
Biochemical and Yield Component of Hybrid Chili (Capsicum annuum L.) Resulting from Full Diallel Crosses


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Genetic Material
2.2. Field Experimental
2.2.1. Field Procedure
2.2.2. Field Observation
2.3. Biochemical Experimental
2.3.1. Sample Preparation
2.3.2. Total Phenolic Content (TPC)
2.3.3. Total Flavonoid Content (TFC)
2.3.4. Antioxidant Activity DPPH and FRAP Methods (DPPH and FRAP)
2.3.5. α-Glucosidase Inhibition (AGI)
2.4. Data Analysis
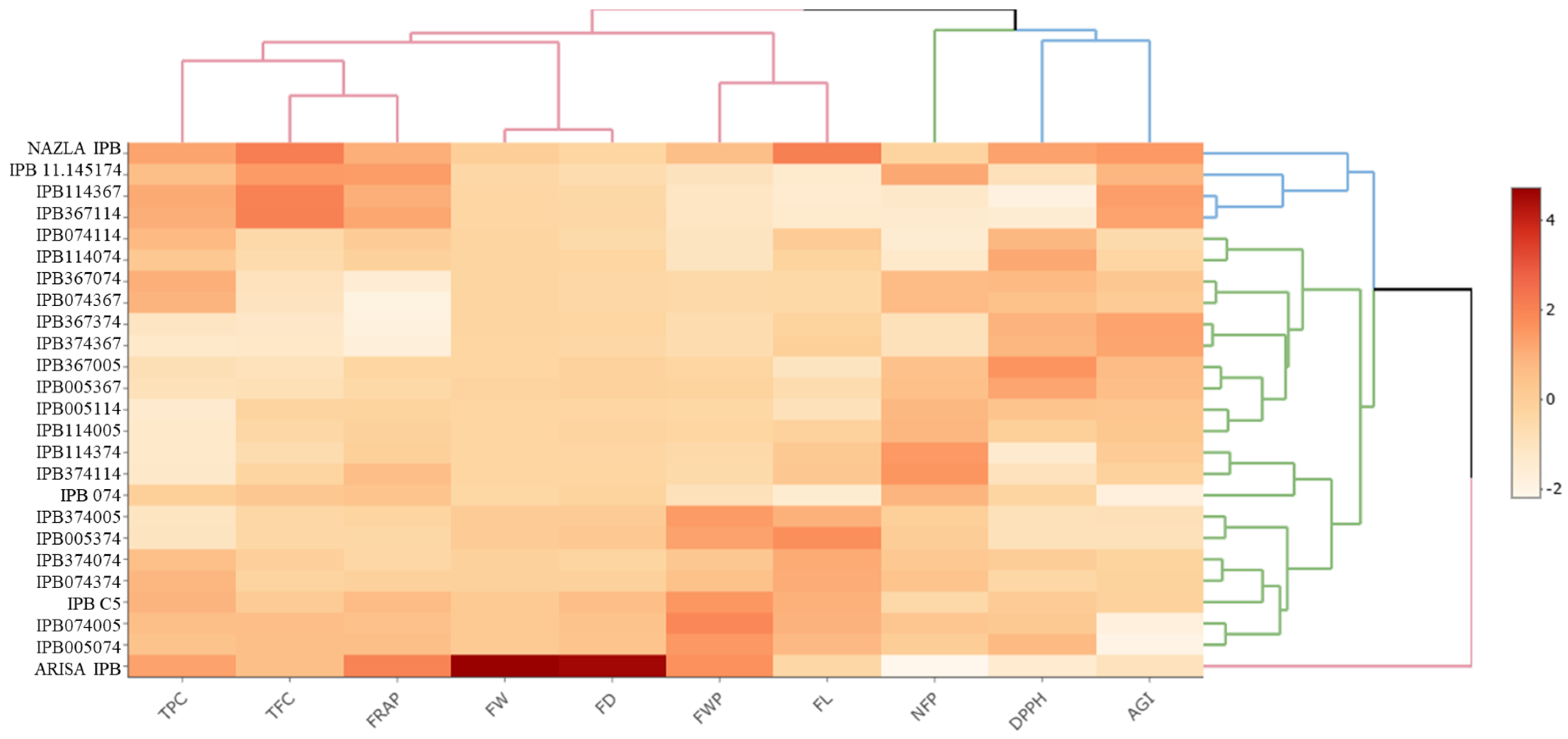
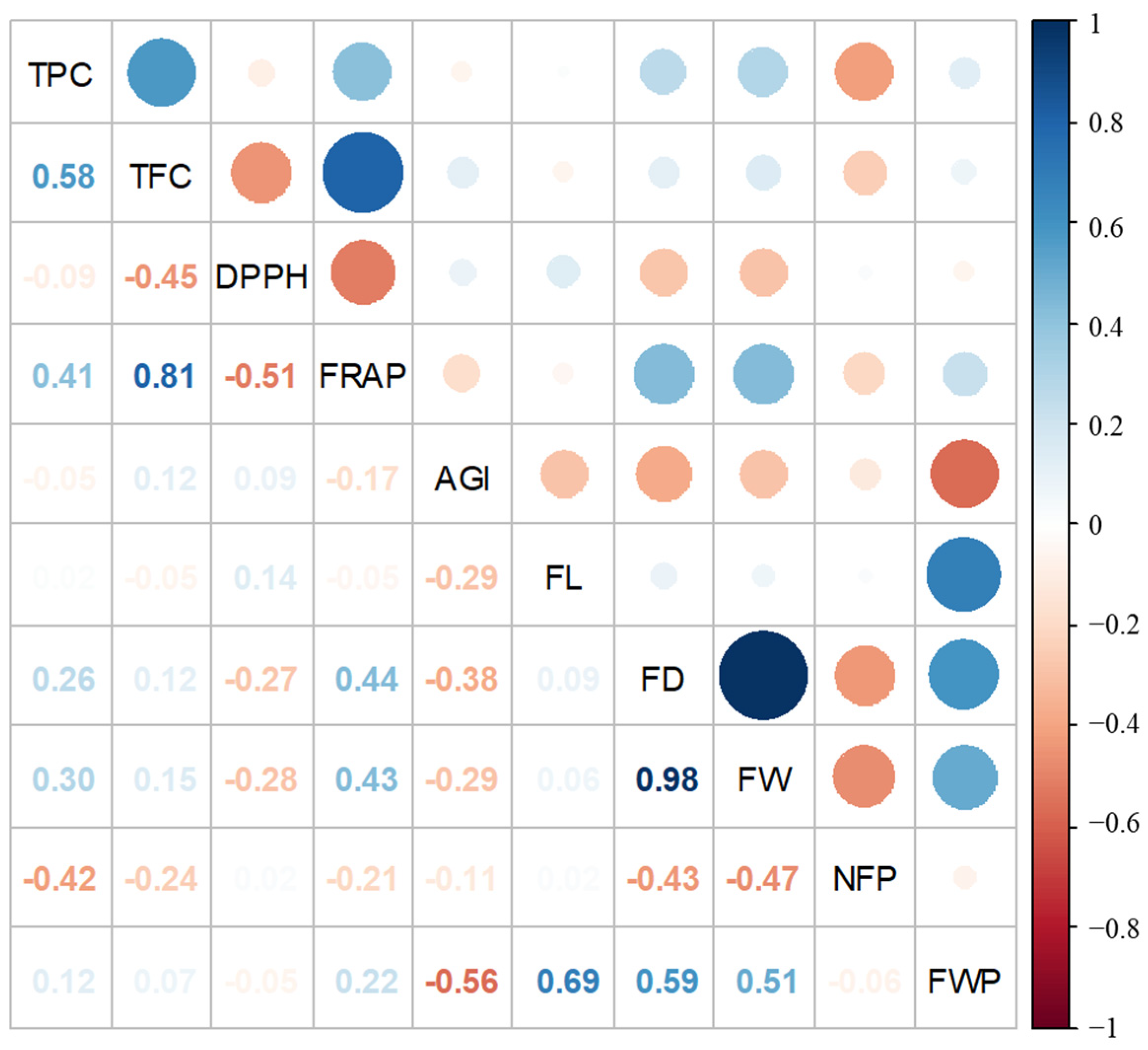
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howe, D.C.; Rahul, S.; Chauhan, A.T.S.; Ronald, M.B. Paradigm shifts caused by the COVID-19 pandemic. Organ. Dyn. 2021, 50, 100804. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, J.; John, M. COVID-19 and global food security. EuroChoices 2020, 19, 26–33. [Google Scholar] [CrossRef]
- Alfauzaan, D.F. Consumer preference for some genotypes of ornamental chili (Capsicum spp.) population F2. IOP Conf. Ser. Earth Environ. Sci. 2021, 694, 012009. [Google Scholar] [CrossRef]
- Barboza, G.E.; Carolina, C.G.; de Bem Bianchetti, L.; María, V.R.; Marisel, S. Monograph of wild and cultivated chili peppers (Capsicum L., Solanaceae). PhytoKeys 2022, 200, 1–423. [Google Scholar] [CrossRef]
- Mujoko, T. Chili Plants: Nutrition Content and Local Varieties as a Genetic Resources. Nusant. Sci. Technol. Proc. 2021, 27, 5–9. [Google Scholar]
- Rachmaniah, M.; Suroso, A.I.; Syukur, M.; Hermadi, I. Supply and demand model for a chili enterprise system using a simultaneous equations system. Economies 2022, 10, 312. [Google Scholar] [CrossRef]
- Surya, R.; Tedjakusuma, F. Diversity of sambals, traditional Indonesian chili pastes. J. Ethn. Food 2022, 9, 25. [Google Scholar] [CrossRef]
- Velázquez, R.; Casquete, R.; Hernández, A.; Martín, A.; Córdoba, M.G.; Coleto, J.M.; Bartolomé, T. Effect of plant density and harvesting type on yield and quality of fresh and dried peppers and paprika. J. Sci. Food Agric. 2019, 99, 400–408. [Google Scholar] [CrossRef]
- Sahid, Z.D.; Syukur, M.; Maharijaya, A.; Nurcholis, W. Total phenolics, flavonoids, antioxidant activity, and α-glucosidase inhibitory activity of ornamental pepper and several other lines. Ornam. Hortic. 2022, 28, 230–238. [Google Scholar] [CrossRef]
