
Targeting IgE and Th2-Cytokines in Allergy: Brief Updates on Monoclonal Antibodies and Antibody Gene Therapy

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Search Strategy
3. Monoclonal Antibody Therapy for Allergy
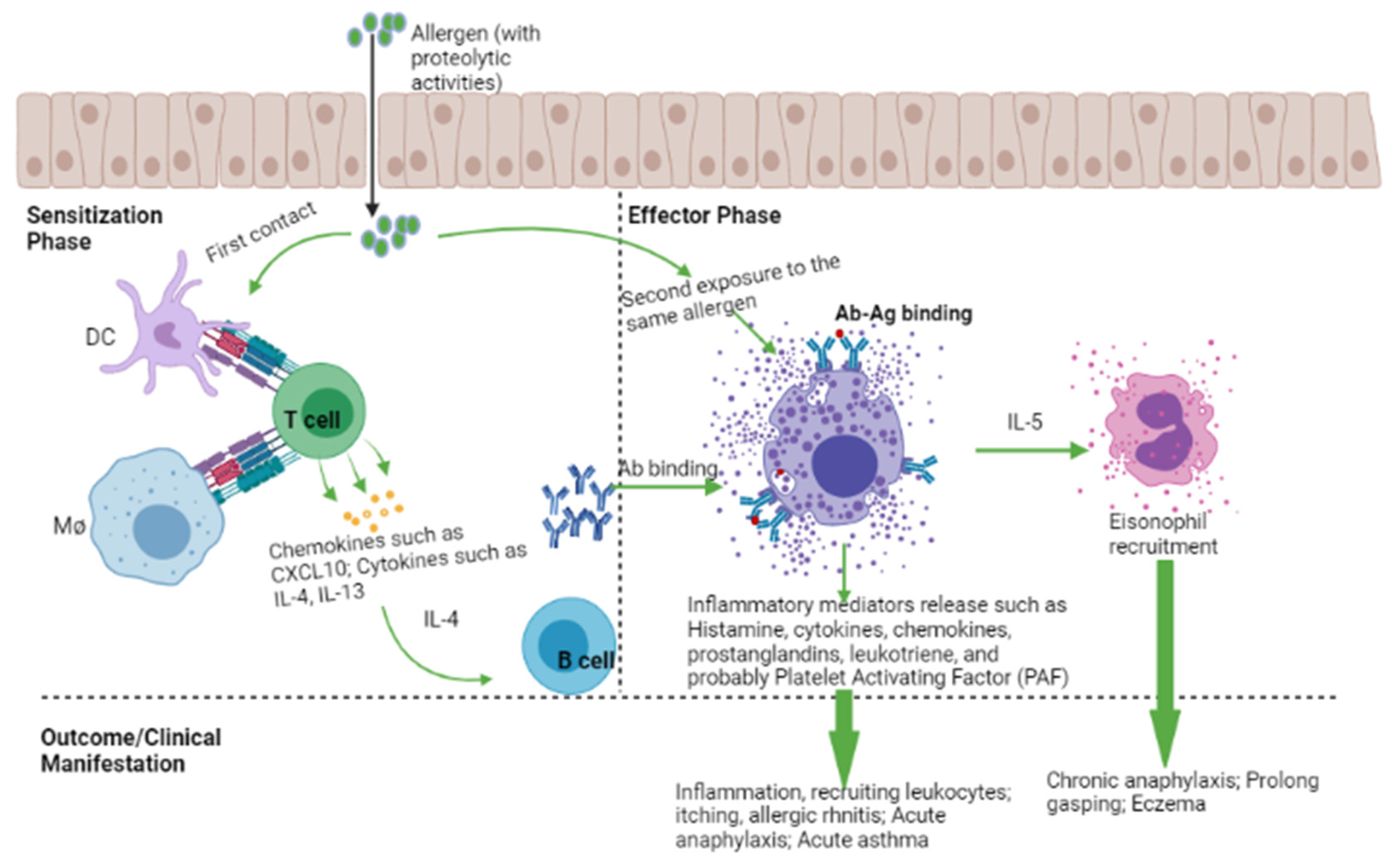
3.1. Mechanism of an Allergic Reaction
3.2. Histamine, Histamine Receptors, and Clinical Outcome
4. Possible Therapeutic Targets That Alleviate Allergic Diseases
4.1. Targeting IgE in Allergic Inflammation
4.2. Targeting Th2-Associated Cytokines in Allergic Disease
4.3. Monoclonal Antibodies Targeting Dual Inflammatory Mediators
4.4. Setbacks and Limitations
5. Antibody Gene Therapy: The Future for Antibody Therapy
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kubo, M. Innate and Adaptive Type 2 Immunity in Lung Allergic Inflammation. Immunol. Rev. 2017, 278, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The Development of Allergic Inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, T.; Tsutsui, H.; Tominaga, K.; Hoshino, K.; Okamura, H.; Akira, S.; Paul, W.E.; Nakanishi, K. IL-18, Although Antiallergic When Administered with IL-12, Stimulates IL-4 and Histamine Release by Basophils. Proc. Natl. Acad. Sci. USA 1999, 96, 13962–13966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J. Pathophysiology of Allergic Inflammation. Immunol. Rev. 2011, 242, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Junttila, I.S. Tuning the Cytokine Responses: An Update on Interleukin (IL)-4 and IL-13 Receptor Complexes. Front. Immunol. 2018, 9, 888. [Google Scholar] [CrossRef] [Green Version]
- Pawankar, R. Allergic Diseases and Asthma: A Global Public Health Concern and a Call to Action. World Allergy Organ. J. 2014, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Pawankar, R.; Canonica, G.W.; Holgate, S.T.; Lockey, R.F. Allergic Diseases and Asthma: A Major Global Health Concern. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 39–41. [Google Scholar] [CrossRef]
- Cayrol, C.; Duval, A.; Schmitt, P.; Roga, S.; Camus, M.; Stella, A.; Burlet-Schiltz, O.; Gonzalez-de-Peredo, A.; Girard, J.-P. Environmental Allergens Induce Allergic Inflammation through Proteolytic Maturation of IL-33. Nat. Immunol. 2018, 19, 375–385. [Google Scholar] [CrossRef]
- Mackay, I.R.; Rosen, F.S.; Kay, A. Allergy and Allergic Diseases. N. Engl. J. Med. 2001, 344, 30–37. [Google Scholar]
- Tang, Y.-J.; Chang, H.-H.; Chiang, C.-Y.; Lai, C.-Y.; Hsu, M.-Y.; Wang, K.-R.; Han, H.-H.; Chen, L.-Y.; Lin, D.P.-C. A Murine Model of Acute Allergic Conjunctivitis Induced by Continuous Exposure to Particulate Matter 2.5. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2118–2126. [Google Scholar] [CrossRef] [Green Version]
- Woo, L.N.; Guo, W.Y.; Wang, X.; Young, A.; Salehi, S.; Hin, A.; Zhang, Y.; Scott, J.A.; Chow, C.W. A 4-Week Model of House Dust Mite (HDM) Induced Allergic Airways Inflammation with Airway Remodeling. Sci. Rep. 2018, 8, 6925. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.L.; Levin, M.E.; Zar, H.J.; Potter, P.C.; Khumalo, N.P.; Volkwyn, L.; Fenemore, B.; du Toit, G. Food Allergy in South African Children with Atopic Dermatitis. Pediatr. Allergy Immunol. 2014, 25, 572–579. [Google Scholar] [CrossRef]
- Canonica, G.W.; Senna, G.; Mitchell, P.D.; O’Byrne, P.M.; Passalacqua, G.; Varricchi, G. Therapeutic Interventions in Severe Asthma. World Allergy Organ. J. 2016, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Dvorin, E.L.; Ebell, M.H. Short-Term Systemic Corticosteroids: Appropriate Use in Primary Care. Am. Fam. Physician 2020, 101, 89–94. [Google Scholar]
- Venekamp, R.P.; Thompson, M.J.; Hayward, G.; Heneghan, C.J.; Mar, C.B.D.; Perera, R.; Glasziou, P.P.; Rovers, M.M. Systemic Corticosteroids for Acute Sinusitis. Cochrane Database Syst. Rev. 2014. [Google Scholar] [CrossRef] [Green Version]
- Aasbjerg, K.; Torp-Pedersen, C.; Vaag, A.; Backer, V. Treating Allergic Rhinitis with Depot-Steroid Injections Increase Risk of Osteoporosis and Diabetes. Respir. Med. 2013, 107, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Hox, V.; Lourijsen, E.; Jordens, A.; Aasbjerg, K.; Agache, I.; Alobid, I.; Bachert, C.; Boussery, K.; Campo, P.; Fokkens, W.; et al. Benefits and Harm of Systemic Steroids for Short- and Long-Term Use in Rhinitis and Rhinosinusitis: An EAACI Position Paper. Clin. Transl. Allergy 2020, 10, 1. [Google Scholar] [CrossRef]
