRef]
- Pietsch, P. Shufflebrain; Houghton Mifflin: Boston, MA, USA, 1981; 273p. [Google Scholar]
- Yasuda, H.; Buskohl, P.R.; Gillman, A.; Murphey, T.D.; Stepney, S.; Vaia, R.A.; Raney, J.R. Mechanical computing. Nature 2021, 598, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.C. The computational stance in biology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180380. [Google Scholar] [CrossRef] [Green Version]
- Parsa, A.; Wang, D.; O’Hern, C.S.; Shattuck, M.D.; Kramer-Bottiglio, R.; Bongard, J. Evolving Programmable Computational Metamaterials. In Proceedings of the Genetic and Evolutionary Computation Conference (GECCO ’22), Boston, MA, USA, 9–13 July 2022; pp. 122–129. [Google Scholar]
- Li, S.; Batra, R.; Brown, D.; Chang, H.D.; Ranganathan, N.; Hoberman, C.; Rus, D.; Lipson, H. Particle robotics based on statistical mechanics of loosely coupled components. Nature 2019, 567, 361–365. [Google Scholar] [CrossRef]
- Gondarenko, A.; Preble, S.; Robinson, J.; Chen, L.; Lipson, H.; Lipson, M. Spontaneous emergence of periodic patterns in a biologically inspired simulation of photonic structures. Phys. Rev. Lett. 2006, 96, 143904. [Google Scholar] [CrossRef]
- Thompson, A. An evolved circuit, intrinsic in silicon, entwined with physics. In Proceedings of the International Conference on Evolvable Systems, Tsukuba, Japan, 7–8 October 1996; pp. 390–405. [Google Scholar]
- Brodbeck, L.; Hauser, S.; Iida, F. Morphological Evolution of Physical Robots through Model-Free Phenotype Development. PLoS ONE 2015, 10, e0128444. [Google Scholar] [CrossRef] [Green Version]
- Abolafia, D.A.; Norouzi, M.; Shen, J.; Zhao, R.; Le, Q.V. Neural program synthesis with priority queue training. arXiv 2018, arXiv:1801.03526. [Google Scholar] [CrossRef]
- Sobania, D.; Briesch, M.; Rothlauf, F. Choose your programming copilot: A comparison of the program synthesis performance of github copilot and genetic programming. In Proceedings of the Genetic and Evolutionary Computation Conference, Boston, MA, USA, 9–13 July 2022; pp. 1019–1027. [Google Scholar]
- Karmaker, S.K.; Hassan, M.M.; Smith, M.J.; Xu, L.; Zhai, C.X.; Veeramachaneni, K. AutoML to Date and Beyond: Challenges and Opportunities. ACM Comput. Surv. 2021, 54, 175. [Google Scholar] [CrossRef]
- Bongard, J.; Levin, M. Living Things Are Not (20th Century) Machines: Updating Mechanism Metaphors in Light of the Modern Science of Machine Behavior. Front. Ecol. Evol. 2021, 9, 147. [Google Scholar] [CrossRef]
- Nicholson, D.J. Is the cell really a machine? J. Theor. Biol. 2019, 477, 108–126. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, D.J. The machine conception of the organism in development and evolution: A critical analysis. Stud. Hist. Philos. Biol. Biomed. Sci. 2014, 48 Pt B, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Witzany, G.; Baluska, F. Life’s code script does not code itself. The machine metaphor for living organisms is outdated. EMBO Rep. 2012, 13, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Box, G.E.P. Robustness in the Strategy of Scientific Model Building. In Robustness in Statistics; Launer, R.L., Wilkinson, G.N., Eds.; Academic Press: New York, NY, USA; London, UK, 1979; pp. 201–236. [Google Scholar]
- James, W. Pragmatism, a New Name for Some Old Ways of Thinking; Longmans, Green, and Co.: New York, NY, USA, 1907. [Google Scholar]
- Fields, C.; Levin, M. Competency in Navigating Arbitrary Spaces as an Invariant for Analyzing Cognition in Diverse Embodiments. Entropy 2022, 24, 819. [Google Scholar] [CrossRef]
- Prakash, C.; Fields, C.; Hoffman, D.D.; Prentner, R.; Singh, M. Fact, Fiction, and Fitness. Entropy 2020, 22, 514. [Google Scholar] [CrossRef] [PubMed]
- Fields, C.; Friston, K.; Glazebrook, J.F.; Levin, M. A free energy principle for generic quantum systems. Prog. Biophys. Mol. Biol. 2022, 173, 36–59. [Google Scholar] [CrossRef]
- Fields, C. Sciences of Observation. Philosophies 2018, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Dennett, D.C. The Intentional Stance; MIT Press: Cambridge, MA, USA, 1987; 388p. [Google Scholar]
- Pezzulo, G.; Levin, M. Top-down models in biology: Explanation and control of complex living systems above the molecular level. J. R. Soc. Interface 2016, 13, 20160555. [Google Scholar] [CrossRef] [Green Version]
- Pezzulo, G.; Levin, M. Re-membering the body: Applications of computational neuroscience to the top-down control of regeneration of limbs and other complex organs. Integr. Biol. 2015, 7, 1487–1517. [Google Scholar] [CrossRef] [Green Version]
- Nanos, V.; Levin, M. Multi-scale Chimerism: An experimental window on the algorithms of anatomical control. Cells Dev. 2022, 169, 203764. [Google Scholar] [CrossRef]
- Manicka, S.; Levin, M. Modeling somatic computation with non-neural bioelectric networks. Sci. Rep. 2019, 9, 18612. [Google Scholar] [CrossRef] [Green Version]
- Grozinger, L.; Amos, M.; Gorochowski, T.E.; Carbonell, P.; Oyarzún, D.A.; Stoof, R.; Fellermann, H.; Zuliani, P.; Tas, H.; Goñi-Moreno, A. Pathways to cellular supremacy in biocomputing. Nat. Commun. 2019, 10, 5250. [Google Scholar] [CrossRef] [Green Version]
- Solé, R.V.; Macia, J. Expanding the landscape of biological computation with synthetic multicellular consortia. Nat. Comput. 2013, 12, 485–497. [Google Scholar] [CrossRef]
- Macia, J.; Posas, F.; Solé, R.V. Distributed computation: The new wave of synthetic biology devices. Trends Biotechnol. 2012, 30, 342–349. [Google Scholar] [CrossRef]
- Ausländer, S.; Ausländer, D.; Müller, M.; Wieland, M.; Fussenegger, M. Programmable single-cell mammalian biocomputers. Nature 2012, 487, 123–127. [Google Scholar] [CrossRef]
- Regot, S.; Macia, J.; Conde, N.; Furukawa, K.; Kjellén, J.; Peeters, T.; Hohmann, S.; de Nadal, E.; Posas, F.; Solé, R. Distributed biological computation with multicellular engineered networks. Nature 2011, 469, 207–211. [Google Scholar] [CrossRef]
- Qian, L.; Winfree, E.; Bruck, J. Neural network computation with DNA strand displacement cascades. Nature 2011, 475, 368–372. [Google Scholar] [CrossRef]
