Genetics Unveil the Genealogical Ancestry and Physical Appearance of an Unknown Historical Figure: Lady Leonor of Castile (Spain) (1256–1275)

 , , , and
, , , and
Abstract
:1. Introduction
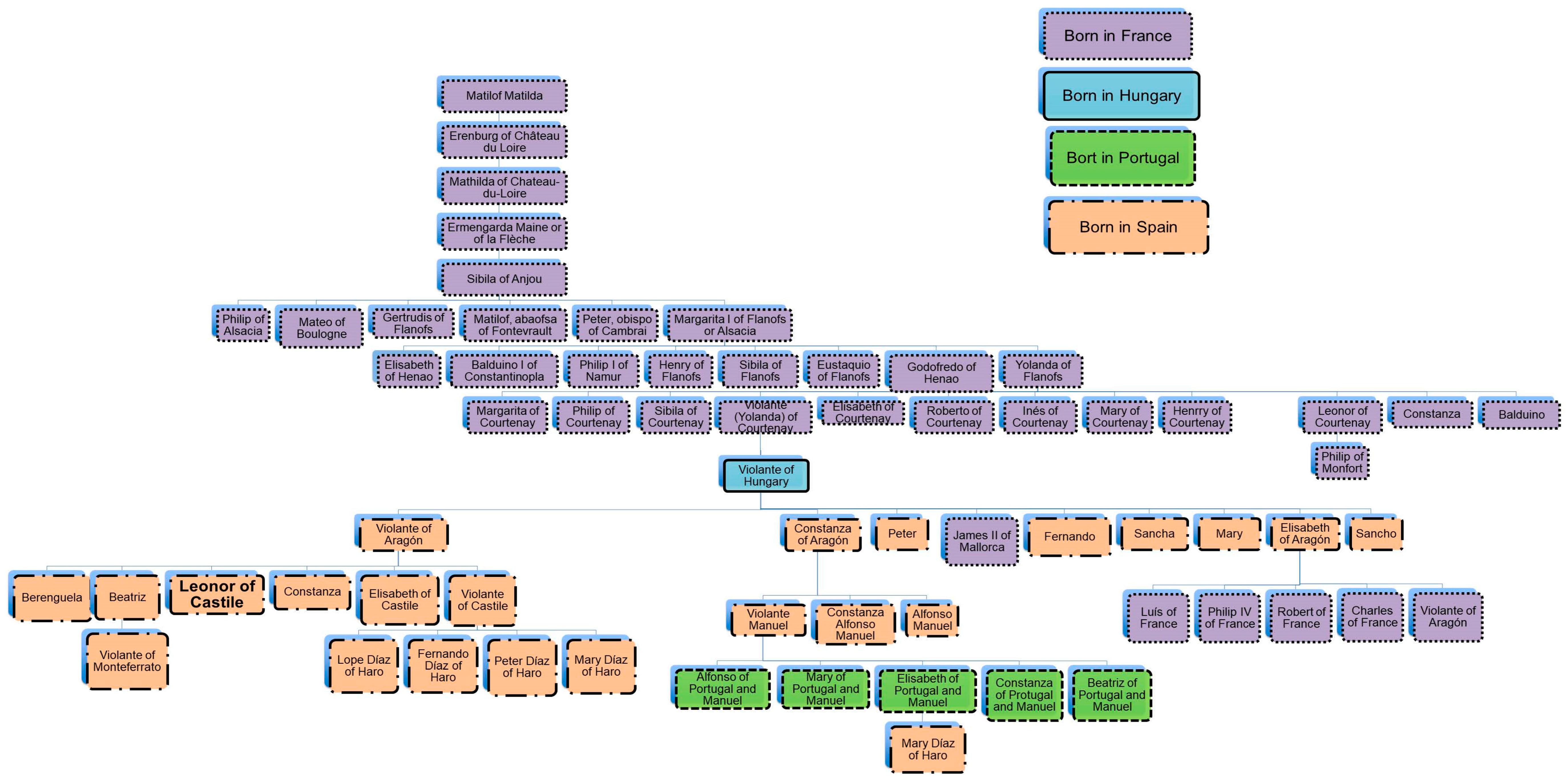
Leonor of Castile, an Unknown Princess
2. Materials and Methods
- (a)
- The amplification of the DNA regions of interest supplied us with two different kinds of information: biogeographical origin and externally visible characteristics (EVCs).
