

In Vitro Shoot Multiplication and Regeneration of the Recalcitrant Rocket (Eruca sativa Mill.) Variety Domaća Rukola
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Conditions
2.2. Shoot Culture Establishment, Multiplication, and Rooting
2.3. Extract Preparation and HPLC Analysis of Flavonoids in Multiplied Shoots
2.4. De Novo Shoot Regeneration from Different Explants of Rocket Seedlings
2.5. Somatic Embryogenesis from Immature Zygotic Embryos
2.6. Histological Analysis
2.7. Statistical Analysis
3. Results
3.1. Shoot Multiplication from Epicotyl Explants of Seedlings
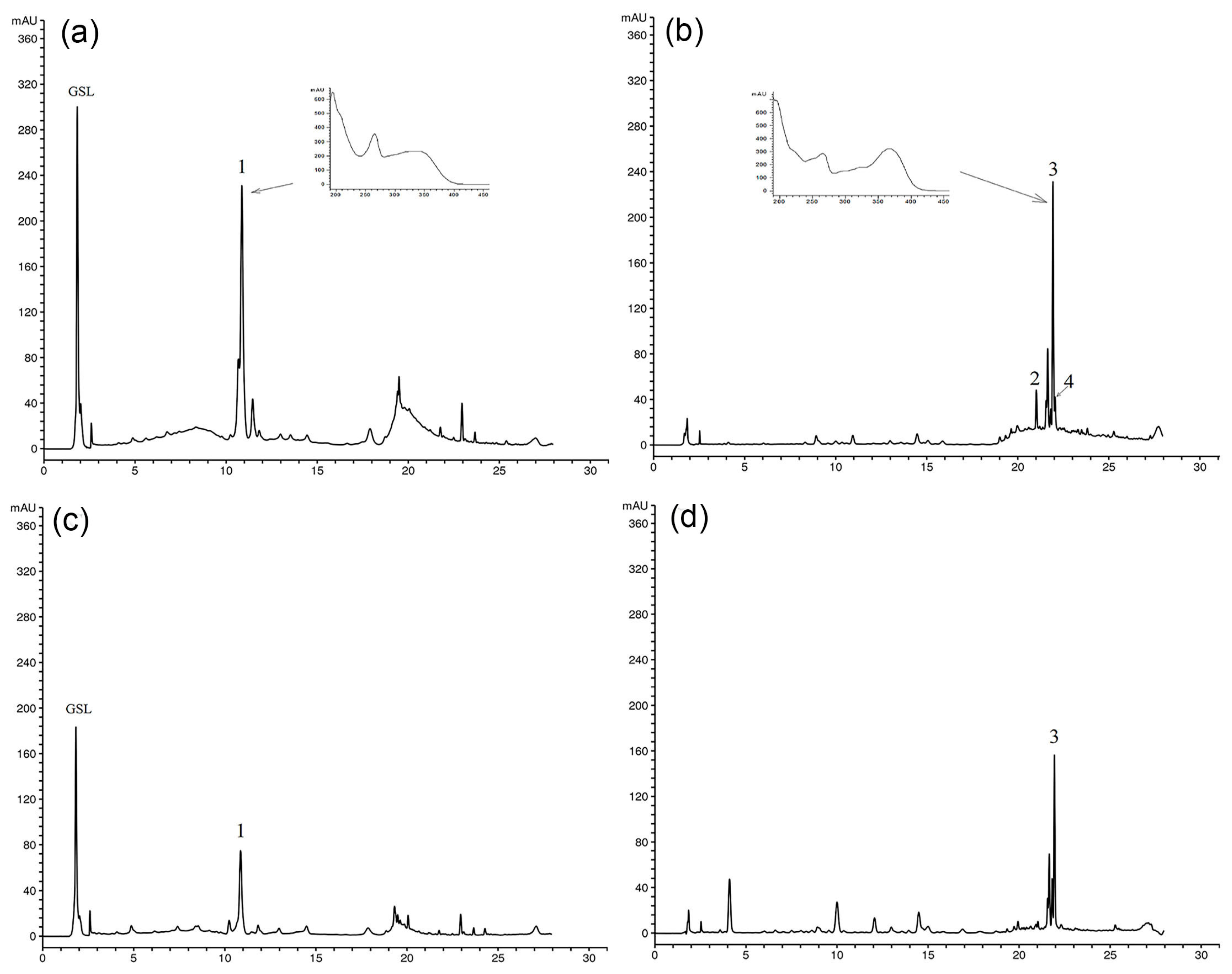
3.2. The Content of Secondary Metabolites in Multiplied Shoots Grown In Vitro
3.3. Rooting of Multiplied Shoots
3.4. De Novo Shoot Organogenesis
3.5. Somatic Embryogenesis from Immature Zygotic Embryos
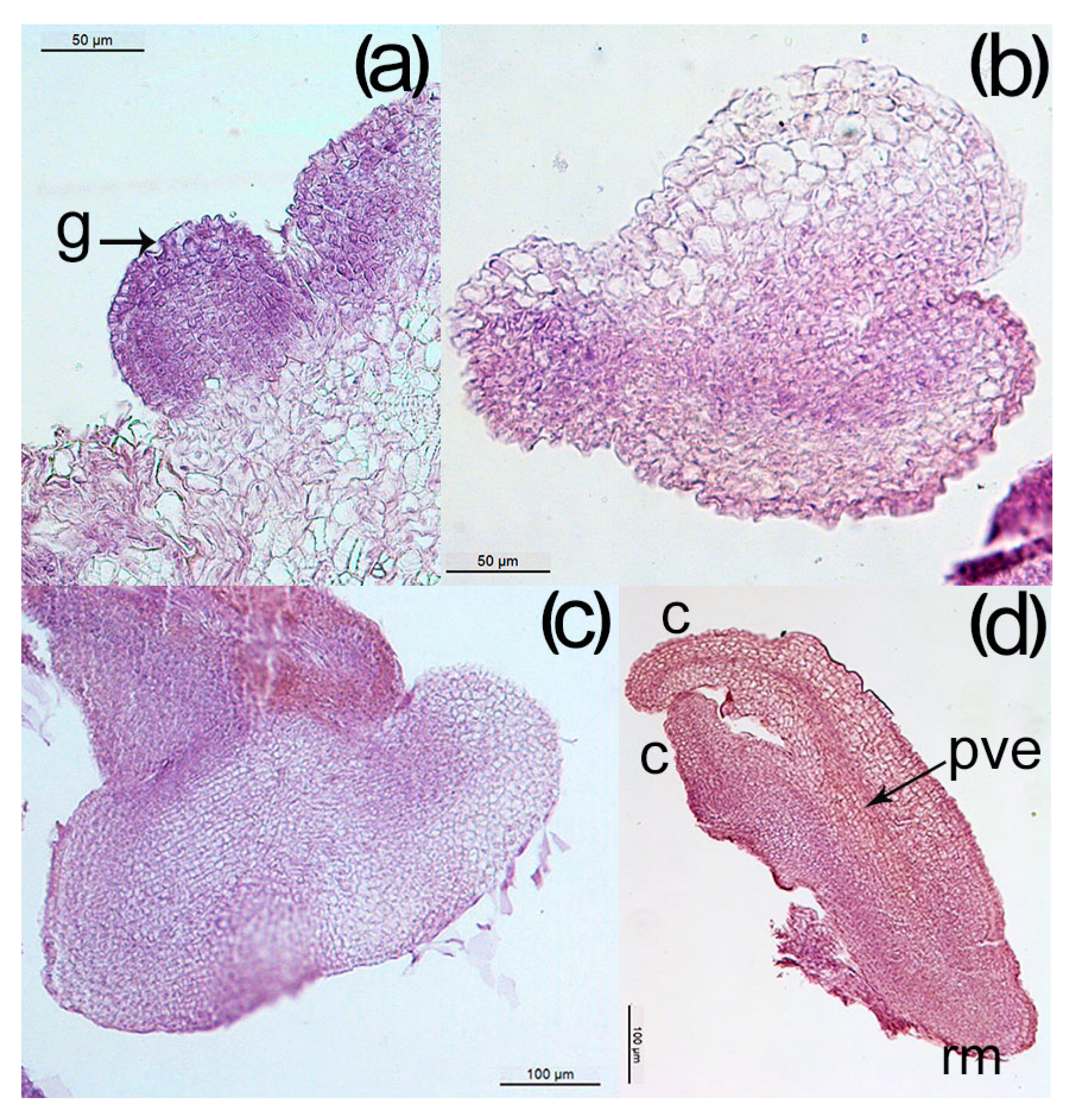
3.6. Histological Analysis of Somatic Embryogenesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, R.N.; Carvalho, R.; Mellon, F.A.; Eagles, J.; Rosa, E.A.S. Identification and Quantification of Glucosinolates in Sprouts Derived from Seeds of Wild Eruca sativa L. (Salad Rocket) and Diplotaxis tenuifolia L. (Wild Rocket) from Diverse Geographical Locations. J. Agric. Food Chem. 2007, 55, 67–74. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R. Protein Nutritional Value of Rocket Leaves and Possibilities of Its Modification during Plant Growth. Turk. J. Agric. For. 2015, 39, 1023–1028. [Google Scholar] [CrossRef]
- Morshedy, S.A.; Abdelmodather, A.M.; Basyony, M.M.; Zahran, S.A.; Hassan, M.A. Effects of Rocket Seed Oil, Wheat Germ Oil, and Their Mixture on Growth Performance, Feed Utilization, Digestibility, Redox Status, and Meat Fatty Acid Profile of Growing Rabbits. Agriculture 2021, 11, 662. [Google Scholar] [CrossRef]
- Yaniv, Z.; Schafferman, D.; Amar, Z. Tradition, Uses and Biodiversity of Rocket (Eruca sativa, Brassicaceae) in Israel. Econ. Bot. 1998, 52, 394–400. [Google Scholar] [CrossRef]
- Kim, S.J.; Ishii, G. Glucosinolate Profiles in the Seeds, Leaves and Roots of Rocket Salad (Eruca sativa Mill.) and Anti-Oxidative Activities of Intact Plant Powder and Purified 4-Methoxyglucobrassicin. Soil Sci. Plant Nutr. 2006, 52, 394–400. [Google Scholar] [CrossRef]