- Nicholson, D.J. Organisms not equal Machines. Stud. Hist. Philos. Biol. Biomed. Sci. 2013, 44, 669–678. [Google Scholar] [CrossRef]
- Clawson, W.P.; Levin, M. Endless Forms Most Beautiful: Teleonomy and the bioengineering of chimeric and synthetic organisms. Biol. J. Linn. Soc. 2022, blac073. [Google Scholar] [CrossRef]
- Ollé-Vila, A.; Duran-Nebreda, S.; Conde-Pueyo, N.; Montañez, R.; Solé, R. A morphospace for synthetic organs and organoids: The possible and the actual. Integr. Biol. 2016, 8, 485–503. [Google Scholar] [CrossRef]
- Langton, C.G. Artificial Life: An Overview; MIT Press: Cambridge, MA, USA, 1995; pp. xi+340+346 of plates. [Google Scholar]
- Doursat, R.; Sánchez, C. Growing fine-grained multicellular robots. Soft Robot. 2014, 1, 110–121. [Google Scholar] [CrossRef]
- Doursat, R.; Sayama, H.; Michel, O. A review of morphogenetic engineering. Nat. Comput. 2013, 12, 517–535. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimkhani, M.R.; Levin, M. Synthetic living machines: A new window on life. iScience 2021, 24, 102505. [Google Scholar] [CrossRef]
- Ebrahimkhani, M.R.; Ebisuya, M. Synthetic developmental biology: Build and control multicellular systems. Curr. Opin. Chem. Biol. 2019, 52, 9–15. [Google Scholar] [CrossRef]
- Kamm, R.D.; Bashir, R.; Arora, N.; Dar, R.D.; Gillette, M.U.; Griffith, L.G.; Kemp, M.L.; Kinlaw, K.; Levin, M.; Martin, A.C.; et al. Perspective: The promise of multi-cellular engineered living systems. APL Bioeng. 2018, 2, 040901. [Google Scholar] [CrossRef] [Green Version]
- Kamm, R.D.; Bashir, R. Creating living cellular machines. Ann. Biomed. Eng. 2014, 42, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Levin, M. Life, death, and self: Fundamental questions of primitive cognition viewed through the lens of body plasticity and synthetic organisms. Biochem. Biophys. Res. Commun. 2021, 564, 114–133. [Google Scholar] [CrossRef]
- Levin, M. The Computational Boundary of a “Self”: Developmental Bioelectricity Drives Multicellularity and Scale-Free Cognition. Front. Psychol. 2019, 10, 2688. [Google Scholar] [CrossRef] [Green Version]
- Levin, M. Bioelectrical approaches to cancer as a problem of the scaling of the cellular self. Prog. Biophys. Mol. Biol. 2021, 165, 102–113. [Google Scholar] [CrossRef]
- Friston, K. Life as we know it. J. R. Soc. Interface 2013, 10, 20130475. [Google Scholar] [CrossRef] [Green Version]
- Ramstead, M.J.D.; Badcock, P.B.; Friston, K.J. Answering Schrodinger’s question: A free-energy formulation. Phys. Life Rev. 2018, 24, 1–16. [Google Scholar] [CrossRef]
- Constant, A.; Ramstead, M.J.D.; Veissière, S.P.L.; Campbell, J.O.; Friston, K.J. A variational approach to niche construction. J. R. Soc. Interface 2018, 15, 20170685. [Google Scholar] [CrossRef]
- Allen, M.; Friston, K.J. From cognitivism to autopoiesis: Towards a computational framework for the embodied mind. Synthese 2018, 195, 2459–2482. [Google Scholar] [CrossRef] [Green Version]
- Resnick, M. Turtles, Termites, and Traffic Jams: Explorations in Massively Parallel Microworlds, 1st ed.; MIT Press: Cambridge, MA, USA, 1997; pp. xviii+163p. [Google Scholar]
- Adamatzky, A. Towards fungal computer. Interface Focus 2018, 8, 20180029. [Google Scholar] [CrossRef]
- Siccardi, S.; Adamatzky, A. Models of Computing on Actin Filaments. In Advances in Unconventional Computing: Volume 2: Prototypes, Models and Algorithms; Adamatzky, A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 309–346. [Google Scholar]
- Maley, C.C. DNA computation: Theory, practice, and prospects. Evol. Comput. 1998, 6, 201–229. [Google Scholar] [CrossRef]
- Dorigo, M.; Di Caro, G. Ant colony optimization: A new meta-heuristic. In Proceedings of the 1999 Congress on Evolutionary Computation-CEC99 (Cat. No. 99TH8406), Washington, DC, USA, 6–9 July 1999; pp. 1470–1477. [Google Scholar]
- Churchland, P.S.; Sejnowski, T.J. The Computational Brain, 25th Anniversary ed.; The MIT Press: Cambridge, MA, USA, 2017; pp. xix+544p. [Google Scholar]
- Friston, K.J.; Stephan, K.E.; Montague, R.; Dolan, R.J. Computational psychiatry: The brain as a phantastic organ. Lancet Psychiatry 2014, 1, 148–158. [Google Scholar] [CrossRef]
- Corlett, P.R.; Fletcher, P.C. Computational psychiatry: A Rosetta Stone linking the brain to mental illness. Lancet Psychiatry 2014, 1, 399–402. [Google Scholar] [CrossRef]
- Bray, D. Wetware: A Computer in Every Living Cell; Yale University Press: New Haven, CT, USA, 2009; pp. xii+267p. [Google Scholar]
- Bray, D. Protein molecules as computational elements in living cells. Nature 1995, 376, 307–312. [Google Scholar] [CrossRef]
- Fields, C.; Bischof, J.; Levin, M. Morphological Coordination: A Common Ancestral Function Unifying Neural and Non-Neural Signaling. Physiology 2020, 35, 16–30. [Google Scholar] [CrossRef]
- Tanaka, G.; Yamane, T.; Heroux, J.B.; Nakane, R.; Kanazawa, N.; Takeda, S.; Numata, H.; Nakano, D.; Hirose, A. Recent advances in physical reservoir computing: A review. Neural Netw. 2019, 115, 100–123. [Google Scholar] [CrossRef]
- Jose, A.M. Replicating and Cycling Stores of Information Perpetuate Life. BioEssays 2018, 40, e1700161. [Google Scholar] [CrossRef]
- Davies, J.; Levin, M. Synthetic morphology via active and agential matter. Nat. Rev. Bioeng. 2022, 1, 46–59. [Google Scholar] [CrossRef]
- Manicka, S.; Levin, M. The Cognitive Lens: A primer on conceptual tools for analysing information processing in developmental and regenerative morphogenesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180369. [Google Scholar] [CrossRef] [Green Version]
- Nocera, D.G. The artificial leaf. Acc. Chem. Res. 2012, 45, 767–776. [Google Scholar] [CrossRef]
- Chin, Y.W.; Kok, J.M.; Zhu, Y.Q.; Chan, W.L.; Chahl, J.S.; Khoo, B.C.; Lau, G.K. Efficient flapping wing drone arrests high-speed flight using post-stall soaring. Sci. Robot. 2020, 5, eaba2386. [Google Scholar] [CrossRef]
- Leiserson, C.E.; Thompson, N.C.; Emer, J.S.; Kuszmaul, B.C.; Lampson, B.W.; Sanchez, D.; Schardl, T.B. There’s plenty of room at the Top: What will drive computer performance after Moore’s law? Science 2020, 368, eaam9744. [Google Scholar] [CrossRef]