- Choo, K.J.L.; Simons, E.; Sheikh, A. Glucocorticoids for the Treatment of Anaphylaxis: Cochrane Systematic Review. Allergy 2010, 65, 1205–1211. [Google Scholar] [CrossRef]
- Ring, J.; Gutermuth, J. 100 Years of Hyposensitization: History of Allergen-Specific Immunotherapy (ASIT). Allergy 2011, 66, 713–724. [Google Scholar] [CrossRef]
- Durham, S.R.; Nelson, H. Allergen Immunotherapy: A Centenary Celebration. World Allergy Organ. J. 2011, 4, 104–106. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, M.C.; Sur, S. Down Regulation of the High-Affinity IgE Receptor Associated with Successful Treatment of Chronic Idiopathic Urticaria with Omalizumab. Clin. Mol. Allergy CMA 2011, 9, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagnasco, D.; Ferrando, M.; Varricchi, G.; Passalacqua, G.; Canonica, G.W. A Critical Evaluation of Anti-IL-13 and Anti-IL-4 Strategies in Severe Asthma. Int. Arch. Allergy Immunol. 2016, 170, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Le Floch, A.; Allinne, J.; Nagashima, K.; Scott, G.; Birchard, D.; Asrat, S.; Bai, Y.; Lim, W.K.; Martin, J.; Huang, T.; et al. Dual Blockade of IL-4 and IL-13 with Dupilumab, an IL-4Rα Antibody, Is Required to Broadly Inhibit Type 2 Inflammation. Allergy 2020, 75, 1188–1204. [Google Scholar] [CrossRef] [PubMed]
- Ghassemian, A.; Park, J.J.; Tsoulis, M.W.; Kim, H. Targeting the IL-5 Pathway in Eosinophilic Asthma: A Comparison of Mepolizumab to Benralizumab in the Reduction of Peripheral Eosinophil Counts. Allergy Asthma Clin. Immunol. 2021, 17, 3. [Google Scholar] [CrossRef]
- Damask, C.C.; Ryan, M.W.; Casale, T.B.; Castro, M.; Franzese, C.B.; Lee, S.E.; Levy, J.M.; Lin, S.Y.; Lio, P.A.; Peters, A.T.; et al. Targeted Molecular Therapies in Allergy and Rhinology. Otolaryngol.--Head Neck Surg. Off. J. Am. Acad. Otolaryngol.-Head Neck Surg. 2021, 164, S1–S21. [Google Scholar] [CrossRef]
- Landolina, N.; Levi-Schaffer, F. Monoclonal Antibodies: The New Magic Bullets for Allergy: IUPHAR Review 17. Br. J. Pharmacol. 2016, 173, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Laffer, S.; Lupinek, C.; Rauter, I.; Kneidinger, M.; Drescher, A.; Jordan, J.-H.; Krauth, M.-T.; Valent, P.; Kricek, F.; Spitzauer, S.; et al. A High-Affinity Monoclonal Anti-IgE Antibody for Depletion of IgE and IgE-Bearing Cells. Allergy 2008, 63, 695–702. [Google Scholar] [CrossRef]
- Chen, H.Y.; Liu, F.-T.; Hou, C.M.H.; Huang, J.S.W.; Sharma, B.B.; Chang, T.W. Monoclonal Antibodies against the C(Epsilon)MX Domain of Human Membrane-Bound IgE and Their Potential Use for Targeting IgE-Expressing B Cells. Int. Arch. Allergy Immunol. 2002, 128, 315–324. [Google Scholar] [CrossRef]
- Lupinek, C.; Derfler, K.; Lee, S.; Prikoszovich, T.; Movadat, O.; Wollmann, E.; Cornelius, C.; Weber, M.; Fröschl, R.; Selb, R.; et al. Extracorporeal IgE Immunoadsorption in Allergic Asthma: Safety and Efficacy. EBioMedicine 2017, 17, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Rajakulendran, M.; Tham, E.H.; Soh, J.Y.; Van Bever, H.P. Novel Strategies in Immunotherapy for Allergic Diseases. Asia Pac. Allergy 2018, 8, e14. [Google Scholar] [CrossRef]
- He, S.; Zhang, H.; Zeng, X.; Chen, D.; Yang, P. Mast Cells and Basophils Are Essential for Allergies: Mechanisms of Allergic Inflammation and a Proposed Procedure for Diagnosis. Acta Pharmacol. Sin. 2013, 34, 1270–1283. [Google Scholar] [CrossRef] [Green Version]