- (b)
- For the external phenotypic prediction, 35 SNPs were analyzed in Lab 2 to calculate the most probable hair, skin, and eye color pigmentation, using the primers and PCR conditions of Ruiz et al. (2013) (Maroñas et al. 2014). The EVCs PCR conditions are collected in Table 2. For each externally visible characteristic (EVC), the hypotheses considered were (a) iris pigmentation (brown, intermediate, and blue); (b) hair pigmentation (fair versus dark, and for the color pigmentation (red, blond, brown, and black); and (c) skin pigmentation (white, intermediate, and black) (Maroñas et al. 2014). Statistical analysis was performed with the online Snipper app suite v2.5 software (Phillips et al. 2007).
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alves-Cardoso, Francisca, Sara Palomo-Díez, Alejandro Alonso-Conde, Cláudia Gomes, Silvia Casimiro, Rodrigo Banha da Silva, and Eduardo Arroyo-Pardo. 2022. Metagenomic Research of Infectious Diseases in Archealogical Contexts: Evidence from the Hospital Real de Todos-os-Santos (Portugal). Applied Sciences 12: 6069. [Google Scholar] [CrossRef]
- Amorim, Carlos Eduardo G., Stefania Vai, Cosimo Posth, Alessandra Modi, István Koncz, Susanne Hakenbeck, Maria Cristina La Rocca, Balazs Mende, Dean Bobo, Walter Pohl, and et al. 2018. Understanding the 6th-century barbarian social organization and migration through paleogenomics. Nature Communications 9: 1–11. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S., Alan T. Bankier, B. G. Barrell, M. H. De Bruijn, Alan R. Coulson, Jacques Drouin, Ian C. Eperon, D. P. Nierlich, B. A. Roe, Fred Sanger, and et al. 1981. Sequence and organization of the human mitochondrial genome. Nature 290: 457–65. [Google Scholar] [CrossRef]
- Andrews, Richard M., Iwona Kubacka, Patrick F. Chinnery, Robert N. Lightowlers, Douglass M. Turnbull, and Neil Howell. 1999. Reanalysis and revisión of the Cambridge reference sequence for human mitochondrial DNA. Nature Genetics 23: 147. [Google Scholar] [CrossRef]
- Arias Guillén, Fernando. 2015. Enterramientos regios en Castilla y León (c. 842–1504). La dispersión de los espacios funerarios y el fracaso de la memoria dinástica. Anuario de Estudios Medievales 45: 643–75. [Google Scholar] [CrossRef]
- de Salazar Acha, Jaime. 1990. «Precisiones y nuevos datos sobre el entorno familiar de Alfonso X el Sabio, fundador de Ciudad Real». Cuaderno de estudios manchegos 20: 211–31. [Google Scholar]
- Fernández, Eva. 2005. Polimorfismos de ADN mitocondrial en poblaciones antiguas de la cuenca mediterránea. Ph.D. thesis, Universidad Central de Barcelona, Barcelona, Spain. [Google Scholar]
- Fernandez, Eva, Alejandro Pérez-Pérez, Cristina Gamba, Eva Prats, Pedro Cuesta, Josep Anfruns, Miquel Molist, Eduardo Arroyo-Pardo, and Daniel Turbón. 2014. Ancient DNA analysis of 8000 B.C. near eastern farmers supports an early neolithic pioneer maritime colonization of Mainland Europe through Cyprus and the Aegean Islands. PLoS Genetics 10: e1004401. [Google Scholar] [CrossRef]
- Fondevila, Manuel, Christopher Phillips, Cristina Santos, Ana Freire Aradas, P. M. Vallone, John M. Butler, María Victoria Lareu, and Ángel Carracedo. 2013. Revision of the SNPforID 34-plex forensic ancestry test: Assay enhancements, standard reference sample genotypes and extended population studies. Forensic Science International: Genetics 7: 63–74. [Google Scholar] [CrossRef]
- Gamba, Cristina, Eva Fernandez, Mirian Tirado, Francisco Pastor, and Eduardo Arroyo-Pardo. 2011. Ancient Nuclear DNA and Kinship Analysis: The Case of a Medieval Burial in San Esteban Church in Cuellar (Segovia, Central Spain). American Journal of Physical Anthropology 144: 485–91. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Gerard Remolins, Ana María López-Parra, Juan Francisco Gibaja, M. Fondeliva, Francesco De Angelis, V. Veltre, María Eulália Subirá, Carlos Baeza-Richer, D. Guerrero, and et al. 2020. Paleogenetic evidence of a Pyrenean Neolithic family: Kinship, Physical appearance, and biogeography multidisciplinary analysis. Journal of Archaeological Science 123: 105226. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Manuel Fondevila, Concepción Magaña-Loarte, J. Fernández-Jiménez, José Fernández-Serrano, Sara Palomo-Díez, Carlos Baeza-Richer, Ana María López-Parra, and Eduardo Arroyo-Pardo. 2019. An unusual kinship case from the Spanish Civil War (1936–1939): Ancient versus degraded sample’s investigation. Forensic Science International: Genetics Supplement Series 7: 690–91. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Manuel Fondevila, Sara Palomo-Díez, Antonio F. Pardiñas, César López-Matayoshi, Carlos Baeza-Richer, Ana. María López-Parra, María Victoria Lareu, B. López, and Eduardo Arroyo-Pardo. 2017. Phenotyping the ancient world: The physical appearance and ancestry of very degraded samples form a chalcolithic human remains. Forensic Science International: Supplement Series 6: e484–e486. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Sara Palomo-Díez, Joan Roig, Ana López-Parra, Carlos Baeza-Richer, Angel Espara-Arroyo, Juan Gibaja, and Eduardo Arroyo-Pardo. 2015. Nondestructive extraction DNA method from bones or teeth, true or false? Forensic Science International: Genetics Supplement Series 5: e279–e282. [Google Scholar] [CrossRef]
- González González, Carmen, and Cándido Aniz Iriarte. 1993. Real Monasterio de Santo Domingo de Caleruega, 1st ed. Salamanca: Editorial San Esteban. ISBN 978-84-87557-56-9. [Google Scholar]
- Gutierrez Baños, Fernando. 2014. El sepulcro de la infanta doña Leonor, hija de Alfonso X el Sabio, en el Real Monasterio de Santo Domingo de Caleruega (Burgos). Publicaciones de la Universidad de Santiago de Compostela. Quintana No. 13 2014. Santiago: Universidad de Santiago de Compostela, pp. 185–97. ISSN 1579-7414. [Google Scholar]
- Haeusler, Martin, Cordula Haas, Sandra Lösch, Negahnaz Moghaddam, Igor M. Villa, Susan Walsh, Manfred Kayser, Roger Seiler, Frank Ruehli, Manuel Janosa, and et al. 2016. Multidisciplinary Identification of the Controversial Freedom Fighter Jörg Jenatsch, Assassinated 1639 in Chur, Switzerland. PLoS ONE 11: e0168014. [Google Scholar] [CrossRef]
- Huber, Nicole, Walter Parson, and Arne Dür. 2018. Next generation database search algorithm for forensic mitogenome analyses. Forensic Science International: Genetics 37: 204–14. [Google Scholar] [CrossRef] [PubMed]
- Larmuseau, Maarten H. D., and Martin Bodner. 2018. The biological relevance of a medieval king’s DNA. Biochemical Society Transactions 46: 1013–20. [Google Scholar] [CrossRef]
- Lee, Hwan Young, Injee Song, Eunho Ha, Sung-Bae Cho, Woo Ick Yang, and Kyoung-Jin Shin. 2008. mtDNAmanager: A Web-based tool for the management and quality analysis of mitochondrial DNA control-region sequences. BMC Bioinformatics 9: 483. [Google Scholar] [CrossRef]
- Maroñas, Olalla, Chris Phillips, Jens Söchtig, Antonio Gomez-Tato, Raquel Cruz, José Alvarez-Dios, María Casares de Cal, Yarimar Ruiz, Manuel Fondevila, Ángel Carracedo, and et al. 2014. Development of a forensic skin colour predictive test. Forensic Science International: Genetics 13: 34–44. [Google Scholar] [CrossRef] [PubMed]
- Martínez Jarreta, María Begoña. 2018. Los estudios antropológicos de los restos óseos conservados. En Panteones Reales de Aragón. Zaragoza: Gobierno de Aragón, pp. 240–53. [Google Scholar]
- Martínez Santamarta, H. Salvador, and J. Robert. 2010. Alfonso X, the Learned: A biography. Volumen 146 de Studies in the History of Christian Traditions, 1st ed. Leiden: BRILL. ISBN 9789004181472. [Google Scholar]
- Martínez-Labarga, Cristina, and Olga Rickards. 1999. La utilización del DNA antiguo en la investigación de la historia evolutiva humana. Revista Española de Antropología Biológica. León: Sociedad Española de Antropología Biológica, Universidad de León, España, vol. 20, pp. 195–213. [Google Scholar]
- Mattoso, José. 2011. Introdução: Legitimação e linhagem, e-Spania. Available online: http://journals.openedition.org/e-spania/20258 (accessed on 17 May 2019).