- Rahwan, I.; Cebrian, M.; Obradovich, N.; Bongard, J.; Bonnefon, J.F.; Breazeal, C.; Crandall, J.W.; Christakis, N.A.; Couzin, I.D.; Jackson, M.O.; et al. Machine behaviour. Nature 2019, 568, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Boldt, J. Machine metaphors and ethics in synthetic biology. Life Sci. Soc. Policy 2018, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E.A.; Li, Y.I.; Pritchard, J.K. An Expanded View of Complex Traits: From Polygenic to Omnigenic. Cell 2017, 169, 1177–1186. [Google Scholar] [CrossRef] [Green Version]
- Plaut, D.C.; McClelland, J.L. Locating object knowledge in the brain: Comment on Bowers’s (2009) attempt to revive the grandmother cell hypothesis. Psychol. Rev. 2010, 117, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Bongard, J.C. Evolutionary robotics. Commun. ACM 2013, 56, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Wright, L.G.; Onodera, T.; Stein, M.M.; Wang, T.; Schachter, D.T.; Hu, Z.; McMahon, P.L. Deep physical neural networks trained with backpropagation. Nature 2022, 601, 549–555. [Google Scholar] [CrossRef]
- Kriegman, S.; Blackiston, D.; Levin, M.; Bongard, J. Kinematic self-replication in reconfigurable organisms. Proc. Natl. Acad. Sci. USA 2021, 118, e2112672118. [Google Scholar] [CrossRef]
- Kriegman, S.; Blackiston, D.; Levin, M.; Bongard, J. A scalable pipeline for designing reconfigurable organisms. Proc. Natl. Acad. Sci. USA 2020, 117, 1853–1859. [Google Scholar] [CrossRef] [Green Version]
- Floridi, L.; Chiriatti, M. GPT-3: Its Nature, Scope, Limits, and Consequences. Minds Mach. 2020, 30, 681–694. [Google Scholar] [CrossRef]
- Ramesh, A.; Dhariwal, P.; Nichol, A.; Chu, C.; Chen, M. Hierarchical text-conditional image generation with clip latents. arXiv 2022, arXiv:2204.06125. [Google Scholar] [CrossRef]
- Ortiz, C.L., Jr. Why we need a physically embodied Turing test and what it might look like. AI Mag. 2016, 37, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Giselbrecht, S.; Rapp, B.E.; Niemeyer, C.M. The chemistry of cyborgs--interfacing technical devices with organisms. Angew. Chem. Int. Ed. Engl. 2013, 52, 13942–13957. [Google Scholar] [CrossRef]
- Nam, Y.; Wheeler, B.C. In vitro microelectrode array technology and neural recordings. Crit. Rev. Biomed. Eng. 2011, 39, 45–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothschild, R.M. Neuroengineering tools/applications for bidirectional interfaces, brain-computer interfaces, and neuroprosthetic implants—A review of recent progress. Front. Neuroeng. 2010, 3, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuchling, F.; Friston, K.; Georgiev, G.; Levin, M. Morphogenesis as Bayesian inference: A variational approach to pattern formation and control in complex biological systems. Phys. Life Rev. 2020, 33, 88–108. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Levin, M.; Sengupta, B.; Pezzulo, G. Knowing one’s place: A free-energy approach to pattern regulation. J. R. Soc. Interface 2015, 12, 20141383. [Google Scholar] [CrossRef] [Green Version]
- Friston, K. A free energy principle for a particular physics. arXiv 2019, arXiv:1906.10184. [Google Scholar] [CrossRef]
- Genty, G.; Salmela, L.; Dudley, J.M.; Brunner, D.; Kokhanovskiy, A.; Kobtsev, S.; Turitsyn, S.K. Machine learning and applications in ultrafast photonics. Nat. Photonics 2021, 15, 91–101. [Google Scholar] [CrossRef]
- Nakajima, K. Physical reservoir computing—An introductory perspective. Jpn. J. Appl. Phys. 2020, 59, 060501. [Google Scholar] [CrossRef]
- Gyongyosi, L.; Imre, S. A Survey on quantum computing technology. Comput. Sci. Rev. 2019, 31, 51–71. [Google Scholar] [CrossRef]
- Paul, C. Morphological computation—A basis for the analysis of morphology and control requirements. Robot. Auton. Syst. 2006, 54, 619–630. [Google Scholar] [CrossRef]
- Zahedi, K.; Ay, N. Quantifying Morphological Computation. Entropy 2013, 15, 1887–1915. [Google Scholar] [CrossRef] [Green Version]
- Heimburg, T.; Jackson, A.D. On soliton propagation in biomembranes and nerves. Proc. Natl. Acad. Sci. USA 2005, 102, 9790–9795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoel, E.P. Agent Above, Atom Below: How Agents Causally Emerge from Their Underlying Microphysics. In Wandering towards a Goal: How Can Mindless Mathematical Laws Give Rise to Aims and Intention? Aguirre, A., Foster, B., Merali, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 63–76. [Google Scholar]
- Hoel, E.P. When the Map Is Better Than the Territory. Entropy 2017, 19, 188. [Google Scholar] [CrossRef] [Green Version]
- Albantakis, L.; Marshall, W.; Hoel, E.; Tononi, G. What Caused What? A Quantitative Account of Actual Causation Using Dynamical Causal Networks. Entropy 2019, 21, 459. [Google Scholar] [CrossRef] [Green Version]
- Hoel, E.P.; Albantakis, L.; Marshall, W.; Tononi, G. Can the macro beat the micro? Integrated information across spatiotemporal scales. Neurosci. Conscious. 2016, 2016, niw012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoel, E.P.; Albantakis, L.; Tononi, G. Quantifying causal emergence shows that macro can beat micro. Proc. Natl. Acad. Sci. USA 2013, 110, 19790–19795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, B.; Hoel, E. Uncertainty and causal emergence in complex networks. arXiv 2019, arXiv:1907.03902. [Google Scholar] [CrossRef]
- Kim, H.; Sayama, H. How Criticality of Gene Regulatory Networks Affects the Resulting Morphogenesis under Genetic Perturbations. Artif. Life 2018, 24, 85–105. [Google Scholar] [CrossRef] [Green Version]
- Graudenzi, A.; Serra, R.; Villani, M.; Colacci, A.; Kauffman, S.A. Robustness analysis of a Boolean model of gene regulatory network with memory. J. Comput. Biol. 2011, 18, 559–577. [Google Scholar] [CrossRef]
- Darabos, C.; Di Cunto, F.; Tomassini, M.; Moore, J.H.; Provero, P.; Giacobini, M. Additive functions in boolean models of gene regulatory network modules. PLoS ONE 2011, 6, e25110. [Google Scholar] [CrossRef] [Green Version]
- Bassel, G.W. Information Processing and Distributed Computation in Plant Organs. Trends Plant Sci. 2018, 23, 994–1005. [Google Scholar] [CrossRef]