- Kamijo, S.; Takeda, H.; Tokura, T.; Suzuki, M.; Inui, K.; Hara, M.; Matsuda, H.; Matsuda, A.; Oboki, K.; Ohno, T. IL-33–Mediated Innate Response and Adaptive Immune Cells Contribute to Maximum Responses of Protease Allergen–Induced Allergic Airway Inflammation. J. Immunol. 2013, 190, 4489–4499. [Google Scholar] [CrossRef] [Green Version]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Dittrich, A.M. Expression and Function of Histamine and Its Receptors in Atopic Dermatitis. Mol. Cell. Pediatr. 2015, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Bakker, R.A.; Schoonus, S.B.; Smit, M.J.; Timmerman, H.; Leurs, R. Histamine H1-Receptor Activation of Nuclear Factor-ΚB: Roles for Gβγ-and Gαq/11-Subunits in Constitutive and Agonist-Mediated Signaling. Mol. Pharmacol. 2001, 60, 1133–1142. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast Cell Secretory Granules: Armed for Battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Buckland, K.F.; Williams, T.J.; Conroy, D.M. Histamine Induces Cytoskeletal Changes in Human Eosinophils via the H4 Receptor. Br. J. Pharmacol. 2003, 140, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Seifert, R.; Strasser, A.; Schneider, E.H.; Neumann, D.; Dove, S.; Buschauer, A. Molecular and Cellular Analysis of Human Histamine Receptor Subtypes. Trends Pharmacol. Sci. 2013, 34, 33–58. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Jadhav, H.R. Histamine H3 Receptor Function and Ligands: Recent Developments. Mini Rev. Med. Chem. 2013, 13, 47–57. [Google Scholar] [CrossRef]
- Lippert, U.; Artuc, M.; Grützkau, A.; Babina, M.; Guhl, S.; Haase, I.; Blaschke, V.; Zachmann, K.; Knosalla, M.; Middel, P.; et al. Human Skin Mast Cells Express H2 and H4, but Not H3 Receptors. J. Investig. Dermatol. 2004, 123, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, X.; Qian, Q.; Wang, Y.; Dong, H.; Li, N.; Qian, Y.; Jin, W. Histamine Upregulates the Expression of Histamine Receptors and Increases the Neuroprotective Effect of Astrocytes. J. Neuroinflammation 2018, 15, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theoharides, T.C.; Tsilioni, I.; Bawazeer, M. Mast Cells, Neuroinflammation and Pain in Fibromyalgia Syndrome. Front. Cell. Neurosci. 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhang, X.; Wang, Y.; Zhou, X.; Qian, Y.; Zhang, S. Suppression of Brain Mast Cells Degranulation Inhibits Microglial Activation and Central Nervous System Inflammation. Mol. Neurobiol. 2017, 54, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Dinh, Q.T.; Cryer, A.; Dinh, S.; Peiser, C.; Wu, S.; Springer, J.; Hamelmann, E.; Klapp, B.F.; Heppt, W.; Fischer, A. Transcriptional Up-Regulation of Histamine Receptor-1 in Epithelial, Mucus and Inflammatory Cells in Perennial Allergic Rhinitis. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2005, 35, 1443–1448. [Google Scholar] [CrossRef]
- Merves, J.; Chandramouleeswaran, P.M.; Benitez, A.J.; Muir, A.B.; Lee, A.J.; Lim, D.M.; Dods, K.; Mehta, I.; Ruchelli, E.D.; Nakagawa, H.; et al. Altered Esophageal Histamine Receptor Expression in Eosinophilic Esophagitis (EoE): Implications on Disease Pathogenesis. PLoS ONE 2015, 10, e0114831. [Google Scholar] [CrossRef]
- Eckman, J.A.; Sterba, P.M.; Kelly, D.; Alexander, V.; Liu, M.C.; Bochner, B.S.; Macglashan, D.W.; Saini, S.S. Effects of Omalizumab on Basophil and Mast Cell Responses Using an Intranasal Cat Allergen Challenge. J. Allergy Clin. Immunol. 2010, 125, 889–895.e7. [Google Scholar] [CrossRef] [Green Version]
- Noga, O.; Hanf, G.; Kunkel, G.; Kleine-Tebbe, J. Basophil Histamine Release Decreases during Omalizumab Therapy in Allergic Asthmatics. Int. Arch. Allergy Immunol. 2008, 146, 66–70. [Google Scholar] [CrossRef]
- MacGlashan, D.; Saini, S.S. Omalizumab Increases the Intrinsic Sensitivity of Human Basophils to IgE-Mediated Stimulation. J. Allergy Clin. Immunol. 2013, 132, 906–911.e1-4. [Google Scholar] [CrossRef] [Green Version]
- Eckl-Dorna, J.; Fröschl, R.; Lupinek, C.; Kiss, R.; Gattinger, P.; Marth, K.; Campana, R.; Mittermann, I.; Blatt, K.; Valent, P.; et al. Intranasal Administration of Allergen Increases Specific IgE Whereas Intranasal Omalizumab Does Not Increase Serum IgE Levels—A Pilot Study. Allergy 2018, 73, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Kowal, K.; Bielecki, P.; DuBuske, I.V.; DuBuske, L.M. In Vitro Impact of Anti-Immunoglobulin E Monoclonal Antibodies, Including Omalizumab on Whole Blood Basophil Histamine Release: Assessment of Direct Induction of Basophil Histamine Release and Evaluation of Modulation of Allergen-Induced Basophil Histamine Release. Allergy Asthma Proc. 2017, 38. [Google Scholar] [CrossRef]