- Syukur, M.; Maharijaya, A.; Aisyah, S.I.; Sukma, D.; Ritonga, A.W.; Hakim, A.; Istiqlal, M.R.A.; Sahid, Z.D. Recent progress in the ornamental pepper breeding in Indonesia. In Proceedings of the II International Symposium on Tropical and Subtropical Ornamentals, Bogor, Indonesia, 22–24 July 2020; Volume 1334, pp. 21–28. [Google Scholar]
- Dos Anjos, G.L.; Moreira, G.C.; Carneiro, C.N.; Dias, F.D.S. Effect of phytoregulators on the composition of phenolic compounds in chili peppers (Capsicum frutescens) and exploratory analysis. Sci. Hortic. 2022, 292, 110660. [Google Scholar] [CrossRef]
- Guo, Y.; Bai, J.; Duan, X.; Wang, J. Accumulation characteristics of carotenoids and adaptive fruit color variation in ornamental pepper. Sci. Hortic. 2021, 275, 109699. [Google Scholar] [CrossRef]
- Taiti, C.; Costa, C.; Migliori, C.A.; Comparini, D.; Figorilli, S.; Mancuso, S. Correlation between volatile compounds and spiciness in domesticated and wild fresh chili peppers. Food Bioprocess Technol. 2019, 12, 1366–1380. [Google Scholar] [CrossRef]
- Zamljen, T.; Slatnar, A.; Hudina, M.; Veberic, R.; Medic, A. Characterization and quantification of capsaicinoids and phenolic compounds in two types of chili olive oils, using HPLC/MS. Foods 2022, 11, 2256. [Google Scholar] [CrossRef]
- Sahid, Z.D.; Syukur, M.; Maharijaya, A.; Nurcholis, W. Total phenolic and flavonoid contents, antioxidant, and α-glucosidase inhibitory activities of several big chili (Capsicum annuum L.) genotypes. Cienc. Rural. 2023, 53, e20210913. [Google Scholar] [CrossRef]
- Li, M.; Bao, X.; Zhang, X.; Ren, H.; Cai, S.; Hu, X.; Yi, J. Exploring the phytochemicals and inhibitory effects against α-glucosidase and dipeptidyl peptidase-IV in Chinese pickled chili pepper: Insights into mechanisms by molecular docking analysis. LWT 2022, 162, 113467. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef]
- Devi, J.; Sagar, V.; Kaswan, V.; Ranjan, J.K.; Kumar, R.; Mishra, G.P.; Verma, R.K. Advances in breeding strategies of bell peppers (Capsicum annuum). Adv. Plant Breed. Strateg. Veg. Crops 2021, 9, 3–58. [Google Scholar]
- Naves, E.R.; Scossa, F.; Araújo, W.L.; Nunes-Nesi, A.; Fernie, A.R.; Zsögön, A. Hotter chili peppers by hybridisation: Heterosis and reciprocal effects. bioRxiv 2021, 9, 460669. [Google Scholar]
- Zhao, Y.; Mette, M.F.; Reif, J.C. Genomic selection in hybrid breeding. Plant Breed 2015, 134, 1–10. [Google Scholar] [CrossRef]