- Calvo, P.; Friston, K. Predicting green: Really radical (plant) predictive processing. J. R. Soc. Interface 2017, 14, 20170096. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Kim, S.J.; Hara, M.; Aono, M. Remarkable problem-solving ability of unicellular amoeboid organism and its mechanism. R. Soc. Open Sci. 2018, 5, 180396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, R.; Amor, D.R.; Duran-Nebreda, S.; Conde-Pueyo, N.; Carbonell-Ballestero, M.; Montañez, R. Synthetic collective intelligence. Biosystems 2016, 148, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.R.; MacDonald, H.; Mann, R.P.; Marshall, J.A.; Latty, T.; Garnier, S. Decision-making without a brain: How an amoeboid organism solves the two-armed bandit. J. R. Soc. Interface 2016, 13, 20160030. [Google Scholar] [CrossRef] [Green Version]
- Vallverdú, J.; Castro, O.; Mayne, R.; Talanov, M.; Levin, M.; Baluška, F.; Gunji, Y.; Dussutour, A.; Zenil, H.; Adamatzky, A. Slime mould: The fundamental mechanisms of biological cognition. Biosystems 2018, 165, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugler, A.; Kittisopikul, M.; Hayden, L.; Liu, J.; Wiggins, C.H.; Süel, G.M.; Walczak, A.M. Noise Expands the Response Range of the Bacillus subtilis Competence Circuit. PLoS Comput. Biol. 2016, 12, e1004793. [Google Scholar] [CrossRef]
- Prindle, A.; Liu, J.; Asally, M.; Ly, S.; Garcia-Ojalvo, J.; Süel, G.M. Ion channels enable electrical communication in bacterial communities. Nature 2015, 527, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Larkin, J.W.; Zhai, X.; Kikuchi, K.; Redford, S.E.; Prindle, A.; Liu, J.; Greenfield, S.; Walczak, A.M.; Garcia-Ojalvo, J.; Mugler, A.; et al. Signal Percolation within a Bacterial Community. Cell Syst. 2018, 7, 137–145.e133. [Google Scholar] [CrossRef] [Green Version]
- Baluška, F.; Reber, A.S.; Miller, W.B., Jr. Cellular sentience as the primary source of biological order and evolution. Biosystems 2022, 218, 104694. [Google Scholar] [CrossRef]
- Baluška, F.; Miller, W.B.; Reber, A.S. Cellular and evolutionary perspectives on organismal cognition: From unicellular to multicellular organisms. Biol. J. Linn. Soc. 2022, blac005. [Google Scholar] [CrossRef]
- Reber, A.S.; Baluška, F. Cognition in some surprising places. Biochem. Biophys. Res. Commun. 2021, 564, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Baluška, F.; Levin, M. On Having No Head: Cognition throughout Biological Systems. Front. Psychol. 2016, 7, 902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M. Bioelectric signaling: Reprogrammable circuits underlying embryogenesis, regeneration, and cancer. Cell 2021, 184, 1971–1989. [Google Scholar] [CrossRef]
- Harris, M.P. Bioelectric signaling as a unique regulator of development and regeneration. Development 2021, 148, dev180794. [Google Scholar] [CrossRef] [PubMed]
- Bonzanni, M.; Rouleau, N.; Levin, M.; Kaplan, D.L. Optogenetically induced cellular habituation in non-neuronal cells. PLoS ONE 2020, 15, e0227230. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.S.; Uzel, S.G.; Akagi, J.; Wlodkowic, D.; Andreeva, V.; Yelick, P.C.; Devitt-Lee, A.; Paré, J.F.; Levin, M. Bioelectric signalling via potassium channels: A mechanism for craniofacial dysmorphogenesis in KCNJ2-associated Andersen-Tawil Syndrome. J. Physiol. 2016, 594, 3245–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.S.; Lemire, J.M.; Kramer, R.H.; Levin, M. Optogenetics in Developmental Biology: Using light to control ion flux-dependent signals in Xenopus embryos. Int. J. Dev. Biol. 2014, 58, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.S.; Tseng, A.S.; Levin, M. Light-activation of the Archaerhodopsin H(+)-pump reverses age-dependent loss of vertebrate regeneration: Sparking system-level controls in vivo. Biol. Open 2013, 2, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Fields, C.; Levin, M. Scale-Free Biology: Integrating Evolutionary and Developmental Thinking. BioEssays 2020, 42, e1900228. [Google Scholar] [CrossRef]
- Bizzari, M.; Brash, D.E.; Briscoe, J.; Grieneisen, V.A.; Stern, C.D.; Levin, M. A call for a better understanding of causation in cell biology. Nat. Rev. Mol. Cell Biol. 2019, 20, 261–262. [Google Scholar] [CrossRef]
- Stern, C.D. Reflections on the past, present and future of developmental biology. Dev. Biol. 2022, 488, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Jianying, H.; Mingjie, Z.; Tiangui, D.; Hui, L.; Yuhong, R. Optical holographic data storage—The time for new development. Opto-Electron. Eng. 2019, 46, 180642. [Google Scholar] [CrossRef]
- Kim, J.Z.; Bassett, D.S. A Neural Programming Language for the Reservoir Computer. arXiv 2022, arXiv:2203.05032. [Google Scholar] [CrossRef]
- Cardelli, L.; Csikász-Nagy, A. The cell cycle switch computes approximate majority. Sci. Rep. 2012, 2, 656. [Google Scholar] [CrossRef] [Green Version]
- Bray, D.; Gilbert, D. Cytoskeletal elements in neurons. Annu. Rev. Neurosci. 1981, 4, 505–523. [Google Scholar] [CrossRef] [PubMed]
- Aubin, C.A.; Gorissen, B.; Milana, E.; Buskohl, P.R.; Lazarus, N.; Slipher, G.A.; Keplinger, C.; Bongard, J.; Iida, F.; Lewis, J.A.; et al. Towards enduring autonomous robots via embodied energy. Nature 2022, 602, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Sanford, E.M.; Emert, B.L.; Cote, A.; Raj, A. Gene regulation gravitates toward either addition or multiplication when combining the effects of two signals. Elife 2020, 9, e59388. [Google Scholar] [CrossRef]
- Hameroff, S.R.; Craddock, T.J.; Tuszynski, J.A. “Memory bytes”—Molecular match for CaMKII phosphorylation encoding of microtubule lattices. J. Integr. Neurosci. 2010, 9, 253–267. [Google Scholar] [CrossRef] [Green Version]
- Craddock, T.J.; Beauchemin, C.; Tuszynski, J.A. Information processing mechanisms in microtubules at physiological temperature: Model predictions for experimental tests. Biosystems 2009, 97, 28–34. [Google Scholar] [CrossRef]
- Hentschel, H.G.; Fine, A.; Pencea, C.S. Biological computing with diffusion and excitable calcium stores. Math. Biosci. Eng. 2004, 1, 147–159. [Google Scholar] [CrossRef]