- Wu, Y.-F.; Su, M.-W.; Chiang, B.-L.; Yang, Y.-H.; Tsai, C.-H.; Lee, Y.L. A Simple Prediction Tool for Inhaled Corticosteroid Response in Asthmatic Children. BMC Pulm. Med. 2017, 17, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rhyou, H.-I.; Nam, Y.-H. Predictive Factors of Response to Inhaled Corticosteroids in Newly Diagnosed Asthma: A Real-World Observational Study. Ann. Allergy. Asthma. Immunol. 2020, 125, 177–181. [Google Scholar] [CrossRef]
- Keskin, O.; Farzan, N.; Birben, E.; Akel, H.; Karaaslan, C.; Maitland-van der Zee, A.H.; Wechsler, M.E.; Vijverberg, S.J.; Kalayci, O. Genetic Associations of the Response to Inhaled Corticosteroids in Asthma: A Systematic Review. Clin. Transl. Allergy 2019, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Kau, A.L.; Korenblat, P.E. Anti-Interleukin 4 and 13 for Asthma Treatment in the Era of Endotypes. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 570. [Google Scholar] [CrossRef] [Green Version]
- Nabe, T. Interleukin (IL)-33: New Therapeutic Target for Atopic Diseases. J. Pharmacol. Sci. 2014, 126, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Volmer, T.; Effenberger, T.; Trautner, C.; Buhl, R. Consequences of Long-Term Oral Corticosteroid Therapy and Its Side-Effects in Severe Asthma in Adults: A Focused Review of the Impact Data in the Literature. Eur. Respir. J. 2018, 52, 1800703. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, V. Asthma and Medicines–Long-Term Side-Effects, Monitoring and Dose Titration. Indian, J. Pediatr. 2018, 85, 748–756. [Google Scholar] [CrossRef]
- Reddel, H.K.; FitzGerald, J.M.; Bateman, E.D.; Bacharier, L.B.; Becker, A.; Brusselle, G.; Buhl, R.; Cruz, A.A.; Fleming, L.; Inoue, H. GINA 2019: A Fundamental Change in Asthma Management: Treatment of Asthma with Short-Acting Bronchodilators Alone Is No Longer Recommended for Adults and Adolescents. Eur. Respir. J. 2019, 53, 1901046. [Google Scholar] [CrossRef] [Green Version]
- Eff, A.R.Y. Incidence of Hypertension in Asthma Patients Who Treated with Beta-2 Agonists Bronchodilators. Int J Pharm Pharm Sci 2017, 9, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.-L.; Wang, H.H.; Lin, C.-H. Effect of Allergic Phenotype on Treatment Response to Inhaled Bronchodilators with or without Inhaled Corticosteroids in Patients with COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2017, 12, 2231. [Google Scholar] [CrossRef] [Green Version]
- Amegadzie, J.E.; Gorgui, J.; Acheampong, L.; Gamble, J.-M.; Farrell, J.; Gao, Z. Comparative Safety and Effectiveness of Inhaled Bronchodilators and Corticosteroids for Treating Asthma–COPD Overlap: A Systematic Review and Meta-Analysis. J. Asthma 2021, 58, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, C.; Corcione, N.; Rea, G.; Stefanelli, F.; Meoli, I.; Vatrella, A. Impact of Long-Term Treatment with Inhaled Corticosteroids and Bronchodilators on Lung Function in a Patient with Post-Infectious Bronchiolitis Obliterans. J. Bras. Pneumol. 2016, 42, 228–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onay, Z.R.; Gursoy, T.R.; Aslan, A.T.; Eyuboglu, T.S.; Kibar, B.S.; Pekcan, S.; Hangul, M.; Kose, M.; Budakoglu, I.İ.; Gokturk, B. Misdiagnosis of Asthma May Delay the Post Infectious Bronchiolitis Obliterans Diagnosis. Eur. Respir. J. 2019, 54, PA1015. [Google Scholar]
- Buhl, R.; FitzGerald, J.M.; Busse, W.W. Tiotropium Add-on to Inhaled Corticosteroids versus Addition of Long-Acting Β2-Agonists for Adults with Asthma. Respir. Med. 2018, 143, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Kupczyk, M.; Wenzel, S. US and European Severe Asthma Cohorts: What Can They Teach Us about Severe Asthma? J. Intern. Med. 2012, 272, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Pike, K.C.; Levy, M.L.; Moreiras, J.; Fleming, L. Managing Problematic Severe Asthma: Beyond the Guidelines. Arch. Dis. Child. 2018, 103, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Busse, W.; Corren, J.; Lanier, B.Q.; McAlary, M.; Fowler-Taylor, A.; Cioppa, G.D.; van As, A.; Gupta, N. Omalizumab, Anti-IgE Recombinant Humanized Monoclonal Antibody, for the Treatment of Severe Allergic Asthma. J. Allergy Clin. Immunol. 2001, 108, 184–190. [Google Scholar] [CrossRef]