- International Plant Genetic Resources Institute. Descriptors for Capsicum (Capsicum spp.); Roma Publisher: Amsterdam, The Netherlands, 1995; Volume 51. [Google Scholar]
- Wang, C.; Rong, H.; Zhang, X.; Shi, W.; Hong, X.; Liu, W.; Yu, Q. Effects and mechanisms of foliar application of silicon and selenium composite sols on diminishing cadmium and lead translocation and affiliated physiological and biochemical responses in hybrid rice (Oryza sativa L.) exposed to cadmium and lead. Chemosphere 2020, 251, 126347. [Google Scholar] [CrossRef]
- Senguttuvel, P.; Sravanraju, N.; Jaldhani, V.; Divya, B.; Beulah, P.; Nagaraju, P.; Subrahmanyam, D. Evaluation of genotype by environment interaction and adaptability in lowland irrigated rice hybrids for grain yield under high temperature. Sci. Rep. 2021, 11, 15825. [Google Scholar] [CrossRef] [PubMed]
- Yidan, O.; Li, X.; Zhang, Q. Understanding the genetic and molecular constitutions of heterosis for developing hybrid rice. J. Genet. Genom. 2022, 49, 385–393. [Google Scholar]
- Barth, E.; Resende, J.T.V.; Zeist, A.R.; Mariguele, K.H.; Zeist, R.A.; Gabriel, A.; Piran, F. Yield and quality of strawberry hybrids under subtropical conditions. Genet. Mol. Res. 2019, 18, GMR18156. [Google Scholar] [CrossRef]
- Saridas, M.A.; Simsek, O.; Donmez, D.; Kacar, Y.A.; Kargi, S.P. Genetic diversity and fruit characteristics of new superior hybrid strawberry (Fragaria × ananassa Duchesne ex Rozier) genotypes. Genet. Resour. Crop Evol. 2021, 68, 741–758. [Google Scholar] [CrossRef]
- Kamiński, P.; Marasek-Ciolakowska, A.; Podwyszyńska, M.; Starzycki, M.; Starzycka-Korbas, E.; Nowak, K. Development and Characteristics of Interspecific Hybrids between Brassica oleracea L. and B. napus L. Agronomy 2020, 10, 1339. [Google Scholar] [CrossRef]
- Akhatar, J.; Kumar, H.; Kaur, H. Recent Progress in Brassica Hybrid Breeding. In Plant Male Sterility Systems for Accelerating Crop Improvement; Springer: Berlin/Heidelberg, Germany, 2022; pp. 195–219. [Google Scholar]
- Arlee, R.; Suanphairoch, S.; Pakdeechanuan, P. Differences in chemical components and antioxidant-related substances in virgin coconut oil from coconut hybrids and their parents. Int. Food Res. J. 2013, 20, 2103. [Google Scholar]