- Mutation Surveyor® software. n.d. (Copyright 2011–2012). Available online: http://www.softgenetics.com/mutationSurveyor.html (accessed on 9 January 2023).
- Olalde, Íñigo, Swapan Mallick, Nick Patterson, Nadin Rohland, Vanessa Villalba-Mouco, Marina Silva, Katharina Dulias, Ceiridwen Edwards, Francesca Gandini, Maria Pala, and et al. 2019. The genomic history of the Iberian Peninsula over the past 8000 years. Science 363: 1230–34. [Google Scholar] [CrossRef]
- Olasz, Judit, Verena Seidenberg, Susanne Hummel, Zoltán Szentirmay, György Szabados, Béla Melegh, and Miklós Kásler. 2019. DNA profiling of Hungarian King Béla III and other skeletal remains originating from the Royal Basilica of Székesfehérvár. Archaeological and Anthropological Sciences 11: 1345–57. [Google Scholar] [CrossRef]
- Pääbo, Svante, Hendrik Poinar, David Serre, Viviane Jaenicke-Despres, Juliane Hebler, Nadine Rohland, Melanie Kuch, Johannes Krause, Linda Vigilant, and Michael Hofreiter. 2004. Genetic analyses from ancient DNA. Annual Review of Genetics 38: 645–79. [Google Scholar] [CrossRef] [PubMed]
- Parson, Walther, and Arne Dür. 2007. EMPOP-A Forensic mtDNA database. Forensic Sci International: Genetics 1: 88–92. [Google Scholar] [CrossRef]
- Pastor, Juan Francisco, Felix De Paz, Mercedes Barbosa, Natividad García, Isabel Sanjosé, Sara Palomo-Díez, and Eduardo Arroyo-Pardo. 2021. Natural mummifications in Castile & Leon (Spain). European Journal of Anatomy 25: 221–32. [Google Scholar]
- Phillips, Chris, Antonio Salas, Juan Sánchez, Manuel Fondevila, Antonio Gómez-Tato, José AntonioAlvarez-Dios, Manuel Calaza, María Casares de Cal, David Ballard, María Victoria Lareu, and et al. 2007. Inferring ancestral origin using a single multiplex assay of ancestry-informative marker SNPs. Forensic Science International: Genetics 1: 273–80. [Google Scholar] [CrossRef] [PubMed]
- Rollo, Franco, Luca Ermini, Stefania Luciani, Isolina Marota, Cristina Olivieri, and Donata Luiselli. 2006. Fine characterization of the Iceman’s mtDNA haplogroup. American Journal of Physical Anthropology 130: 557–64. [Google Scholar] [CrossRef]
- Ruiz, Yarimar, Chris Phillips, Antonio Gomez-Tato, José Antonio Alvarez-Dios, Manuel Casares de Cal, R. Cruz, Olalla Maroñas, Jens Söchtig, Manuel Fondevila, M. J. Rodriguez-Cid, and et al. 2013. Further development of forensic eye color predictive tests. Forensic Science International: Genetics 7: 28–40. [Google Scholar] [CrossRef] [PubMed]
- Salerno, Alfredo, Giampietro Lago, Andrea Berti, Francesco Calì, Mario Mirisola, Patrizia Carta, Carmen Ferro, and Valentino Romano. 2005. mtDNA analysis of the human remains buried in the sarcophagus of Federico II. Journal of Cultural Heritage 6: 313–19. [Google Scholar] [CrossRef]
- Stolarek, Ireneusz, Anna Juras, Luiza Handschuh, M. Marcinkowska-Swojak, A. Philips, M. Zenczak, A. Debski, H. Kócka-Drenz, Janusz Piontek, Piotr Kzlowski, and et al. 2018. A mosaic genetic structure of the human population living in the South Baltic region during the Iron Age. Scientific Reports 8: 2455. [Google Scholar] [CrossRef]