- Deleanu, D.; Nedelea, I. Biological Therapies for Atopic Dermatitis: An Update. Exp. Ther. Med. 2019, 17, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Holgate, S.; Bousquet, J.; Wenzel, S.; Fox, H.; Liu, J.; Castellsague, J. Efficacy of Omalizumab, an Anti-Immunoglobulin E Antibody, in Patients with Allergic Asthma at High Risk of Serious Asthma-Related Morbidity and Mortality. Curr. Med. Res. Opin. 2001, 17, 233–240. [Google Scholar] [CrossRef]
- Fahy, J.V.; Fleming, H.E.; Wong, H.H.; Liu, J.T.; Su, J.Q.; Reimann, J.; Fick, R.B., Jr.; Boushey, H.A. The Effect of an Anti-IgE Monoclonal Antibody on the Early- and Late-Phase Responses to Allergen Inhalation in Asthmatic Subjects. Am. J. Respir. Crit. Care Med. 1997, 155. [Google Scholar] [CrossRef] [Green Version]
- Trischler, J.; Lieb, A.; Arnold, M.; Schulze, J.; Rosewich, M.; Schubert, R.; Bottoli, I.; Zielen, S. Omalizumab Effectively Protects against Early and Late Allergic Responses in Asthma after 4 Weeks. Allergy 2017, 72, 1912–1915. [Google Scholar] [CrossRef]
- Pelaia, G.; Canonica, G.W.; Matucci, A.; Paolini, R.; Triggiani, M.; Paggiaro, P. Targeted Therapy in Severe Asthma Today: Focus on Immunoglobulin, E. Drug Des. Devel. Ther. 2017, 11, 1979. [Google Scholar] [CrossRef] [Green Version]
- Esquivel, A.; Busse, W.W.; Calatroni, A.; Togias, A.G.; Grindle, K.G.; Bochkov, Y.A.; Gruchalla, R.S.; Kattan, M.; Kercsmar, C.M.; Khurana Hershey, G. Effects of Omalizumab on Rhinovirus Infections, Illnesses, and Exacerbations of Asthma. Am. J. Respir. Crit. Care Med. 2017, 196, 985–992. [Google Scholar] [CrossRef]
- Acer, E.; Kaya Erdogan, H.; Yüksel Çanakçı, N.; Saracoglu, Z.N. The Effect of Omalizumab on Hematological and Inflammatory Parameters in Patients with Chronic Spontaneous Urticaria. Cutan. Ocul. Toxicol. 2019, 38, 5–8. [Google Scholar] [CrossRef]
- Masieri, S.; Cavaliere, C.; Begvarfaj, E.; Rosati, D.; Minni, A. Effects of Omalizumab Therapy on Allergic Rhinitis: A Pilot Study. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5249–5255. [Google Scholar]
- Ricciardi, L.; Papia, F.; Liotta, M.; Cicero, F.; Isola, S.; Tartarisco, G.; Furci, F.; Gangemi, S. Omalizumab in Middle-Aged or Older Patients with Severe Allergic Asthma-COPD Overlap. Adv. Dermatol. Allergol. Dermatol. Alergol. 2022, 39, 88–93. [Google Scholar] [CrossRef]
- Heck, S.; Nguyen, J.; Le, D.-D.; Bals, R.; Dinh, Q.T. Pharmacological Therapy of Bronchial Asthma: The Role of Biologicals. Int. Arch. Allergy Immunol. 2015, 168, 241–252. [Google Scholar] [CrossRef]
- Maurer, M.; Giménez-Arnau, A.M.; Sussman, G.; Metz, M.; Baker, D.R.; Bauer, A.; Bernstein, J.A.; Brehler, R.; Chu, C.-Y.; Chung, W.-H. Ligelizumab for Chronic Spontaneous Urticaria. N. Engl. J. Med. 2019, 381, 1321–1332. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Chen, J.; Ye, L.; Cai, Z.; Sun, J.; Ji, K. Anti-IgE Therapy for IgE-Mediated Allergic Diseases: From Neutralizing IgE Antibodies to Eliminating IgE+ B Cells. Clin. Transl. Allergy 2018, 8, 27. [Google Scholar] [CrossRef]
- Wedi, B. Ligelizumab for the Treatment of Chronic Spontaneous Urticaria. Expert Opin. Biol. Ther. 2020, 20, 853–861. [Google Scholar] [CrossRef]
- Wedi, B.; Traidl, S. Anti-IgE for the Treatment of Chronic Urticaria. ImmunoTargets Ther. 2021, 10, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.S.; Dobson, C.L.; Käck, H.; Wang, B.; Sims, D.A.; Lloyd, C.O.; England, E.; Rees, D.G.; Guo, H.; Karagiannis, S.N. A Novel IgE-Neutralizing Antibody for the Treatment of Severe Uncontrolled Asthma. mAbs 2014, 6, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, E.; Schwickart, M.; Li, J.; Kim, K.; Crouch, S.; Parveen, S.; Kell, C.; Birrell, C. Pharmacokinetics, Pharmacodynamics, and Safety of MEDI4212, an Anti-IgE Monoclonal Antibody, in Subjects with Atopy: A Phase I Study. Adv. Ther. 2016, 33, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Scheerens, H.; Arron, J.R.; Zheng, Y.; Putnam, W.S.; Erickson, R.W.; Choy, D.F.; Harris, J.M.; Lee, J.; Jarjour, N.N.; Matthews, J.G. The Effects of Lebrikizumab in Patients with Mild Asthma Following Whole Lung Allergen Challenge. Clin. Exp. Allergy 2014, 44, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.M.; Cabanski, C.R.; Scheerens, H.; Samineni, D.; Bradley, M.S.; Cochran, C.; Staubach, P.; Metz, M.; Sussman, G.; Maurer, M. A Randomized Trial of Quilizumab in Adults with Refractory Chronic Spontaneous Urticaria. J. Allergy Clin. Immunol. 2016, 138, 1730–1732. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.M.; Maciuca, R.; Bradley, M.S.; Cabanski, C.R.; Scheerens, H.; Lim, J.; Cai, F.; Kishnani, M.; Liao, X.C.; Samineni, D. A Randomized Trial of the Efficacy and Safety of Quilizumab in Adults with Inadequately Controlled Allergic Asthma. Respir. Res. 2016, 17, 29. [Google Scholar] [CrossRef] [Green Version]