- Mahayu, W.; Kumaunang, J.; Maskromo, I. Identification of superior Dwarf coconut (Cocos nucifera L.) parental cultivars for hybrid breeding. SABRAO J. Breed. Genet. 2021, 53, 278–289. [Google Scholar]
- Sahid, Z.D.; Syukur, M.; Maharijaya, A. Diversity of capsaicin content, quantitative, and yield components in chili (Capsicum annuum) genotypes and their F1 hybrid. Biodiversitas 2020, 21, 2251–2257. [Google Scholar] [CrossRef]
- Naves, E.R.; Silva, L.Á.; Sulpice, R.; Araújo, W.L.; Nunes-Nesi, A.; Peres, L.E.P.; Zsögön, A. Capsaicinoids: Pungency beyond Capsicum. Trends Plant Sci. 2019, 24, 109–120. [Google Scholar] [CrossRef]
- Baenas, N.; Belović, M.; Ilic, N.; Moreno, D.A.; García-Viguera, C. Industrial use of pepper (Capsicum annum L.) derived products: Technological benefits and biological advantages. Food Chem. 2019, 274, 872–885. [Google Scholar] [CrossRef]
- Lu, M.; Chen, C.; Lan, Y.; Xiao, J.; Li, R.; Huang, J.; Ho, C.T. Capsaicin—The major bioactive ingredient of chili peppers: Bio-efficacy and delivery systems. Food Funct. 2020, 11, 2848–2860. [Google Scholar] [CrossRef]
- Azlan, A.; Sultana, S.; Huei, C.S.; Razman, M.R. Antioxidant, anti-obesity, nutritional and other beneficial effects of different chili pepper: A review. Molecules 2022, 27, 898. [Google Scholar] [CrossRef]
- Thongin, S.; Den-Udom, T.; Uppakara, K.; Sriwantana, T.; Sibmooh, N.; Laolob, T.; Ketsawatsomkron, P. Beneficial effects of capsaicin and dihydrocapsaicin on endothelial inflammation, nitric oxide production and antioxidant activity. Biomed. Pharmacother. 2022, 154, 113521. [Google Scholar] [CrossRef]
- Johnson, J.B.; Mani, J.S.; Naiker, M. Correlations between Capsaicin, Dihydrocapsaicin and Phenolic Content in Habanero Chillies. Biol. Life Sci. Forum 2021, 6, 30. [Google Scholar]
- Liu, Y.; Chen, Y.; Wang, Y.; Chen, J.; Huang, Y.; Yan, Y.; Xiao, Y. Total phenolics, capsaicinoids, antioxidant activity, and α-glucosidase inhibitory activity of three varieties of pepper seeds. Int. J. Food Prop. 2020, 23, 1016–1035. [Google Scholar] [CrossRef]
- Gupta, R.; Kapoor, B.; Gulati, M.; Kumar, B.; Gupta, M.; Singh, S.K.; Awasthi, A. Sweet pepper and its principle constituent capsiate: Functional properties and health benefits. Crit. Rev. Food Sci. Nutr. 2022, 62, 7370–7394. [Google Scholar] [CrossRef]