- Sarwar Alam, M.; Kaur, G.; Jabbar, Z.; Javed, K.; Athar, M. Eruca sativa Seeds Possess Antioxidant Activity and Exert a Protective Effect on Mercuric Chloride Induced Renal Toxicity. Food Chem. Toxicol. 2007, 45, 910–920. [Google Scholar] [CrossRef]
- Alqasoumi, S. Carbon Tetrachloride-Induced Hepatotoxicity: Protective Effect of “Rocket” Eruca sativa L. in Rats. Am. J. Chin. Med. 2010, 38, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, G.; Bottino, A.; Di Stasio, E.; Vallone, S.; Maggio, A. Proline and Light as Quality Enhancers of Rocket (Eruca sativa Miller) Grown under Saline Conditions. Sci. Hortic. 2011, 128, 393–400. [Google Scholar] [CrossRef]
- Barillari, J.; Canistro, D.; Paolini, M.; Ferroni, F.; Pedulli, G.F.; Iori, R.; Valgimigli, L. Direct Antioxidant Activity of Purified Glucoerucin, the Dietary Secondary Metabolite Contained in Rocket (Eruca sativa Mill.) Seeds and Sprouts. J. Agric. Food Chem. 2005, 53, 2475–2482. [Google Scholar] [CrossRef] [PubMed]
- Sacan, O.; Orak, H.; Yanardag, R. Antioxidant Activity of Water Extract of Eruca sativa Mill. Asian J. Chem. 2008, 20, 3462–3474. [Google Scholar]
- Iori, R.; Bernardi, R.; Gueyrard, D.; Rollin, P.; Palmieri, S. Formation of Glucoraphanin by Chemoselective Oxidation of Natural Glucoerucin: A Chemoenzymatic Route to Sulforaphane. Bioorg. Med. Chem. Lett. 1999, 9, 1047–1048. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, X.B.; Jiang, J.L.; Wang, X.Y. Plantlet Regeneration from Cotyledon, Cotyledon Petiole, and Hypocotyl Explants via Somatic Embryogenesis Pathway in Roquette (Eruca sativa Mill). Plant Biosyst. 2011, 145, 68–76. [Google Scholar] [CrossRef]
- Bonnesen, C.; Eggleston, I.M.; Hayes, J.D. Dietary Indoles and Isothiocyanates That Are Generated from Cruciferous Vegetables Can Both Stimulate Apoptosis and Confer Protection against DNA Damage in Human Colon Cell Lines. Cancer Res. 2001, 61, 6120–6130. [Google Scholar]
- Qaddoumi, S.; El-Banna, N. Antimicrobial Activity of Arugula (Eruca sativa) Leaves on Some Pathogenic Bacteria. Int. J. Biol. 2019, 11, 10–15. [Google Scholar] [CrossRef]