- Kirak, O.; Riethmüller, G. A Novel, Nonanaphylactogenic, Bispecific IgE-CD3 Antibody Eliminates IgE+ B Cells. J. Allergy Clin. Immunol. 2015, 136, 800–802. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.Y.; Horton, H.M.; Pong, E.; Leung, I.W.; Chen, H.; Nguyen, D.-H.; Bautista, C.; Muchhal, U.S.; Bernett, M.J.; Moore, G.L. Reduction of Total IgE by Targeted Coengagement of IgE B-Cell Receptor and FcγRIIb with Fc-Engineered Antibody. J. Allergy Clin. Immunol. 2012, 129, 1102–1115. [Google Scholar] [CrossRef]
- Saggini, A.; Maccauro, G.; Tripodi, D.; De Lutiis, M.A.; Conti, F.; Felaco, P.; Fulcheri, M.; Galzio, R.; Caraffa, A.; Antinolfi, P. Allergic Inflammation: Role of Cytokines with Special Emphasis on IL-4; SAGE Publications Sage UK: London, UK, 2011. [Google Scholar]
- Corry, D.B.; Folkesson, H.G.; Warnock, M.L.; Erle, D.J.; Matthay, M.A.; Wiener-Kronish, J.P.; Locksley, R.M. Interleukin 4, but Not Interleukin 5 or Eosinophils, Is Required in a Murine Model of Acute Airway Hyperreactivity. J. Exp. Med. 1996, 183, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Coyle, A.J.; Le Gros, G.; Bertrand, C.; Tsuyuki, S.; Heusser, C.H.; Kopf, M.; Anderson, G.P. Interleukin-4 Is Required for the Induction of Lung Th2 Mucosal Immunity. Am. J. Respir. Cell Mol. Biol. 1995, 13, 54–59. [Google Scholar] [CrossRef]
- Akdis, C.A. Therapies for Allergic Inflammation: Refining Strategies to Induce Tolerance. Nat. Med. 2012, 18, 736–749. [Google Scholar] [CrossRef]
- Vladich, F.D.; Brazille, S.M.; Stern, D.; Peck, M.L.; Ghittoni, R.; Vercelli, D. IL-13 R130Q, a Common Variant Associated with Allergy and Asthma, Enhances Effector Mechanisms Essential for Human Allergic Inflammation. J. Clin. Investig. 2005, 115, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Walter, D.M.; McIntire, J.J.; Berry, G.; McKenzie, A.N.; Donaldson, D.D.; DeKruyff, R.H.; Umetsu, D.T. Critical Role for IL-13 in the Development of Allergen-Induced Airway Hyperreactivity. J. Immunol. 2001, 167, 4668–4675. [Google Scholar] [CrossRef] [Green Version]
- Wang, I.-M.; Lin, H.; Goldman, S.J.; Kobayashi, M. STAT-1 Is Activated by IL-4 and IL-13 in Multiple Cell Types. Mol. Immunol. 2004, 41, 873–884. [Google Scholar] [CrossRef]
- Wollenberg, A.; Howell, M.D.; Guttman-Yassky, E.; Silverberg, J.I.; Kell, C.; Ranade, K.; Moate, R.; van der Merwe, R. Treatment of Atopic Dermatitis with Tralokinumab, an Anti–IL-13 MAb. J. Allergy Clin. Immunol. 2019, 143, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Bagnasco, D.; Caminati, M.; Ferrando, M.; Aloè, T.; Testino, E.; Canonica, G.W.; Passalacqua, G. Anti-IL-5 and IL-5Ra: Efficacy and Safety of New Therapeutic Strategies in Severe Uncontrolled Asthma. BioMed Res. Int. 2018, 2018, 5698212. [Google Scholar] [CrossRef]
- Lawrence, M.G.; Steinke, J.W.; Borish, L. Cytokine-Targeting Biologics for Allergic Diseases. Ann. Allergy. Asthma. Immunol. 2018, 120, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Wegmann, M. Targeting Cytokines in Asthma Therapy: Could IL-37 Be a Solution? Expert Rev. Respir. Med. 2017, 11, 675–677. [Google Scholar] [CrossRef] [Green Version]
- Kasaian, M.T.; Marquette, K.; Fish, S.; DeClercq, C.; Agostinelli, R.; Cook, T.A.; Brennan, A.; Lee, J.; Fitz, L.; Brooks, J. An IL-4/IL-13 Dual Antagonist Reduces Lung Inflammation, Airway Hyperresponsiveness, and IgE Production in Mice. Am. J. Respir. Cell Mol. Biol. 2013, 49, 37–46. [Google Scholar] [CrossRef]
- Corren, J.; Castro, M.; Chanez, P.; Fabbri, L.; Joish, V.N.; Amin, N.; Graham, N.M.H.; Mastey, V.; Abbé, A.; Taniou, C.; et al. Dupilumab Improves Symptoms, Quality of Life, and Productivity in Uncontrolled Persistent Asthma. Ann. Allergy. Asthma. Immunol. 2019, 122, 41–49.e2. [Google Scholar] [CrossRef] [Green Version]