- Dhurhania, C.E.; Novianto, A. Uji Kandungan Total fenolik dan Pengaruhnya terhadap aktivitas antioksidan dari berbagai bentuk sediaan sarang semut (Myrmecodia pendens). J. Farm. Dan Ilmu Kefarmasian Indones. 2018, 5, 62–68. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The flavonoid biosynthesis network in plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2018, 12, 1–12. [Google Scholar] [CrossRef]
- Huang, P.; Jin, L.X.; Lu, J.F.; Gao, Y.H.; Guo, S.B. A theoretical study of radical scavenging antioxidant activity of 3-styrylchromone derivatives using DFT based on quantum chemical descriptors. Theor. Chem. Acc. 2021, 140, 48. [Google Scholar] [CrossRef]
- Nurcholis, W.; Khumaida, N.; Syukur, M.; Bintang, M. Evaluation of free radical scavenging activity of ethanolic extract from promising accessions of Curcuma aeruginosa Roxb. Molekul 2017, 12, 133–138. [Google Scholar] [CrossRef]
- Ramadhan, H.; Baidah, D.; Lestrai, N.P.; Yuliana, K.A. Antioxidant activity of 96% ethanol extract of terap (Artocarpus odorratissimus) leaf, flesh and peel using cuprac method. Farmasainsm 2020, 7, 7–12. [Google Scholar] [CrossRef]
- Rodrigues-Salvador, A.; Lana-Costa, J.; Omena-Garcia, R.P.; Batista-Silva, W.; Scossa, F.; Rosado-Souza, L.; Pérez-Díaz, J.L.; Menezes-Silva, P.E.; DaMatta, F.M.; Sulpice, R.; et al. Metabolic shifts during fruit development in pungent and non-pungent peppers. Food Chem. 2021, 375, 131850. [Google Scholar] [CrossRef] [PubMed]
- Nurcholis, W.; Putri, D.S.; Husnawati, H.; Aisyah, S.I.; Priosoeryanto, B.P. Total flavonoid content and antioxidant activity of ethanol and ethyl acetate extracts from accessions of Amomum compactum fruits. Ann. Agric. Sci. 2021, 66, 58–62. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Y.; Zhou, Z. Phenolic and flavonoid contents of mandarin (Citrus reticulata Blanco) fruit tissues and their antioxidant capacity as evaluated by DPPH and ABTS methods. J. Integr. Agric. 2018, 17, 256–263. [Google Scholar] [CrossRef]
- Watcharachaisoponsiri, T.; Sornchan, P.; Charoenkiatkul, S.; Suttisansanee, U. The [alpha]-glucosidase and [alpha]-amylase inhibitory activity from different chili pepper extracts. Int. Food Res. J. 2016, 23, 1439. [Google Scholar]
- Idrees, S.; Hanif, M.A.; Ayub, M.A.; Hanif, A.; Ansari, T.M. Chili pepper. In Medicinal Plants of South Asia; Elsevier: Amsterdam, The Netherlands, 2020; pp. 113–124. [Google Scholar]