- Jin, J.; Koroleva, O.A.; Gibson, T.; Swanston, J.; Maganj, J.; Zhang, Y.A.N.; Rowland, I.R.; Wagstaff, C. Analysis of Phytochemical Composition and Chemoprotective Capacity of Rocket (Eruca sativa and Diplotaxis tenuifolia) Leafy Salad Following Cultivation in Different Environments. J. Agric. Food Chem. 2009, 57, 5227–5234. [Google Scholar] [CrossRef]
- Zhang, T.; Cao, Z.Y.; Wang, X.Y. Induction of Somatic Embryogenesis and Plant Regeneration from Cotyledon and Hypocotyl Explants of Eruca sativa Mill. Vitr. Cell. Dev. Biol. Plant 2005, 41, 655–657. [Google Scholar] [CrossRef]
- Panić, M.; Stojšin, V.; Budakov, D.; Grahovac, M. Pseudomycosis and Mycosis of Rocket Plant (Arugula). Biljn. Lek. 2021, 49, 38–53. [Google Scholar] [CrossRef]
- Srivastava, J.N.; Singh, A.K. Pulses, Oil Seeds, Narcotics, and Sugar Crops. In Diseases of Field Crops Diagnosis and Management, 1st ed.; Diseases of Taramira (Eruca sativa Mill.) and Their Management: Indian Perspective; Srivastava, J.N., Singh, A.K., Eds.; Apple Academic Press: Cambridge, MA, USA, 2020; Volume 2, pp. 307–319. [Google Scholar] [CrossRef]
- Kular, J.; Kumar, S. Quantification of Avoidable Yield Losses in Oilseed Brassica Caused by Insect Pests. J. Plant Prot. Res. 2011, 51, 38–43. [Google Scholar] [CrossRef]
- Soroka, J.; Grenkow, L. Susceptibility of Brassicaceous Plants to Feeding by Flea Beetles, Phyllotreta Spp. (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2013, 106, 2557–2567. [Google Scholar] [CrossRef]
- Ashloowalia, B.S. Somatic Embryogenesis and Plant Regeneration in Eruca sativa. Crop Sci. 1987, 27, 813–814. [Google Scholar] [CrossRef]
- Parkash, S.; Chowdhury, J.B. Jain RK Callus Initiation and Regeneration Potential in Different Genotypes of Eruca sativa. Curr. Sci. 1989, 58, 979–980. [Google Scholar]
- Slater, S.M.H.; Keller, W.A.; Scoles, G. Agrobacterium-Mediated Transformation of Eruca sativa. Plant Cell. Tissue Organ Cult. 2011, 106, 253–260. [Google Scholar] [CrossRef]
- Sharma, M.M.; Dhingra, M.; Dave, A.; Batra, A. Plant Regeneration and Stimulation of In Vitro Flowering in Eruca sativa Mill. Afr. J. Biotechnol. 2012, 11, 7906–7911. [Google Scholar] [CrossRef]