- Olin, J.T.; Wechsler, M.E. Asthma: Pathogenesis and Novel Drugs for Treatment. BMJ 2014, 349, g5517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonstam, K.; Swanson, B.N.; Mannent, L.P.; Cardell, L.-O.; Tian, N.; Wang, Y.; Zhang, D.; Fan, C.; Holtappels, G.; Hamilton, J.D.; et al. Dupilumab Reduces Local Type 2 Pro-Inflammatory Biomarkers in Chronic Rhinosinusitis with Nasal Polyposis. Allergy 2019, 74, 743–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, W.; Yuan, H.; Li, Y.; Lv, Z.; Cui, Y.; Liu, J.; Ying, S. Current State of Monoclonal Antibody Therapy for Allergic Diseases. Engineering 2021, 7, 1552–1556. [Google Scholar] [CrossRef]
- Baldo, B.A. Adverse Events to Monoclonal Antibodies Used for Cancer Therapy. Oncoimmunology 2013, 2, e26333. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.S. How Safe Are the Biologicals in Treating Asthma and Rhinitis? Allergy Asthma Clin. Immunol. Off. J. Can. Soc. Allergy Clin. Immunol. 2009, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Hansel, T.T.; Kropshofer, H.; Singer, T.; Mitchell, J.A.; George, A.J.T. The Safety and Side Effects of Monoclonal Antibodies. Nat. Rev. Drug Discov. 2010, 9, 325–338. [Google Scholar] [CrossRef]
- Kaplan, A.P.; Murphy, T.R.; Holden, M.; Iqbal, A.; Yoo, B.; Bernstein, J.A. Impact of Omalizumab Treatment Withdrawal After 24 and 48 Weeks in Patients with Chronic Idiopathic Urticaria: Results From the XTEND-CIU Study. J. Allergy Clin. Immunol. 2019, 143, AB209. [Google Scholar] [CrossRef] [Green Version]
- Nopp, A.; Johansson, S.G.O.; Adédoyin, J.; Ankerst, J.; Palmqvist, M.; Oman, H. After 6 Years with Xolair; a 3-Year Withdrawal Follow-Up. Allergy 2010, 65, 56–60. [Google Scholar] [CrossRef]
- Wu, A.C.; Fuhlbrigge, A.L.; Robayo, M.A.; Shaker, M. Cost-Effectiveness of Biologics for Allergic Diseases. J. Allergy Clin. Immunol. Pract. 2021, 9, 1107–1117. [Google Scholar] [CrossRef]
- Samaranayake, H.; Wirth, T.; Schenkwein, D.; Räty, J.K.; Ylä-Herttuala, S. Challenges in Monoclonal Antibody-Based Therapies. Ann. Med. 2009, 41, 322–331. [Google Scholar] [CrossRef]
- Zimmermann, M.; Rind, D.; Chapman, R.; Kumar, V.; Kahn, S.; Carlson, J. Economic Evaluation of Dupilumab for Moderate-to-Severe Atopic Dermatitis: A Cost-Utility Analysis. J. Drugs Dermatol. JDD 2018, 17, 750–756. [Google Scholar]
- Mingozzi, F.; High, K.A. Immune Responses to AAV Vectors: Overcoming Barriers to Successful Gene Therapy. Blood 2013, 122, 23–36. [Google Scholar] [CrossRef]
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-Associated Virus Vector as a Platform for Gene Therapy Delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. Biodrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, W.; Zhu, W.; Chen, J.; Ma, B.; Ding, J.; Wang, Z.; Li, Y.; Wang, Y.; Zhang, X. Adeno-associated Virus (AAV)-Based Gene Therapy for Glioblastoma. Cancer Cell Int. 2021, 21, 76. [Google Scholar] [CrossRef]
- Camilleri, A.E.; Nag, S.; Russo, A.R.; Stiles, K.M.; Crystal, R.G.; Pagovich, O.E. Gene Therapy for a Murine Model of Eosinophilic Esophagitis. Allergy 2021, 76, 2740–2752. [Google Scholar] [CrossRef]
- Pagovich, O.E.; Wang, B.; Chiuchiolo, M.J.; Kaminsky, S.M.; Sondhi, D.; Jose, C.L.; Price, C.C.; Brooks, S.F.; Mezey, J.G.; Crystal, R.G. Anti-HIgE Gene Therapy of Peanut-Induced Anaphylaxis in a Humanized Murine Model of Peanut Allergy. J. Allergy Clin. Immunol. 2016, 138, 1652–1662.e7. [Google Scholar] [CrossRef] [Green Version]
- Zavorotinskaya, T.; Tomkinson, A.; Murphy, J.E. Treatment of Experimental Asthma by Long-Term Gene Therapy Directed against IL-4 and IL-13. Mol. Ther. J. Am. Soc. Gene Ther. 2003, 7, 155–162. [Google Scholar] [CrossRef]
- Nault, J.-C.; Datta, S.; Imbeaud, S.; Franconi, A.; Mallet, M.; Couchy, G.; Letouzé, E.; Pilati, C.; Verret, B.; Blanc, J.-F.; et al. Recurrent AAV2-Related Insertional Mutagenesis in Human Hepatocellular Carcinomas. Nat. Genet. 2015, 47, 1187–1193. [Google Scholar] [CrossRef]