- Vai, Stefania, Andrea Brunelli, Alessandra Modi, Francesca Tassi, Chiara Vergata, Elena Pilli, Martina Lari, Roberta Rosa Susca, Caterina Giostra, Luisella Pejrani Baricco, and et al. 2019. A genetic perspective on Longobard-Era migrations. European Journal of Human Genetics 27: 647–56. [Google Scholar] [CrossRef]
- van Oven, Mannis, and Manfred Kayser. 2009. Updated Comprehensive Phylogenetic Tree of Global Human Mitochondrial DNA Variation. Human Mutation 30: E386–E394. [Google Scholar] [CrossRef]
- van Oven, Mannis. 2015. PhyloTree Build 17: Growing the human mitochondrial DNA tree. Forensic Science International: Genetics Supplement Series 5: 9–11. Available online: http://www.phylotree.org (accessed on 9 January 2023). [CrossRef]
- Vargas Blanco, Rufino. 1968. En torno a la boda del Infante don Felipe de Castilla y doña Cristina, Infanta de Noruega. Boletín de la Institución Fernán González 171: 380–81. [Google Scholar]
- Veeramah, Krishna R., Andreas Rott, Melanie Groß, Lucy van Dorp, Saioa López, Karola Kirsanow, Christian Sell, Jens Blöcher, Daniel Wegmann, Vivian Link, and et al. 2018. Population genomic analysis of elongated skulls reveals extensive female-biased immigration in Early Medieval Bavaria. Proceedings of the National Academy of Sciences 115: 3494–99. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| PCR Conditions | Cycling Protocol | ||||
|---|---|---|---|---|---|
| Final Concentration | Volume (mL) | ||||
| Buffer | 1 X | 0.615 | |||
| BSA | 3.2 mg/mL | 0.615 | 95 °C | 15 min | |
| Cl2Mg | 6.3 mM | 1.615 | 95 °C | 30 s | 35 cycles |
| dNTPs | 0.625 mM | 0.4 | 60 °C | 50 s | |
| Primer | Variable | 2 | 65 °C | 40 s | |
| TaqGold | 0.5 U | 0.1 | 65 °C | 6 min | |
| DNA | n/a | 1–4 mL | |||
| PCR Conditions | Cycling Protocol | ||||
|---|---|---|---|---|---|
| Final Concentration | Volume (mL) | ||||
| buffer | 1 X | 0.625 | 95 °C | 15 min | |
| BSA | 3.2 mg/mL | 0.625 | |||
| Cl2Mg | 6.3 mM | 1.625 | 95 °C | 30 s | 35 cycles |
| dNTPs | 0.625 mM | 0.43 | 60 °C | 50 s | |
| primer | Variable | 1.5 | 65 °C | 40 s | |
| TaqGold | 0.5 U | 0.1 | |||
| DNA | n/a | 1–4 uL | 65 °C | 6 min | |
| Laboratory Staff | Mitochondrial DNA Haplotype |
|---|---|
| RESEARCHER A | 16362C 55G 150T 239C 263G 309.1C 309.2C 315.1C |
| RESEARCHER B | 263G 315.1C |
| RESEARCHER C | 16145A 16172C 16222T 16261T 16305T 73G 242T 263G 295T 309.1C 315.1C |
| RESEARCHER D | 263G 315.1C |
| RESEARCHER E | 16126C 16294T 16296T 16304C 73G 263G 315.1C |
| RESEARCHER F | 73G 263G 315.1C |
| SNP | RESULT | SNP | RESULT | SNP | RESULT |
|---|---|---|---|---|---|
| rs5997008 | C | rs773658 | C | rs2026721 | G |
| rs2304925 | GT | rs10141763 | A | rs4540055 | A |
| rs917118 | G | rs182549 | CT | rs1335873 | A |
| rs1321333 | T | rs1573020 | A | rs16891982 | C |
| rs2814778 | C | rs896788 | C | rs730570 | T |
| rs1024116 | A | rs2065160 | A | rs1886510 | NN |
| rs7897550 | C | rs2572307 | G | rs5030240 | C |