- Hossain, U.; Das, A.K.; Ghosh, S.; Sil, P.C. An overview on the role of bioactive α-glucosidase inhibitors in ameliorating diabetic complications. Food Chem. Toxicol. 2020, 145, 111738. [Google Scholar] [CrossRef]
- Xie, X.; Chen, C.; Fu, X. Screening α-glucosidase inhibitors from four edible brown seaweed extracts by ultra-filtration and molecular docking. LWT 2021, 138, 110654. [Google Scholar] [CrossRef]
- Barchenger, D.W.; Clark, R.A.; Gniffke, P.A.; Ledesma, D.R.; Lin, S.W.; Hanson, P.; Kumar, S. Stability of yield and yield components of pepper (Capsicum annuum), and evaluation of publicly available predictive meteorological data in East and Southeast Asia. HortScience 2018, 53, 1776–1783. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| ♀ | ♂ | Code |
|---|---|---|
| IPB C5 | ARISA IPB | IPB005374 |
| IPB C5 | NAZLA IPB | IPB005367 |
| IPB C5 | IPB 11.145174 | IPB005114 |
| IPB C5 | IPB 074 | IPB005074 |
| ARISA IPB | IPB C5 | IPB374005 |
| ARISA IPB | NAZLA IPB | IPB374367 |
| ARISA IPB | IPB 11.145174 | IPB374114 |
| ARISA IPB | IPB 074 | IPB374074 |
| NAZLA IPB | IPB C5 | IPB367005 |
| NAZLA IPB | ARISA IPB | IPB367374 |
| NAZLA IPB | IPB 11.145174 | IPB367114 |
| NAZLA IPB | IPB 074 | IPB367074 |
| IPB 11.145174 | IPB C5 | IPB114005 |
| IPB 11.145174 | ARISAIPB | IPB114374 |
| IPB 11.145174 | NAZLA IPB | IPB114367 |
| IPB 11.145174 | IPB 074 | IPB114074 |
| IPB 074 | IPB C5 | IPB074005 |
| IPB 074 | ARISA IPB | IPB074374 |
| IPB 074 | NAZLA IPB | IPB074367 |
| IPB 074 | IPB 11.145174 | IPB074114 |
| Sources | Mean Square | ||||
|---|---|---|---|---|---|
| FL | FW | FD | NFP | FWP | |
| Genotype | 27.86 ** | 1498.52 ** | 540.15 ** | 1284.50 ** | 268,976.65 ** |
| Replication | 0.257 * | 1.45 ns | 0.228 ns | 1.61 ns | 1088.35 ns |
| Error | 0.102 | 1.423 | 0.226 | 11.072 | 946.33 |
| Coefficient of Variance (%) | 4.12 | 11.64 | 3.36 | 4.92 | 6.89 |
| Sources | Mean Square | ||||
|---|---|---|---|---|---|
| TPC | TFC | DPPH | FRAP | AGI | |
| Genotype | 213.84 ** | 1.97 ** | 0.08 ** | 235.80 ** | 302.05 ** |
| Replication | 4.90 ** | 0.01 ns | 0.002 ns | 3.07 * | 38.40 ** |
| Error | 0.79 | 0.006 | 0.004 | 1.26 | 1.13 |
| Coefficient of Variance (%) | 3.16 | 3.20 | 1.19 | 3.26 | 1.60 |
| Genotype | FL (cm) | FD (mm) | FW (g) | NFP | FWP (g) |
|---|---|---|---|---|---|
| IPB005374 | 12.94 b | 17.09 d | 12.93 bc | 68.00 hi | 833.50 d |
