
Different Cutting Methods Affect the Quality of Fresh-Cut Cucumbers by Regulating ROS Metabolism
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Treatment
2.2. Colour Parameters, Weight Loss, Respiration Rate, and Firmness Assay
2.3. Total Soluble Solid (TSS), Ascorbic Acid (AsA), and Glutathione (GSH) Content Assay
2.4. Total Phenols, Lignin Content, and PAL Activity Assay
2.5. O2−· and H2O2 Content Assay
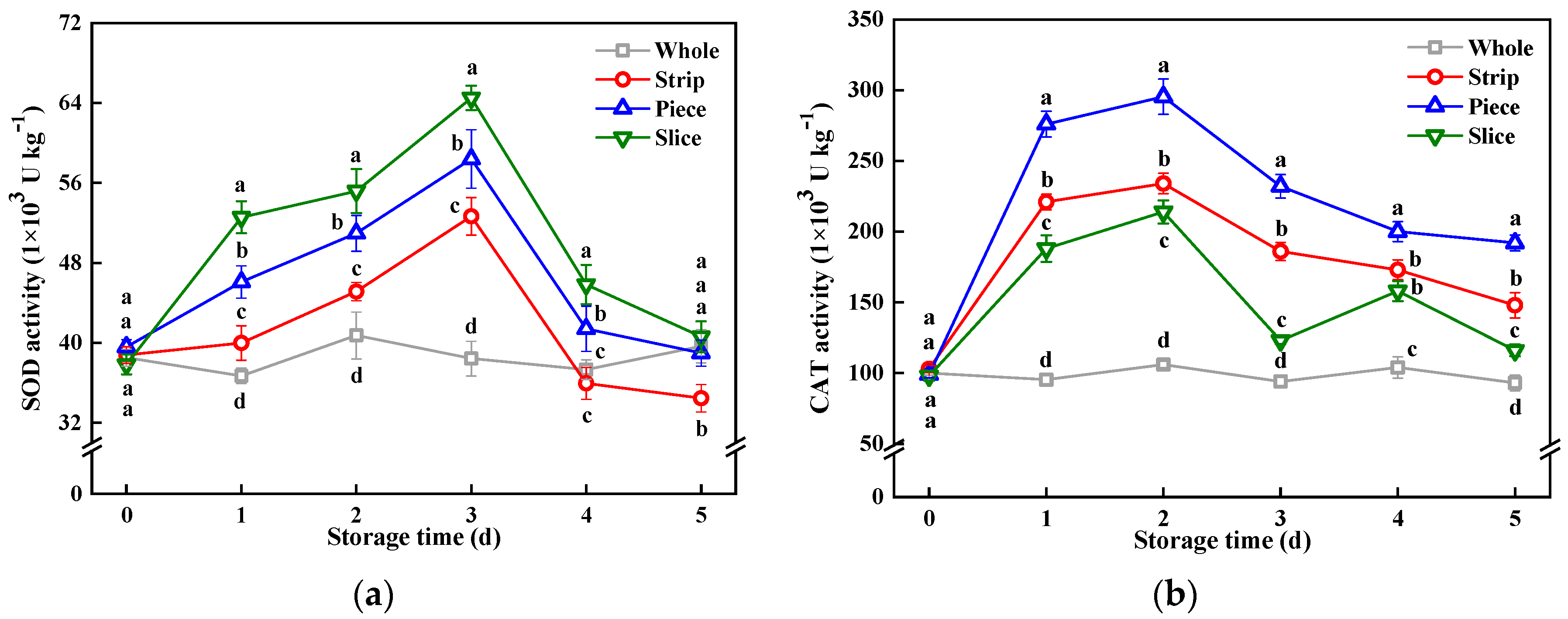
2.6. Antioxidant Enzymes Activity Assay
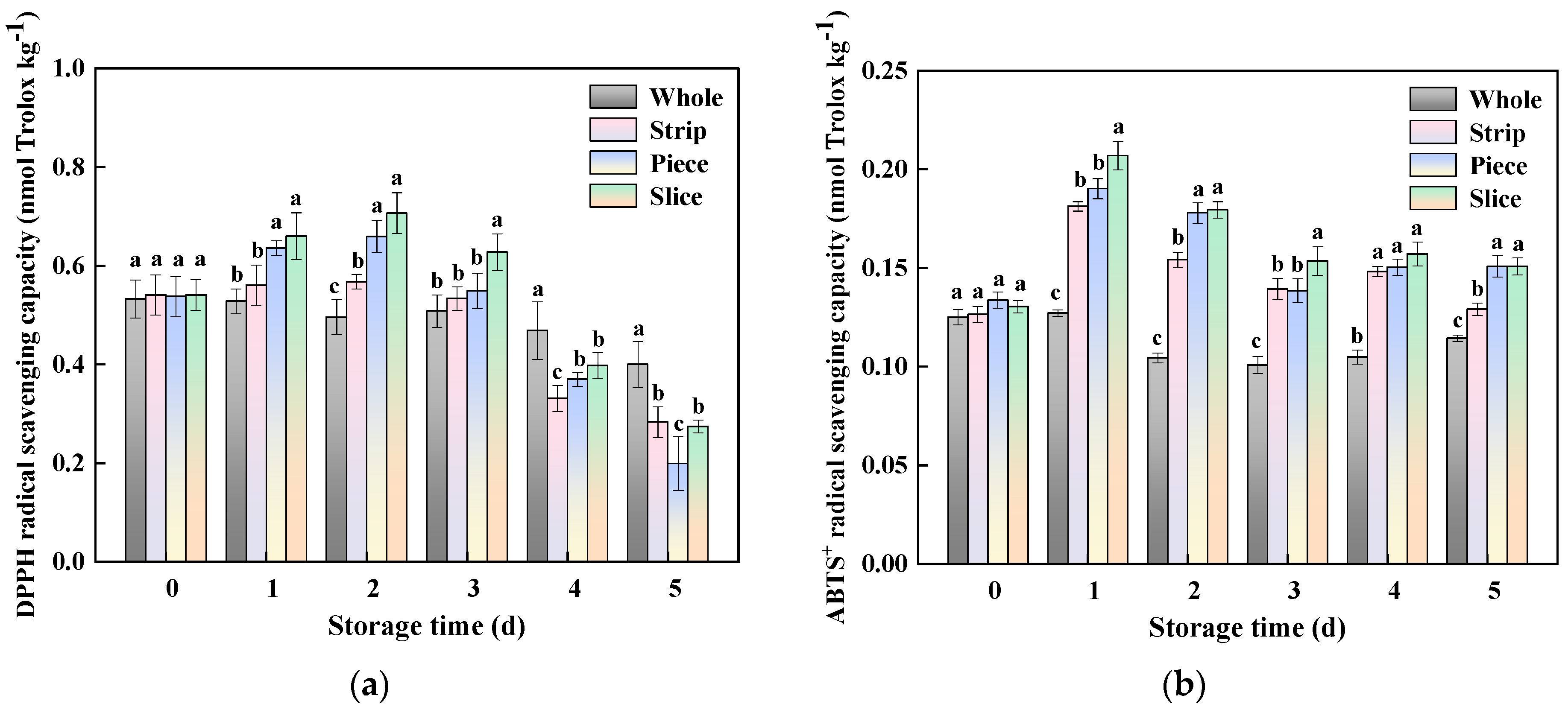
2.7. Antioxidant Activity Assay
2.8. Statistical Assay
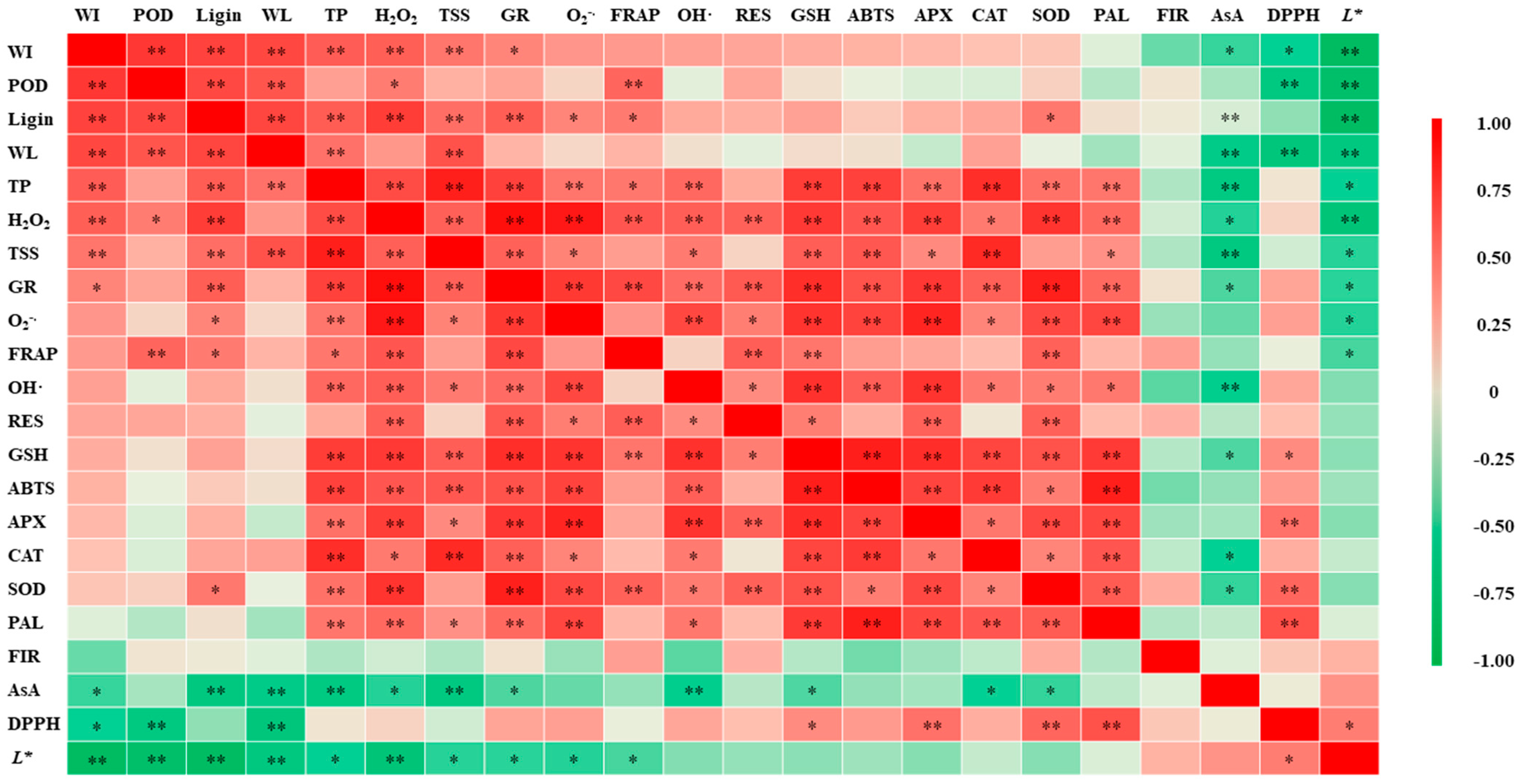
3. Results
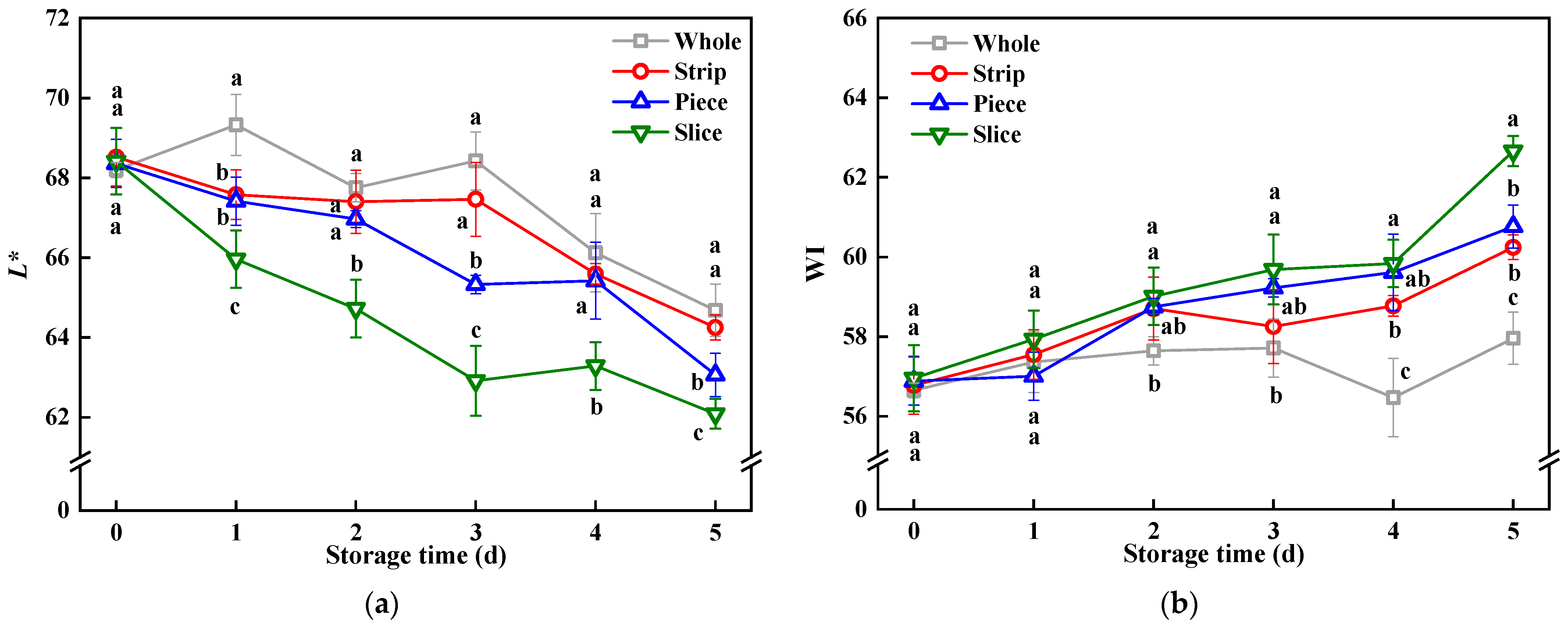
3.1. Colorimetric Values
3.2. Weight Loss and Total Soluble Solid
3.3. Respiration Rate and Firmness