- Habibi, I.; Cheong, R.; Lipniacki, T.; Levchenko, A.; Emamian, E.S.; Abdi, A. Computation and measurement of cell decision making errors using single cell data. PLoS Comput. Biol. 2017, 13, e1005436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timsit, Y.; Gregoire, S.P. Towards the Idea of Molecular Brains. Int. J. Mol. Sci. 2021, 22, 11868. [Google Scholar] [CrossRef] [PubMed]
- Johnston, I.G.; Bassel, G.W. Identification of a bet-hedging network motif generating noise in hormone concentrations and germination propensity in Arabidopsis. J. R. Soc. Interface 2018, 15, 20180042. [Google Scholar] [CrossRef] [Green Version]
- Ray, S.K.; Valentini, G.; Shah, P.; Haque, A.; Reid, C.R.; Weber, G.F.; Garnier, S. Information Transfer During Food Choice in the Slime Mold Physarum polycephalum. Front. Ecol. Evol. 2019, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Iwayama, K.; Zhu, L.P.; Hirata, Y.; Aono, M.; Hara, M.; Aihara, K. Decision-making ability of Physarum polycephalum enhanced by its coordinated spatiotemporal oscillatory dynamics. Bioinspir. Biomim. 2016, 11, 036001. [Google Scholar] [CrossRef] [PubMed]
- Tarabella, G.; D’Angelo, P.; Cifarelli, A.; Dimonte, A.; Romeo, A.; Berzina, T.; Erokhin, V.; Iannotta, S. A hybrid living/organic electrochemical transistor based on the Physarum polycephalum cell endowed with both sensing and memristive properties. Chem. Sci. 2015, 6, 2859–2868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamatzky, A. Slime mould processors, logic gates and sensors. Philos. Trans. A Math. Phys. Eng. Sci. 2015, 373, 20140216. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.; Adamatzky, A. Towards Physarum binary adders. BioSystems 2010, 101, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Katz, Y.; Goodman, N.D.; Kersting, K.; Kemp, C.; Tenenbaum, J.B. Modeling Semantic Cognition as Logical Dimensionality Reduction. In Proceedings of the 30th Annual Conference of the Cognitive Science Society (CogSci 2008), Washington, DC, USA, 23–26 July 2008. [Google Scholar]
- Katz, Y. Embodying probabilistic inference in biochemical circuits. arXiv 2018, arXiv:1806.10161. [Google Scholar] [CrossRef]
- Katz, Y.; Springer, M. Probabilistic adaptation in changing microbial environments. PeerJ 2016, 4, e2716. [Google Scholar] [CrossRef] [Green Version]
- Tay, S.; Hughey, J.J.; Lee, T.K.; Lipniacki, T.; Quake, S.R.; Covert, M.W. Single-cell NF-kappaB dynamics reveal digital activation and analogue information processing. Nature 2010, 466, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzulo, G.; LaPalme, J.; Durant, F.; Levin, M. Bistability of somatic pattern memories: Stochastic outcomes in bioelectric circuits underlying regeneration. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20190765. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Pietak, A.M.; Bischof, J. Planarian regeneration as a model of anatomical homeostasis: Recent progress in biophysical and computational approaches. Semin. Cell Dev. Biol. 2018, 87, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Pietak, A.; Levin, M. Bioelectric gene and reaction networks: Computational modelling of genetic, biochemical and bioelectrical dynamics in pattern regulation. J. R. Soc. Interface 2017, 14, 20170425. [Google Scholar] [CrossRef] [PubMed]
- Fields, C.; Levin, M. Multiscale memory and bioelectric error correction in the cytoplasm—Cytoskeleton-membrane system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 10, e1410. [Google Scholar] [CrossRef]
- Law, R.; Levin, M. Bioelectric memory: Modeling resting potential bistability in amphibian embryos and mammalian cells. Theor. Biol. Med. Model. 2015, 12, 22. [Google Scholar] [CrossRef] [Green Version]
- Raman, K.; Wagner, A. The evolvability of programmable hardware. J. R. Soc. Interface 2011, 8, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Manicka, S.; Levin, M. Minimal Developmental Computation: A Causal Network Approach to Understand Morphogenetic Pattern Formation. Entropy 2022, 24, 107. [Google Scholar] [CrossRef]
- Durant, F.; Bischof, J.; Fields, C.; Morokuma, J.; LaPalme, J.; Hoi, A.; Levin, M. The Role of Early Bioelectric Signals in the Regeneration of Planarian Anterior/Posterior Polarity. Biophys. J. 2019, 116, 948–961. [Google Scholar] [CrossRef] [Green Version]
- Stepney, S.; Kendon, V. The representational entity in physical computing. Nat. Comput. 2021, 20, 233–242. [Google Scholar] [CrossRef]
- Berend, D.; Dolev, S.; Frenkel, S.; Hanemann, A. Towards holographic “brain” memory based on randomization and Walsh-Hadamard transformation. Neural Netw. 2016, 77, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Wess, O.; Roder, U. A holographic model for associative memory chains. Biol. Cybern. 1977, 27, 89–98. [Google Scholar] [CrossRef]
- Levin, M. Collective Intelligence of Morphogenesis as a Teleonomic Process. In Evolution “on Purpose”: Teleonomy in Living Systems; Corning, P.A., Kauffman, S.A., Noble, D., Shapiro, J.A., Vane-Wright, R.I., Pross, A., Eds.; MIT Press: Cambridge, MA, USA, 2023; pp. 175–198. [Google Scholar]
- Sajid, N.; Parr, T.; Hope, T.M.; Price, C.J.; Friston, K.J. Degeneracy and Redundancy in Active Inference. Cereb. Cortex 2020, 30, 5750–5766. [Google Scholar] [CrossRef] [PubMed]
- Madhani, H.D. The frustrated gene: Origins of eukaryotic gene expression. Cell 2013, 155, 744–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soen, Y.; Knafo, M.; Elgart, M. A principle of organization which facilitates broad Lamarckian-like adaptations by improvisation. Biol. Direct 2015, 10, 68. [Google Scholar] [CrossRef] [Green Version]
- Elgart, M.; Snir, O.; Soen, Y. Stress-mediated tuning of developmental robustness and plasticity in flies. Biochim. Biophys. Acta 2015, 1849, 462–466. [Google Scholar] [CrossRef]
- McEwen, B.S. Stress, adaptation, and disease. Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef]
- Ball, J.M.; Chen, S.; Li, W. Mitochondria in cone photoreceptors act as microlenses to enhance photon delivery and confer directional sensitivity to light. Sci. Adv. 2022, 8, eabn2070. [Google Scholar] [CrossRef]