| rs722098 | AG | rs2303798 | C | rs3827760 | A |
| rs10843344 | CT | rs2065982 | A | ||
| rs12913832 | AG | rs3785181 | C | ||
| rs239031 | NN | rs881929 | NN | ||
| rs2040411 | AG | rs1498444 | A | ||
| rs1978806 | T | rs1426654 | T | ||
| LIKELIHOOD RATIO RESULTS | |||||
| HYPOTHESIS | LR | ||||
| H1: European origin vs. H2: American origin | LR = 1.95 × 1016 vs. 1 | ||||
| H1: European origin vs. H2: Asia origin | LR = 1.02 × 1018 vs. 1 | ||||
| H1: European origin vs. H2: North-African origin | LR = 10382,18 vs. 1 | ||||
| H1: European origin vs. H2: Sub-Saharian origin | LR = 2.18 × 1031 vs. 1 | ||||
| Iris Eye Color System | Hair Color System | Skyn Color System | |||
|---|---|---|---|---|---|
| SNP | RESULT | SNP | RESULT | SNP | RESULT |
| rs12913832 | AG | rs1129038 | NN | rs10777129 | NN |
| rs1129038 | NN | rs11547464 | NN | rs13289 | C |
| rs11636232 | CT | rs12913832 | AG | rs1408799 | G |
| rs12203592 | C | rs12931267 | C | rs1426654 | A |
| rs12896399 | A | rs1805006 | C | rs1448484 | A |
| rs1393350 | G | rs1805007 | NN | rs16891982 | C |
| rs1667394 | CT | rs1805008 | G | rs2402130 | A |
| rs16891982 | C | rs1805009 | G | rs3829241 | C |
| rs1800407 | C | rs28777 | NN | rs6058017 | NN |
| rs4778232 | G | rs35264875 | A | rs6119471 | C |
| rs4778241 | GT | rs4778138 | CT | ||
| rs7183877 | A | rs7495174 | CT | ||
| rs8024968 | G | ||||
| LIKELIHOOD RATIO RESULTS | |||||
| HYPOTHESIS | LR | HYPOTHESIS | LR | HYPOTHESIS | LR |
| H1: Dark hair vs. H2: Light hair | LR = 487.06 vs. 1 | H1: Green-hazel eyes vs. | LR = 26.71 vs. 1 | H1: White skin vs. | LR = 7.54 × 105 vs. 1 |
| H2: Brown eyes | H2: Intermediate skin | ||||
| H1: Black hair vs. H2: Brown hair | LR = 2.65 vs. 1 | H1: Green-hazel eyes vs. | LR = 2.64 × 106 vs. 1 | H1: White skin vs. | LR = 1.98 × 1010 vs. 1 |
| H2: Blue eyes | H2: Black skin | ||||
| H1: Black hair vs. H2: Red hair | LR = 215.63 vs. 1 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomo-Díez, S.; Gomes, C.; Fondevila, M.S.; Esparza-Arroyo, Á.; López-Parra, A.M.; Lareu, M.V.; Arroyo-Pardo, E.; Pastor, J.F. Genetics Unveil the Genealogical Ancestry and Physical Appearance of an Unknown Historical Figure: Lady Leonor of Castile (Spain) (1256–1275). Genealogy 2023, 7, 28. https://doi.org/10.3390/genealogy7020028
Palomo-Díez S, Gomes C, Fondevila MS, Esparza-Arroyo Á, López-Parra AM, Lareu MV, Arroyo-Pardo E, Pastor JF. Genetics Unveil the Genealogical Ancestry and Physical Appearance of an Unknown Historical Figure: Lady Leonor of Castile (Spain) (1256–1275). Genealogy. 2023; 7(2):28. https://doi.org/10.3390/genealogy7020028
Chicago/Turabian StylePalomo-Díez, Sara, Cláudia Gomes, María Sonia Fondevila, Ángel Esparza-Arroyo, Ana María López-Parra, María Victoria Lareu, Eduardo Arroyo-Pardo, and Juan Francisco Pastor. 2023. "Genetics Unveil the Genealogical Ancestry and Physical Appearance of an Unknown Historical Figure: Lady Leonor of Castile (Spain) (1256–1275)" Genealogy 7, no. 2: 28. https://doi.org/10.3390/genealogy7020028