- Mokhtar, M.; Nagar, E.; El Nagar, M.M.; Mekawi, E.M. Comparison of Different Genotypes of Rocket (Eruca sativa) in Terms of Chemical Compounds Extracted from Seeds and in the Callus Induced from Tissue Culture. Middle East J. Agric. Res. 2014, 3, 1074–1082. [Google Scholar]
- Abbasi, B.H.; Ali, J.; Ali, M.; Zia, M.; Bokhari, S.A.; Khan, M.A. Free Radical Scavenging Activity in In Vitro-Derived Tissues of Eruca sativa. Toxicol. Ind. Health 2016, 32, 98–105. [Google Scholar] [CrossRef]
- Sikdar, S.R.; Chatterjee, G.; Das, S.; Sen, S.K. Regeneration of Plants from Mesophyll Protoplasts of the Wild Crucifer Eruca sativa Lam. Plant Cell Rep. 1987, 6, 486–489. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Klfriede Linsmaier, B.M.; Skoog, O. Organic Growth Factor Requirements of Tobacco Tissue Cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Llorach, R.; Gil, M.I.; Ferreres, F. Identification of New Flavonoid Glycosides and Flavonoid Profiles to Characterize Rocket Leafy Salads (Eruca vesicaria and Diplotaxis tenuifolia). J. Agric. Food Chem. 2007, 55, 1356–1363. [Google Scholar] [CrossRef]
- Teixeira, A.F.; de Souza, J.; Dophine, D.D.; de Souza Filho, J.D.; Saúde-Guimarães, D.A. Chemical Analysis of Eruca sativa Ethanolic Extract and Its Effects on Hyperuricaemia. Molecules 2022, 27, 1506. [Google Scholar] [CrossRef] [PubMed]
- Panić, M.; Bagi, F. Viruses of Arugula. Biljn. Lek. 2020, 48, 503–509. [Google Scholar] [CrossRef]
- Batra, A.; Dhingra, M. Production of Plantlets of Eruca sativa In Vitro. J. Phytol. Res. 1991, 4, 73–77. [Google Scholar]
- Wawrosch, C.; Zotchev, S.B. Production of Bioactive Plant Secondary Metabolites through In Vitro Technologies—Status and Outlook. Appl. Microbiol. Biotechnol. 2021, 105, 6649–6668. [Google Scholar] [CrossRef] [PubMed]