- Wrabl, J.O.; Gu, J.; Liu, T.; Schrank, T.P.; Whitten, S.T.; Hilser, V.J. The role of protein conformational fluctuations in allostery, function, and evolution. Biophys. Chem. 2011, 159, 129–141. [Google Scholar] [CrossRef]
- Freddolino, P.L.; Yang, J.; Momen-Roknabadi, A.; Tavazoie, S. Stochastic tuning of gene expression enables cellular adaptation in the absence of pre-existing regulatory circuitry. Elife 2018, 7, e31867. [Google Scholar] [CrossRef]
- Freddolino, P.L.; Tavazoie, S. Beyond homeostasis: A predictive-dynamic framework for understanding cellular behavior. Annu. Rev. Cell Dev. Biol. 2012, 28, 363–384. [Google Scholar] [CrossRef] [PubMed]
- Schreier, H.I.; Soen, Y.; Brenner, N. Exploratory adaptation in large random networks. Nat. Commun. 2017, 8, 14826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Clawson, W.; Levin, M. Learning in Transcriptional Network Models: Computational Discovery of Pathway-Level Memory and Effective Interventions. Int. J. Mol. Sci. 2023, 24, 285. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Manicka, S.; Hoel, E.; Levin, M. Gene Regulatory Networks Exhibit Several Kinds of Memory: Quantification of Memory in Biological and Random Transcriptional Networks. iScience 2021, 24, 102131. [Google Scholar] [CrossRef] [PubMed]
- McGregor, S.; Vasas, V.; Husbands, P.; Fernando, C. Evolution of associative learning in chemical networks. PLoS Comput. Biol. 2012, 8, e1002739. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.A.; Buckley, C.L.; Mills, R.; Davies, A. Associative memory in gene regulation networks. In Proceedings of the Artificial Life Conference XII, Odense, Denmark, 19–23 August 2010; pp. 194–201. [Google Scholar]
- Szabó, Á.; Vattay, G.; Kondor, D. A cell signaling model as a trainable neural nanonetwork. Nano Commun. Netw. 2012, 3, 57–64. [Google Scholar] [CrossRef]
- Herrera-Delgado, E.; Perez-Carrasco, R.; Briscoe, J.; Sollich, P. Memory functions reveal structural properties of gene regulatory networks. PLoS Comput. Biol. 2018, 14, e1006003. [Google Scholar] [CrossRef]
- Chen, Z.; Linton, J.M.; Zhu, R.; Elowitz, M.B. A synthetic protein-level neural network in mammalian cells. bioRxiv 2022. [Google Scholar] [CrossRef]
- Lau, M.W.; Ferré-D’Amaré, A.R. Many Activities, One Structure: Functional Plasticity of Ribozyme Folds. Molecules 2016, 21, 1570. [Google Scholar] [CrossRef] [Green Version]
- Müller, F.; Escobar, L.; Xu, F.; Wegrzyn, E.; Nainyte, M.; Amatov, T.; Chan, C.Y.; Pichler, A.; Carell, T. A prebiotically plausible scenario of an RNA-peptide world. Nature 2022, 605, 279–284. [Google Scholar] [CrossRef]
- Couzigou, J.M.; Lauressergues, D.; Bécard, G.; Combier, J.P. miRNA-encoded peptides (miPEPs): A new tool to analyze the roles of miRNAs in plant biology. RNA Biol. 2015, 12, 1178–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raina, M.; King, A.; Bianco, C.; Vanderpool, C.K. Dual-Function RNAs. Microbiol. Spectr. 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Historical review: ATP as a neurotransmitter. Trends Pharmacol. Sci. 2006, 27, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Makalowska, I.; Lin, C.F.; Makalowski, W. Overlapping genes in vertebrate genomes. Comput. Biol. Chem. 2005, 29, 1–12. [Google Scholar] [CrossRef]
- Wright, B.W.; Molloy, M.P.; Jaschke, P.R. Overlapping genes in natural and engineered genomes. Nat. Rev. Genet. 2022, 23, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Tosteson, M.T.; Kim, J.B.; Goldstein, D.J.; Tosteson, D.C. Ion channels formed by transcription factors recognize consensus DNA sequences. Biochim. Biophys. Acta 2001, 1510, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Meléndez García, R.; Haccard, O.; Chesneau, A.; Narassimprakash, H.; Roger, J.; Perron, M.; Marheineke, K.; Bronchain, O. A non-transcriptional function of Yap regulates the DNA replication program in Xenopus laevis. Elife 2022, 11, e75741. [Google Scholar] [CrossRef]
- Elhage, N.; Hume, T.; Olsson, C.; Schiefer, N.; Henighan, T.; Kravec, S.; Hatfield-Dodds, Z.; Lasenby, R.; Drain, D.; Chen, C.; et al. Toy Models of Superposition. arXiv 2022, arXiv:2209.10652. [Google Scholar] [CrossRef]
- Solé, R.; Seoane, L.F. Polysemy and power: Ambiguity in language networks. Linguist. Rev. 2015, 32, 5–35. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, W.; Iizuka, H.; Yamamoto, M.; Taguchi, S. Superposition mechanism as a neural basis for understanding others. Sci. Rep. 2022, 12, 2859. [Google Scholar] [CrossRef]
- Lahoz-Beltra, R.; Hameroff, S.R.; Dayhoff, J.E. Cytoskeletal logic: A model for molecular computation via Boolean operations in microtubules and microtubule-associated proteins. Biosystems 1993, 29, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Craddock, T.J.; Tuszynski, J.A.; Hameroff, S. Cytoskeletal signaling: Is memory encoded in microtubule lattices by CaMKII phosphorylation? PLoS Comput. Biol. 2012, 8, e1002421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, B.T.; Garbus, J.; Pollack, J.B.; Marshall, W.F. A unicellular walker controlled by a microtubule-based finite-state machine. Curr. Biol. 2022, 32, 3745–3757.e3747. [Google Scholar] [CrossRef]
- Tuszynski, J.A.; Friesen, D.; Freedman, H.; Sbitnev, V.I.; Kim, H.; Santelices, I.; Kalra, A.P.; Patel, S.D.; Shankar, K.; Chua, L.O. Microtubules as Sub-Cellular Memristors. Sci. Rep. 2020, 10, 2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priel, A.; Tuszynski, J.A.; Woolf, N.J. Neural cytoskeleton capabilities for learning and memory. J. Biol. Phys. 2010, 36, 3–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantero, M.D.R.; Villa Etchegoyen, C.; Perez, P.L.; Scarinci, N.; Cantiello, H.F. Bundles of Brain Microtubules Generate Electrical Oscillations. Sci. Rep. 2018, 8, 11899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priel, A.; Ramos, A.J.; Tuszynski, J.A.; Cantiello, H.F. A biopolymer transistor: Electrical amplification by microtubules. Biophys. J. 2006, 90, 4639–4643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuszynski, J.A.; Portet, S.; Dixon, J.M.; Luxford, C.; Cantiello, H.F. Ionic wave propagation along actin filaments. Biophys. J. 2004, 86, 1890–1903. [Google Scholar] [CrossRef] [Green Version]