| IPB005367 | 5.83 h | 11.98 fg | 4.84 f | 79.00 def | 382.50 g |
| IPB005114 | 5.07 i | 9.53 jk | 2.81 fg | 84.33 cd | 337.05 ghi |
| IPB005074 | 10.10 d | 19.26 c | 13.24 bc | 68.00 hi | 900.20 bc |
| IPB374005 | 10.70 c | 15.87 e | 11.42 cd | 67.00 hi | 879.13 cd |
| IPB374367 | 7.53 f | 8.56 l | 3.17 fg | 49.67 k | 251.23 j |
| IPB374114 | 8.57 e | 9.72 j | 2.95 fg | 99.67 a | 293.71 ij |
| IPB374074 | 11.10 c | 10.34 ij | 7.23 e | 72.67 gh | 524.94 f |
| IPB367005 | 4.53 j | 11.68 gh | 3.23 fg | 77.33 efg | 349.49 ghi |
| IPB367374 | 7.07 f | 9.90 j | 3.91 fg | 49.67 k | 251.85 j |
| IPB367114 | 3.57 k | 8.73 kl | 2.64 fg | 39.00 l | 110.19 l |
| IPB367074 | 6.33 gh | 8.74 kl | 3.81 fg | 82.00 cde | 312.29 hi |
| IPB114005 | 7.30 f | 10.98 hi | 2.88 fg | 84.67 cd | 351.80 gh |
| IPB114374 | 8.33 e | 9.49 jk | 3.16 fg | 98.67 a | 297.04 hij |
| IPB114367 | 3.37 k | 8.18 lm | 2.54 fg | 41.67 l | 106.07 l |
| IPB114074 | 7.07 f | 9.98 j | 3.53 fg | 40.67 l | 133.96 kl |
| IPB074005 | 10.67 c | 20.01 c | 13.62 b | 74.00 fg | 1008.85 a |
| IPB074374 | 11.03 c | 12.70 f | 7.62 e | 75.67 fg | 593.58 e |
| IPB074367 | 6.33 gh | 8.57 l | 3.60 fg | 82.33 cde | 296.90 hij |
| IPB074114 | 8.10 e | 7.41 m | 3.72 fg | 36.33 l | 135.18 kl |
| ARISA IPB | 6.47 g | 25.3 2 a | 15.68 a | 22.33 m | 946.11 b |
| IPB 11.145174 | 3.63 k | 5.95 n | 1.63 g | 92.00 b | 156.76 kl |
| IPB C5 | 10.70 c | 22.81 b | 14.00 b | 58.33 j | 906.86 bc |
| NAZLA IPB | 3.30 k | 9.89 j | 2.13 g | 86.00 c | 182.92 k |
| IPB 074 | 14.07 a | 11.12 hi | 9.93 d | 62.67 ij | 619.40 e |
| Parent’s Mean | 7.63 | 25.02 | 28.68 | 64.27 | 562.41 |
| Hybrid’s Mean | 7.78 | 11.44 | 5.64 | 68.52 | 417.47 |
| Hybrid vs. Parent | ** | ** | ** | ** | ns |
| Genotype | TPC (mg GAE g−1 DW)) | TFC (mg QE g−1 DW) | DPPH (µmol TE g−1 DW) | FRAP (µmol TE g−1 DW) | AGI (%) |
|---|---|---|---|---|---|
| IPB005374 | 19.30 k | 2.01 gh | 1.60 i | 30.70 ij | 57.47 k |
| IPB005367 | 20.82 j | 1.70 j | 1.95 b | 30.39 j | 72.46 d |
| IPB005114 | 16.37 l | 2.14 g | 1.81 de | 32.64 hi | 69.65 e |
| IPB005074 | 31.59 g | 2.81 c | 1.87 c | 39.61 def | 46.54 l |
| IPB374005 | 18.96 k | 2.03 gh | 1.60 i | 31.91 hij | 57.91 k |
| IPB374367 | 17.07 l | 1.36 l | 1.89 c | 19.36 kl | 78.75 b |
| IPB374114 | 17.38 l | 2.12 g | 1.59 i | 40.21 de | 64.83 h |
| IPB374074 | 32.69 g | 2.29 f | 1.75 fg | 31.00 ij | 63.97 hi |