- Grzegorczyk-Karolak, I.; Kuźma, Ł.; Wysokińska, H. The Effect of Cytokinins on Shoot Proliferation, Secondary Metabolite Production and Antioxidant Potential in Shoot Cultures of Scutellaria alpina. Plant Cell. Tissue Organ Cult. 2015, 122, 699–708. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alamri, S.A.; Alam, P.; Ashraf, M.; Ahmad, P. Potential of Exogenously Sourced Kinetin in Protecting Solanum lycopersicum from NaCl-Induced Oxidative Stress through Up-Regulation of the Antioxidant System, Ascorbate-Glutathione Cycle and Glyoxalase System. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.A.; Khanam, N.; Uddin, M.; Mishra, R.K.; Khan, R. Nanotized Kinetin Enhances Essential Oil Yield and Active Constituents of Mint via Improvement in Physiological Attributes. Chemosphere 2022, 288, 132447–132458. [Google Scholar] [CrossRef]
- Leskovšek, L.; Jakše, M.; Bohanec, B. Doubled Haploid Production in Rocket (Eruca sativa Mill.) through Isolated Microspore Culture. Plant Cell. Tissue Organ Cult. 2008, 93, 181–189. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jinu, U.; Gomathi, M.; Mahendran, D.; Ahmad, N.; Geetha, N.; Sahi, S.V. Role of Silver Nitrate in Plant Regeneration from Cotyledonary Nodal Segment Explants of Prosopis cineraria (L.) Druce.: A Recalcitrant Medicinal Leguminous Tree. Biocatal. Agric. Biotechnol. 2017, 12, 286–291. [Google Scholar] [CrossRef]
- Bhadane, B.S.; Maheshwari, V.L.; Patil, R.H. Quercetin and Silver Nitrate Modulate Organogenesis in Carissa carandas (L.). Vitr. Cell. Dev. Biol. Plant 2018, 54, 600–605. [Google Scholar] [CrossRef]
- Mookkan, M.; Andy, G. AgNO3 Boosted High-Frequency Shoot Regeneration in Vigna mungo (L.) Hepper. Plant Signal. Behav. 2014, 9, e972284-1. [Google Scholar] [CrossRef]
- Sowmya, M.; Jinu, U.; Sarathikannan, D.; Geetha, N.; Girija, S.; Venkatachalam, P. Effect of Silver Nitrate and Growth Regulators on Direct Shoot Organogenesis and In Vitro Flowering from Internodal Segment Explants of Alternanthera sessilis L. Biocatal. Agric. Biotechnol. 2020, 30, 101855. [Google Scholar] [CrossRef]
- Adly, W.M.R.M.; Mazrou, Y.S.A.; El-Denary, M.E.; Mohamed, M.A.; Abd El-Salam, E.S.T.; Fouad, A.S. Boosting Polyamines to Enhance Shoot Regeneration in Potato (Solanum tuberosum L.) Using AgNO3. Horticulturae 2022, 8, 113. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Zhao, Y.; Zong, P.; Zhan, Z.; Piao, Z. Establishment of a Simple and Efficient Agrobacterium-Mediated Genetic Transformation System to Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Hortic. Plant J. 2021, 7, 117–128. [Google Scholar] [CrossRef]