- Hameroff, S.; Nip, A.; Porter, M.; Tuszynski, J. Conduction pathways in microtubules, biological quantum computation, and consciousness. Biosystems 2002, 64, 149–168. [Google Scholar] [CrossRef]
- Jibu, M.; Hagan, S.; Hameroff, S.R.; Pribram, K.H.; Yasue, K. Quantum optical coherence in cytoskeletal microtubules: Implications for brain function. Biosystems 1994, 32, 195–209. [Google Scholar] [CrossRef]
- Rosen, M.R.; Binah, O.; Marom, S. Cardiac memory and cortical memory: Do learning patterns in neural networks impact on cardiac arrhythmias? Circulation 2003, 108, 1784–1789. [Google Scholar] [CrossRef] [Green Version]
- Zoghi, M. Cardiac memory: Do the heart and the brain remember the same? J. Interv. Card. Electrophysiol. 2005, 11, 177–182. [Google Scholar] [CrossRef]
- Chakravarthy, S.V.; Ghosh, J. On Hebbian-like adaptation in heart muscle: A proposal for ‘cardiac memory’. Biol. Cybern. 1997, 76, 207–215. [Google Scholar] [CrossRef]
- Fusi, S.; Miller, E.K.; Rigotti, M. Why neurons mix: High dimensionality for higher cognition. Curr. Opin. Neurobiol. 2016, 37, 66–74. [Google Scholar] [CrossRef]
- Quach, K.T.; Chalasani, S.H. Flexible reprogramming of Pristionchus pacificus motivation for attacking Caenorhabditis elegans in predator-prey competition. Curr. Biol. 2022, 32, 1675–1688.e1677. [Google Scholar] [CrossRef]
- Baars, B.J.; Franklin, S. An architectural model of conscious and unconscious brain functions: Global Workspace Theory and IDA. Neural Netw. 2007, 20, 955–961. [Google Scholar] [CrossRef]
- Dorahy, M.J.; Brand, B.L.; Şar, V.; Krüger, C.; Stavropoulos, P.; Martínez-Taboas, A.; Lewis-Fernández, R.; Middleton, W. Dissociative identity disorder: An empirical overview. Aust. N. Z. J. Psychiatry 2014, 48, 402–417. [Google Scholar] [CrossRef] [Green Version]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]
- De Pitta, M.; Volman, V.; Levine, H.; Ben-Jacob, E. Multimodal encoding in a simplified model of intracellular calcium signaling. Cogn. Process. 2009, 10 (Suppl. 1), S55–S70. [Google Scholar] [CrossRef]
- Chakraborty, A.; Alam, M.; Dey, V.; Chattopadhyay, A.; Mukhopadhyay, D. A survey on adversarial attacks and defences. CAAI Trans. Intell. Technol. 2021, 6, 25–45. [Google Scholar] [CrossRef]
- Martinez-Corral, R.; Liu, J.; Prindle, A.; Süel, G.M.; Garcia-Ojalvo, J. Metabolic basis of brain-like electrical signalling in bacterial communities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.D.; Galera-Laporta, L.; Bialecka-Fornal, M.; Moon, E.C.; Shen, Z.; Briggs, S.P.; Garcia-Ojalvo, J.; Süel, G.M. Magnesium Flux Modulates Ribosomes to Increase Bacterial Survival. Cell 2019, 177, 352–360.e313. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Corral, R.; Liu, J.; Süel, G.M.; Garcia-Ojalvo, J. Bistable emergence of oscillations in growing Bacillus subtilis biofilms. Proc. Natl. Acad. Sci. USA 2018, 115, E8333–E8340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Martinez-Corral, R.; Prindle, A.; Lee, D.D.; Larkin, J.; Gabalda-Sagarra, M.; Garcia-Ojalvo, J.; Süel, G.M. Coupling between distant biofilms and emergence of nutrient time-sharing. Science 2017, 356, 638–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.D.; Prindle, A.; Liu, J.; Süel, G.M. SnapShot: Electrochemical Communication in Biofilms. Cell 2017, 170, 214–214.e211. [Google Scholar] [CrossRef] [PubMed]
- Jablonka, E.; Lamb, M.J.; Zeligowski, A. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life, revised edition; A Bradford Book; The MIT Press: Cambridge, MA, USA; London, UK, 2014; pp. xii+563p. [Google Scholar]
- Jablonka, E.; Lamb, M.J.; Avital, E. ‘Lamarckian’ mechanisms in darwinian evolution. Trends Ecol. Evol. 1998, 13, 206–210. [Google Scholar] [CrossRef]
- Blackiston, D.; Lederer, E.K.; Kriegman, S.; Garnier, S.; Bongard, J.; Levin, M. A cellular platform for the development of synthetic living machines. Sci. Robot. 2021, 6, eabf1571. [Google Scholar] [CrossRef]
- Kouvaris, K.; Clune, J.; Kounios, L.; Brede, M.; Watson, R.A. How evolution learns to generalise: Using the principles of learning theory to understand the evolution of developmental organisation. PLoS Comput. Biol. 2017, 13, e1005358. [Google Scholar] [CrossRef] [Green Version]
- Žliobaite, I.; Stenseth, N.C. Improving Adaptation through Evolution and Learning: A Response to Watson and Szathmary. Trends Ecol. Evol. 2016, 31, 892–893. [Google Scholar] [CrossRef]
- Watson, R.A.; Szathmáry, E. How Can Evolution Learn? Trends Ecol. Evol. 2016, 31, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Watson, R.A.; Mills, R.; Buckley, C.L.; Kouvaris, K.; Jackson, A.; Powers, S.T.; Cox, C.; Tudge, S.; Davies, A.; Kounios, L.; et al. Evolutionary Connectionism: Algorithmic Principles Underlying the Evolution of Biological Organisation in Evo-Devo, Evo-Eco and Evolutionary Transitions. Evol. Biol. 2016, 43, 553–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livnat, A.; Papadimitriou, C. Evolution and Learning: Used Together, Fused Together. A Response to Watson and Szathmáry. Trends Ecol. Evol. 2016, 31, 894–896. [Google Scholar] [CrossRef]
- Blackiston, D.; Kriegman, S.; Bongard, J.; Levin, M. Biological Robots: Perspectives on an Emerging Interdisciplinary Field. arXiv 2022, arXiv:2207.00880. [Google Scholar] [CrossRef]
- Sullivan, K.G.; Emmons-Bell, M.; Levin, M. Physiological inputs regulate species-specific anatomy during embryogenesis and regeneration. Commun. Integr. Biol. 2016, 9, e1192733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmons-Bell, M.; Durant, F.; Hammelman, J.; Bessonov, N.; Volpert, V.; Morokuma, J.; Pinet, K.; Adams, D.S.; Pietak, A.; Lobo, D.; et al. Gap Junctional Blockade Stochastically Induces Different Species-Specific Head Anatomies in Genetically Wild-Type Girardia dorotocephala Flatworms. Int. J. Mol. Sci. 2015, 16, 27865–27896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.; Dennett, D.C. Cognition all the way down. Aeon Essays 2020. Available online: chrome-extension://oemmndcbldboiebfnladdacbdfmadadm/https://ase.tufts.edu/cogstud/dennett/papers/Levin_Dennett_Aeon.pdf (accessed on 23 February 2023).