- Rogers, G.L.; Suzuki, M.; Zolotukhin, I.; Markusic, D.M.; Morel, L.M.; Lee, B.; Ertl, H.C.J.; Herzog, R.W. Unique Roles of TLR9- and MyD88-Dependent and -Independent Pathways in Adaptive Immune Responses to AAV-Mediated Gene Transfer. J. Innate Immun. 2015, 7, 302–314. [Google Scholar] [CrossRef]
- Manno, C.S.; Pierce, G.F.; Arruda, V.R.; Glader, B.; Ragni, M.; Rasko, J.J.; Rasko, J.; Ozelo, M.C.; Hoots, K.; Blatt, P.; et al. Successful Transduction of Liver in Hemophilia by AAV-Factor IX and Limitations Imposed by the Host Immune Response. Nat. Med. 2006, 12, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Nidetz, N.F.; McGee, M.C.; Tse, L.V.; Li, C.; Cong, L.; Li, Y.; Huang, W. Adeno-Associated Viral Vector-Mediated Immune Responses: Understanding Barriers to Gene Delivery. Pharmacol. Ther. 2020, 207, 107453. [Google Scholar] [CrossRef] [PubMed]
- Thran, M.; Mukherjee, J.; Pönisch, M.; Fiedler, K.; Thess, A.; Mui, B.L.; Hope, M.J.; Tam, Y.K.; Horscroft, N.; Heidenreich, R.; et al. MRNA Mediates Passive Vaccination against Infectious Agents, Toxins, and Tumors. EMBO Mol. Med. 2017, 9, 1434–1447. [Google Scholar] [CrossRef] [PubMed]
- Van Hoecke, L.; Roose, K. How MRNA Therapeutics Are Entering the Monoclonal Antibody Field. J. Transl. Med. 2019, 17, 54. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Secreto, A.J.; Shan, X.; Debonera, F.; Glover, J.; Yi, Y.; Muramatsu, H.; Ni, H.; Mui, B.L.; Tam, Y.K.; et al. Administration of Nucleoside-Modified MRNA Encoding Broadly Neutralizing Antibody Protects Humanized Mice from HIV-1 Challenge. Nat. Commun. 2017, 8, 14630. [Google Scholar] [CrossRef] [Green Version]
- Deal, C.E.; Carfi, A.; Plante, O.J. Advancements in MRNA Encoded Antibodies for Passive Immunotherapy. Vaccines 2021, 9, 108. [Google Scholar] [CrossRef]
- Schlake, T.; Thran, M.; Fiedler, K.; Heidenreich, R.; Petsch, B.; Fotin-Mleczek, M. MRNA: A Novel Avenue to Antibody Therapy? Mol. Ther. 2019, 27, 773–784. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Receptors | Effects | Clinical Manifestation | References |
|---|---|---|---|
| H1 | Stimulates nociceptive nerve fiber; bronchoconstriction; increase mucus secretion | Itching; urticaria; allergic rhinoconjunctivitis; allergic asthma | [33,35] |
| H2 | Gastric acid secretion; arrhythmia; increase intracellular cAMP | Peptic ulcer anaphylaxis; negative feedback on mast cell activation | [33,38] |
| H3 | Inflammation on the neurons | Neuro-inflammatory diseases such as epilepsy; CNS disorder | [33,39] |
| H4 | Induce eosinophil shape change; activates eosinophils | Lung asthma | [37] |
| Candidates | Type, Origin, and Target | Major Findings | References |
|---|---|---|---|
| MEDI4212 | Human IgG1λ mAb, an anti-IgE | Reduces both serum IgE and FcεR1 expression on dendritic cells and basophil in patients with atopy; Prevent IgE to FcεRII interaction in asthma. | [82,83] |
| Quilizumab | Humanized IgG1 mAb, an anti-IgE+ B cell | Reduces serum IgE in patients with asthma, allergic rhinitis, and chronic spontaneous urticaria; No significant impact on lung function; Not better than omalizumab. | [84,85,86] |
| Bsc-IgE/CD3 | Anti-IgE+ B cells | Eliminates serum IgE, B-IgE+ cells via cytotoxicity; Prevent mast cell degranulation in rat basophilic cell lines (RBL-2H3); May be useful in treating IgE-mediated allergic disorders. | [79,87] |
| XmAb7195 | Humanized mAb; FcγRIIb-enhanced Fc | Inhibits plasma cell differentiation; Reduces total human IgE 40-fold relative to omalizumab. | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ezechukwu, H.C.; Adegboye, O.A.; Okunowo, W.O.; Emeto, T.I. Targeting IgE and Th2-Cytokines in Allergy: Brief Updates on Monoclonal Antibodies and Antibody Gene Therapy. Allergies 2023, 3, 90-104. https://doi.org/10.3390/allergies3020007
Ezechukwu HC, Adegboye OA, Okunowo WO, Emeto TI. Targeting IgE and Th2-Cytokines in Allergy: Brief Updates on Monoclonal Antibodies and Antibody Gene Therapy. Allergies. 2023; 3(2):90-104. https://doi.org/10.3390/allergies3020007
Chicago/Turabian StyleEzechukwu, Henry C., Oyelola A. Adegboye, Wahab O. Okunowo, and Theophilus I. Emeto. 2023. "Targeting IgE and Th2-Cytokines in Allergy: Brief Updates on Monoclonal Antibodies and Antibody Gene Therapy" Allergies 3, no. 2: 90-104. https://doi.org/10.3390/allergies3020007