- Giridhar, P.; Obul, R.B.; Ravishankar, G.A. Silver Nitrate Influences in vitro Shoot Multiplication and Root Formation in Vanilla planifolia Andr. Curr. Sci. 2001, 81, 1166–1170. [Google Scholar]
- Gao, H.; Xu, P.; Li, J.; Ji, H.; An, L.; Xia, X. AgNO3 Prevents the Occurrence of Hyperhydricity in Dianthus chinensis L. by Enhancing Water Loss and Antioxidant Capacity. In Vitr. Cell. Dev. Biol. Plant 2017, 53, 561–570. [Google Scholar] [CrossRef]
- Elhiti, M.; Tahir, M.; Gulden, R.H.; Khamiss, K.; Stasolla, C. Modulation of Embryo-Forming Capacity in Culture through the Expression of Brassica Genes Involved in the Regulation of the Shoot Apical Meristem. J. Exp. Bot. 2010, 61, 4069–4085. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, S.; Vinterhalter, B.; Zdravković-Korać, S.; Vinterhalter, D.; Zdravković, J.; Cvikić, D.; Mitić, N. Recurrent Somatic Embryogenesis and Plant Regeneration from Immature Zygotic Embryos of Cabbage (Brassica oleracea var. capitata) and Cauliflower (Brassica oleracea var. botrytis). Plant Cell. Tissue Organ Cult. 2013, 113, 397–406. [Google Scholar] [CrossRef]
- Kalam Azad, M.A.; Rabbani, M.G.; Amin, L. Plant Regeneration and Somatic Embryogenesis from Immature Embryos Derived through Interspecific Hybridization among Different Carica Species. Int. J. Mol. Sci. 2012, 13, 17065–17076. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yuan, Y.; Jiang, H.; Bao, Y.; Ning, G.; Zhao, L.; Zhou, X.; Zhou, H.; Gao, J.; Ma, N. Agrobacterium tumefaciens-Mediated Transformation of Modern Rose (Rosa hybrida) Using Leaf-Derived Embryogenic Callus. Hortic. Plant J. 2021, 7, 359–366. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokinin (mg L−1) | Explant No. | Multiplication Index 1 | Viable Shoots (%) | Hyperhydrated Shoots (%) | Necrosis (%) |
|---|---|---|---|---|---|
| BA 0.1 | 279 | 3.45 ± 0.11 b | 91.28 | 8.72 | 0.00 |
| BA 0.2 | 100 | 4.10 ± 0.16 c | 58.54 | 26.58 | 14.88 |
| KIN 0.1 | 152 | 2.98 ± 0.13 a | 90.73 | 9.27 | 0.00 |
| KIN 0.2 | 123 | 3.15 ± 0.14 ab | 75.45 | 24.55 | 0.00 |