- Jakobi, N.; Husbands, P.; Harvey, I. Noise and the reality gap: The use of simulation in evolutionary robotics. In Advances in Artificial Life. ECAL 1995; Lecture Notes in Computer Science; Morán, F., Moreno, A., Merelo, J.J., Chacón, P., Eds.; Springer: Berlin/Heidelberg, Germany, 1995; pp. 704–720. [Google Scholar]
- Tobin, J.; Fong, R.; Ray, A.; Schneider, J.; Zaremba, W.; Abbeel, P. Domain randomization for transferring deep neural networks from simulation to the real world. In Proceedings of the 2017 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Vancouver, BC, Canada, 24–28 September 2017; pp. 23–30. [Google Scholar]
- Bickhard, M.H.; Campbell, D.T. Emergence. In Downward Causation: Minds, Bodies and Matter; Andersen, P.B., Christiansen, P.V., Emmeche, C., Finnemann, N.O., Eds.; Aarhus University Press: Aarhus, Denmark, 2000; pp. 322–348. [Google Scholar]
- Whitehead, A. Process and Reality: An Essay in Cosmology; Free Press: New York, NY, USA, 1978. [Google Scholar]
- Hofstadter, D.R. Godel, Escher, Bach: An Eternal Golden Braid; Basic Books: New York, NY, USA, 1979; pp. xxi+777p. [Google Scholar]
- Blackiston, D.J.; Levin, M. Ectopic eyes outside the head in Xenopus tadpoles provide sensory data for light-mediated learning. J. Exp. Biol. 2013, 216, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Morrie, R.D.; Adams, D.S. V-ATPase-dependent ectodermal voltage and pH regionalization are required for craniofacial morphogenesis. Dev. Dyn. 2011, 240, 1889–1904. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Adams, D.S.; Levin, M. Normalized shape and location of perturbed craniofacial structures in the Xenopus tadpole reveal an innate ability to achieve correct morphology. Dev. Dyn. 2012, 241, 863–878. [Google Scholar] [CrossRef] [Green Version]
- Blackiston, D.J.; Vien, K.; Levin, M. Serotonergic stimulation induces nerve growth and promotes visual learning via posterior eye grafts in a vertebrate model of induced sensory plasticity. npj Regen. Med. 2017, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Blackiston, D.J.; Anderson, G.M.; Rahman, N.; Bieck, C.; Levin, M. A novel method for inducing nerve growth via modulation of host resting potential: Gap junction-mediated and serotonergic signaling mechanisms. Neurotherapeutics 2015, 12, 170–184. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, S.A. The Origins of Order: Self Organization and Selection in Evolution; Oxford University Press: New York, NY, USA, 1993; pp. xviii+709. [Google Scholar]
- Alvarez-Buylla, E.R.; Balleza, E.; Benítez, M.; Espinosa-Soto, C.; Padilla-Longoria, P. Gene regulatory network models: A dynamic and integrative approach to development. SEB Exp. Biol. Ser. 2008, 61, 113–139. [Google Scholar] [PubMed]
- Sauka-Spengler, T.; Bronner-Fraser, M. A gene regulatory network orchestrates neural crest formation. Nat. Rev. Mol. Cell Biol. 2008, 9, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Peter, I.S.; Davidson, E.H. Evolution of gene regulatory networks controlling body plan development. Cell 2011, 144, 970–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramson, C.I.; Levin, M. Behaviorist approaches to investigating memory and learning: A primer for synthetic biology and bioengineering. Commun. Integr. Biol. 2021, 14, 230–247. [Google Scholar] [CrossRef]
- Süel, G.M.; Garcia-Ojalvo, J.; Liberman, L.M.; Elowitz, M.B. An excitable gene regulatory circuit induces transient cellular differentiation. Nature 2006, 440, 545–550. [Google Scholar] [CrossRef]
- Ho, C.; Morsut, L. Novel synthetic biology approaches for developmental systems. Stem Cell Rep. 2021, 16, 1051–1064. [Google Scholar] [CrossRef]
- Toda, S.; Blauch, L.R.; Tang, S.K.Y.; Morsut, L.; Lim, W.A. Programming self-organizing multicellular structures with synthetic cell-cell signaling. Science 2018, 361, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Chater, N. The Mind Is Flat: The Illusion of Mental Depth and the Improvised Mind; Yale University Press: New Haven, CT, USA; London, UK, 2018. [Google Scholar]
- Nasser, A.; Al Haj Hassan, H.; Abou Chaaya, J.; Mansour, A.; Yao, K.C. Spectrum Sensing for Cognitive Radio: Recent Advances and Future Challenge. Sensors 2021, 21, 2408. [Google Scholar] [CrossRef]
- Ronellenfitsch, H.; Lasser, J.; Daly, D.C.; Katifori, E. Topological Phenotypes Constitute a New Dimension in the Phenotypic Space of Leaf Venation Networks. PLoS Comput. Biol. 2015, 11, e1004680. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.P.; Stadler, P.F. Quasi-independence, homology and the unity of type: A topological theory of characters. J. Theor. Biol. 2003, 220, 505–527. [Google Scholar] [CrossRef] [Green Version]
- Stadler, B.M.; Stadler, P.F.; Wagner, G.P.; Fontana, W. The topology of the possible: Formal spaces underlying patterns of evolutionary change. J. Theor. Biol. 2001, 213, 241–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mynard, F.; Seal, G.J. Phenotype spaces. J. Math. Biol. 2010, 60, 247–266. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.A. The extended phenotype(s): A comparison with niche construction theory. Biol. Philos. 2015, 30, 547–567. [Google Scholar] [CrossRef]
- Scholl, B.J.; Tremoulet, P.D. Perceptual causality and animacy. Trends Cogn. Sci. 2000, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Bloom, P.; Veres, C. The perceived intentionality of groups. Cognition 1999, 71, B1–B9. [Google Scholar] [CrossRef] [PubMed]
- Gunji, Y.P.; Nishiyama, Y.; Adamatzky, A. Robust soldier crab ball gate. AIP Conf. Proc. 2011, 1389, 995. [Google Scholar] [CrossRef] [Green Version]
- Adamatzky, A.; Erokhin, V.; Grube, M.; Schubert, T.; Schumann, A. Physarum Chip Project: Growing Computers from Slime Mould. Int. J. Unconv. Comput. 2012, 8, 319–323. [Google Scholar]
- Adamatzky, A. A brief history of liquid computers. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180372. [Google Scholar] [CrossRef] [Green Version]
- Rendell, P. Turing Universality of the Game of Life. In Collision-Based Computing; Adamatzky, A., Ed.; Springer: London, UK, 2002; pp. 513–539. [Google Scholar]
- Boudry, M.; Pigliucci, M. The mismeasure of machine: Synthetic biology and the trouble with engineering metaphors. Stud. Hist. Philos. Biol. Biomed. Sci. 2013, 44, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Bayne, T. On the axiomatic foundations of the integrated information theory of consciousness. Neurosci. Conscious. 2018, 2018, niy007. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, C.; Ravoo, B.J.; van der Wiel, W.G.; Wegner, S.V.; Pernice, W.H.P. The rise of intelligent matter. Nature 2021, 594, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Koseska, A.; Bastiaens, P.I. Cell signaling as a cognitive process. EMBO J. 2017, 36, 568–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNerney, M.P.; Doiron, K.E.; Ng, T.L.; Chang, T.Z.; Silver, P.A. Theranostic cells: Emerging clinical applications of synthetic biology. Nat. Rev. Genet. 2021, 22, 730–746. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bongard, J.; Levin, M. There’s Plenty of Room Right Here: Biological Systems as Evolved, Overloaded, Multi-Scale Machines. Biomimetics 2023, 8, 110. https://doi.org/10.3390/biomimetics8010110
Bongard J, Levin M. There’s Plenty of Room Right Here: Biological Systems as Evolved, Overloaded, Multi-Scale Machines. Biomimetics. 2023; 8(1):110. https://doi.org/10.3390/biomimetics8010110
Chicago/Turabian StyleBongard, Joshua, and Michael Levin. 2023. "There’s Plenty of Room Right Here: Biological Systems as Evolved, Overloaded, Multi-Scale Machines" Biomimetics 8, no. 1: 110. https://doi.org/10.3390/biomimetics8010110