| IPB367005 | 21.38 j | 1.59 jk | 2.01 a | 31.73 hij | 73.45 cd |
| IPB367374 | 19.07 k | 1.37 l | 1.90 c | 18.33 l | 79.07 b |
| IPB367114 | 36.81 cd | 4.00 a | 1.50 j | 45.18 bc | 79.03 b |
| IPB367074 | 36.71 cd | 1.57 k | 1.87 c | 20.76 k | 68.83 ef |
| IPB114005 | 17.21 l | 2.03 gh | 1.74 g | 33.55 gh | 69.18 e |
| IPB114374 | 17.07 l | 1.83 i | 1.52 j | 33.73 gh | 66.89 g |
| IPB114367 | 37.51 bc | 3.97 a | 1.44 k | 43.42 c | 80.25 ab |
| IPB114074 | 29.87 h | 1.90 hi | 1.94 b | 33.36 gh | 62.89 i |
| IPB074005 | 32.96 fg | 2.86 c | 1.78 ef | 38.64 ef | 48.30 l |
| IPB074374 | 34.94 e | 2.14 g | 1.68 h | 33.24 h | 64.37 hi |
| IPB074367 | 35.66 de | 1.52 k | 1.83 d | 17.97 l | 67.32 fg |
| IPB074114 | 34.39 ef | 1.96 h | 1.88 c | 35.30 g | 61.07 j |
| ARISA IPB | 39.07 a | 2.83 c | 1.51 j | 52.15 a | 56.66 k |
| IPB 11.145174 | 33.00 fg | 3.52 b | 1.60 i | 46.88 b | 74.62 c |
| IPB C5 | 35.62 de | 2.43 e | 1.76 fg | 40.70 d | 64.63 hi |
| NAZLA IPB | 38.69 ab | 4.06 a | 1.95 b | 43.55 c | 81.22 a |
| IPB 074 | 27.38 i | 2.56 d | 1.70 h | 38.09 f | 48.28 l |
| Parent’s mean | 34.75 | 3.08 | 1.71 | 44.27 | 65.08 |
| Hybrid’s mean | 26.39 | 2.16 | 1.76 | 32.05 | 66.61 |
| Hybrid vs. Parent | ** | ** | ** | ** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syukur, M.; Maharijaya, A.; Nurcholis, W.; Ritonga, A.W.; Istiqlal, M.R.A.; Hakim, A.; Sulassih, S.; Perdani, A.Y.; Pangestu, A.Y.; Hatta, A.N.N.L.; et al. Biochemical and Yield Component of Hybrid Chili (Capsicum annuum L.) Resulting from Full Diallel Crosses. Horticulturae 2023, 9, 620. https://doi.org/10.3390/horticulturae9060620
Syukur M, Maharijaya A, Nurcholis W, Ritonga AW, Istiqlal MRA, Hakim A, Sulassih S, Perdani AY, Pangestu AY, Hatta ANNL, et al. Biochemical and Yield Component of Hybrid Chili (Capsicum annuum L.) Resulting from Full Diallel Crosses. Horticulturae. 2023; 9(6):620. https://doi.org/10.3390/horticulturae9060620
Chicago/Turabian StyleSyukur, Muhamad, Awang Maharijaya, Waras Nurcholis, Arya Widura Ritonga, Muhammad Ridha Alfarabi Istiqlal, Abdul Hakim, Sulassih Sulassih, Ambar Yuswi Perdani, Arya Yuda Pangestu, Andi Nadia Nurul Lathifa Hatta, and et al. 2023. "Biochemical and Yield Component of Hybrid Chili (Capsicum annuum L.) Resulting from Full Diallel Crosses" Horticulturae 9, no. 6: 620. https://doi.org/10.3390/horticulturae9060620