| Extract Sample | Multiplication Treatment | Kaempferol Content ± SE | |||
|---|---|---|---|---|---|
| Cytokinin | Concentration (mg L−1) | (mg g−1 dw) | |||
| In vitro-grown shoots | KIN | 0.1 | 0.68 ± 0.007 *a | ||
| 0.2 | 1.74 ± 0.010 *d | ||||
| BA | 0.1 | 1.50 ± 0.010 *c | |||
| 0.2 | 1.10 ± 0.009 *b | ||||
| Control | - | - | 2.47 ± 0.011 | ||
| ANOVA Source of Variation | Df | Mean Square | F-Ratio | p-Value | |
| Cytokinin | 1 | 0.02484 | 103.730 | 0.000000 | |
| Concentration | 1 | 0.31493 | 1314.94 | 0.000007 | |
| Cytokinin × concentration | 1 | 1.59432 | 6656.88 | 0.000000 | |
| IBA (mg L−1) | Explants No. | Rooting (%) | Mean No. of Roots | Mean Length of the Longest Root (mm) |
|---|---|---|---|---|
| 0 | 34 | 51.84 ± 12.61 a | 5.12 ± 0.60 a | 59.0 ± 11.4 a |
| 0.1 | 30 | 73.33 ± 7.70 ab | 5.19 ± 0.49 a | 52.6 ± 6.7 a |
| 0.2 | 36 | 69.52 ± 9.35 ab | 5.19 ± 0.36 a | 68.4 ± 9.5 a |
| 0.5 | 36 | 89.38 ± 4.38 b | 4.56 ± 0.25 a | 54.0 ± 7.4 a |
| 1.0 | 36 | 76.19 ± 9.52 ab | 5.29 ± 0.32 a | 69.2 ± 8.1 a |
| Explant Type | Treatment (mg L−1) | Explant No. | Mean No. of Explants with Shoots | Regeneration Frequency (%) | Mean No. of Regenerated Shoots per Regenerative Explant |
|---|---|---|---|---|---|
| cotyledon | 2,4-D 0.5 | 94 | 0 | 0.00 | 0 a |
| 2,4-D 1.0 | 88 | 0 | 0.00 | 0 a | |
| 2,4-D 2.0 | 93 | 0 | 0.00 | 0 a | |
| BA 1.0+NAA 0.1 | 166 | 0 | 0.00 | 0 a | |
| BA 1.0+NAA 0.1+AgNO3 5.0 | 162 | 0 | 0.00 | 0 a | |
| TDZ 1.0+NAA 0.1 | 75 | 0 | 0.00 | 0 a | |
| TDZ 1.0+NAA 0.1+AgNO3 5.0 | 73 | 0 | 0.00 | 0 a | |
| hypocotyl | 2,4-D 0.5 | 43 | 0 | 0.00 | 0 a |
| 2,4-D 1.0 | 43 | 0 | 0.00 | 0 a | |
| 2,4-D 2.0 | 42 | 0 | 0.00 | 0 a | |
| BA 1.0+NAA 0.1 | 272 | 25 | 9.19 | 1.68 ± 0.14 bc | |
| BA 1.0+NAA 0.1+AgNO3 5.0 | 166 | 13 | 7.83 | 2.08 ± 0.26 cd | |
| TDZ 1.0+NAA 0.1 | 221 | 14 | 6.33 | 1.5 ± 0.17 b | |
| TDZ 1.0+NAA 0.1+AgNO3 5.0 | 130 | 33 | 25.38 | 2.18 ± 0.16 d | |
| root | 2,4-D 0.5 | 21 | 0 | 0.00 | 0 a |
| 2,4-D 1.0 | 22 | 0 | 0.00 | 0 a | |
| 2,4-D 2.0 | 22 | 0 | 0.00 | 0 a | |
| BA 1.0+NAA 0.1 | 78 | 3 | 3.85 | 1.33 ± 0.33 bcd | |
| BA 1.0+NAA 0.1+AgNO3 5.0 | 81 | 14 | 17.28 | 1.57 ± 0.17 bc | |
| TDZ 1.0+NAA 0.1 | 72 | 5 | 6.94 | 2.0 ± 0.32 bcd | |
| TDZ 1.0+NAA 0.1+AgNO3 5.0 | 62 | 13 | 20.97 | 2.08 ± 0.24 cd | |
| ANOVA Source of Variation | Df | Mean Square | F-Ratio | p-Value | |
| Explant | 2 | 8.67346 | 16.67265 | 0.000000 | |
| Tretman | 6 | 5.06394 | 9.73422 | 0.000000 | |
| Explant × Treatment | 12 | 1.28534 | 2.47076 | 0.005903 | |
| 2,4-D (mg L−1) | No. of Explants | Explants with SE (%) | Mean No. Somatic Embryos per Embryogenic Explant | Callus (%) | Necrotic Explants (%) | Explants without Response (%) |
|---|---|---|---|---|---|---|
| 0.5 | 335 | 71.48 ± 4.29 a | 4.25 ± 0.27 a | 23.39 ± 4.01 a | 0.71 ± 0.51 a | 4.25 ± 0.60 b |
| 1.0 | 226 | 76.64 ± 5.30 a | 5.13 ± 0.25 b | 11.42 ± 4.42 a | 9.38 ± 0.27 b | 2. 56 ± 2.59 ab |
| 2.0 | 213 | 64.54 ± 6.20 a | 4.00 ± 0.36 a | 24.14 ± 4.29 a | 11.32 ± 0.27 b | 0.00 ± 0.00 a |
| Treatment | No. of SEs Established | Mean No. of Germinated SEs | Hyperhydrated SEs (%) |
|---|---|---|---|
| PEG 6000 30 g L−1 | 35 | 0.0 ± 0.0 | - |
| PGR-free | 35 | 0.0 ± 0.0 | - |
| BAP 0.5 mg L−1 | 35 | 6.70 ± 1.6 | 80 |
| KIN 0.1 mg L−1 | 35 | 12.20 ± 1.9 | 0.0 |
| KIN 0.5 mg L−1 | 30 | 22.50 ± 1.4 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banjac, N.; Krstić-Milošević, D.; Mijalković, T.; Petrović, M.; Ćosić, T.; Stanišić, M.; Vinterhalter, B. In Vitro Shoot Multiplication and Regeneration of the Recalcitrant Rocket (Eruca sativa Mill.) Variety Domaća Rukola. Horticulturae 2023, 9, 533. https://doi.org/10.3390/horticulturae9050533
Banjac N, Krstić-Milošević D, Mijalković T, Petrović M, Ćosić T, Stanišić M, Vinterhalter B. In Vitro Shoot Multiplication and Regeneration of the Recalcitrant Rocket (Eruca sativa Mill.) Variety Domaća Rukola. Horticulturae. 2023; 9(5):533. https://doi.org/10.3390/horticulturae9050533
Chicago/Turabian StyleBanjac, Nevena, Dijana Krstić-Milošević, Tatjana Mijalković, Mirjana Petrović, Tatjana Ćosić, Mariana Stanišić, and Branka Vinterhalter. 2023. "In Vitro Shoot Multiplication and Regeneration of the Recalcitrant Rocket (Eruca sativa Mill.) Variety Domaća Rukola" Horticulturae 9, no. 5: 533. https://doi.org/10.3390/horticulturae9050533