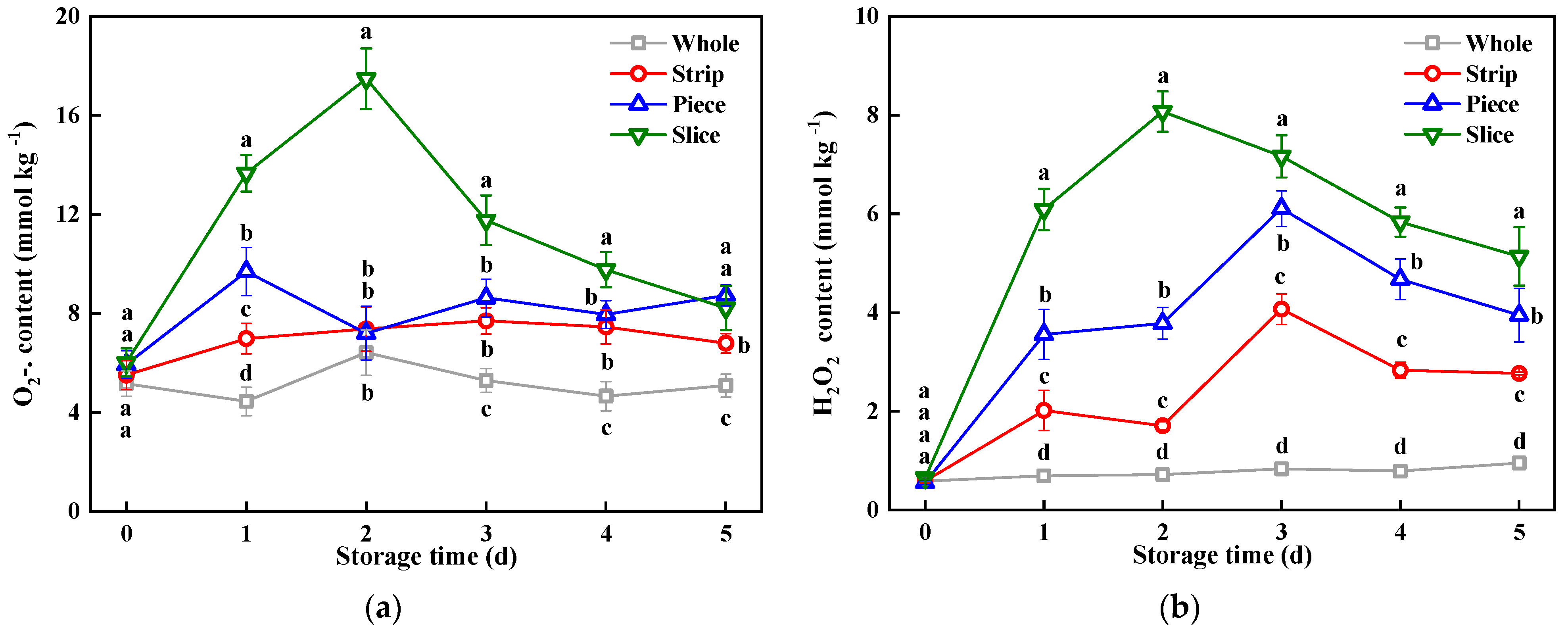
3.4. Reactive Oxygen Species (ROS) Content
3.5. Ascorbic Acid–Glutathione Cycle-Related Indexes
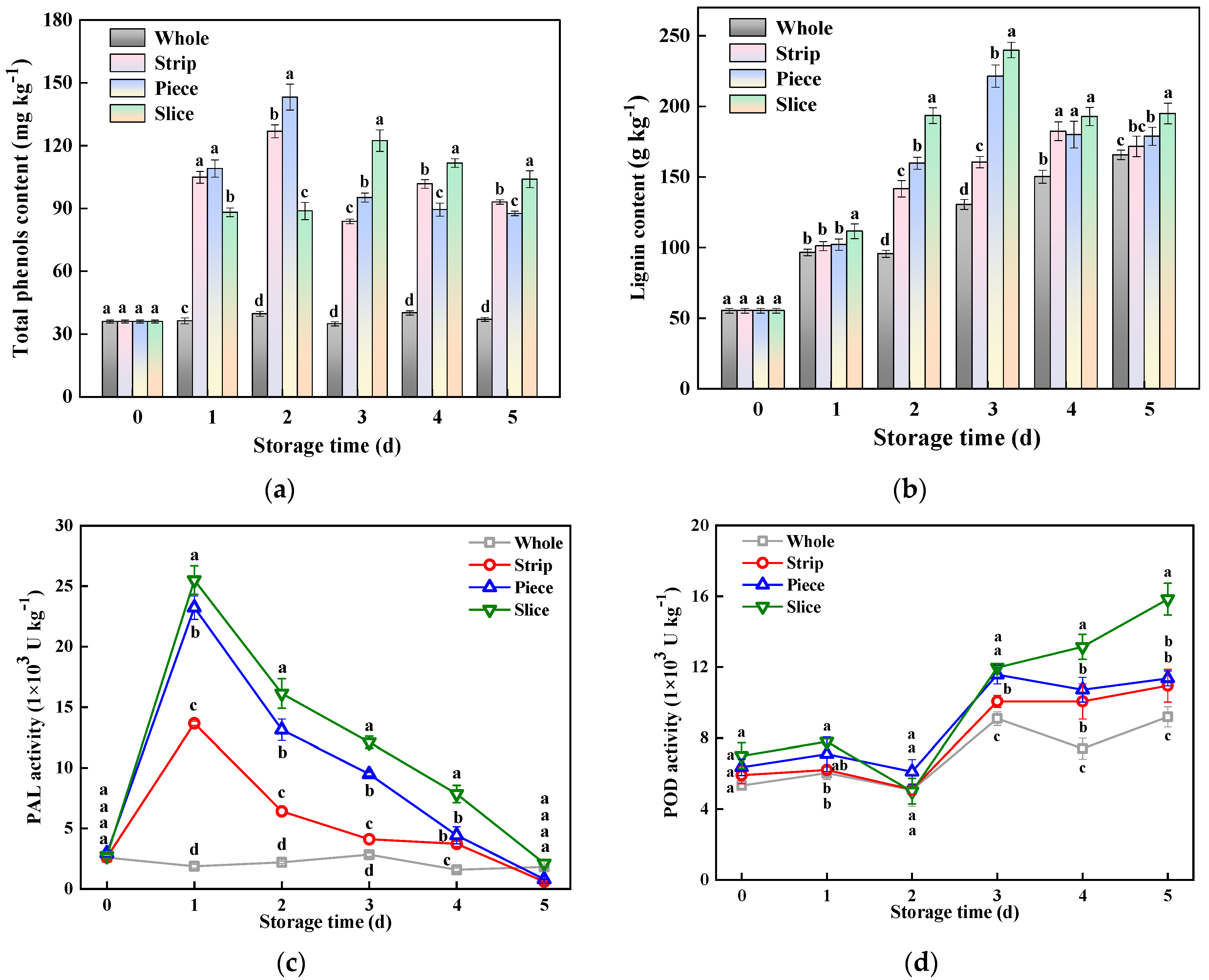
3.6. Phenylpropanoid Metabolism-Related Indexes
3.7. Antioxidant Enzymes Activity
3.8. Antioxidant Capacity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hu, W.; Sarengaowa; Guan, Y.; Feng, K. Biosynthesis of Phenolic Compounds and Antioxidant Activity in Fresh-Cut Fruits and Vegetables. Front. Microbiol. 2022, 13, 906069. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, B.; Li, M.; Fu, X.; Zhao, X.; Min, D.; Li, F.; Zhang, X. Ethylene pretreatment induces phenolic biosynthesis of fresh-cut pitaya fruit by regulating ethylene signaling pathway. Postharvest Biol. Technol. 2022, 192, 112028. [Google Scholar] [CrossRef]
- Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Controlled Abiotic Stresses Revisited: From Homeostasis through Hormesis to Extreme Stresses and the Impact on Nutraceuticals and Quality during Pre- and Postharvest Applications in Horticultural Crops. J. Agric. Food Chem. 2020, 68, 11877–11879. [Google Scholar] [CrossRef] [PubMed]
- Torres-Contreras, A.M.; Senés-Guerrero, C.; Pacheco, A.; González-Agüero, M.; Ramos-Parra, P.A.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Genes differentially expressed in broccoli as an early and late response to wounding stress. Postharvest Biol. Technol. 2018, 145, 172–182. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Santacruz, A.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Postharvest Wounding Stress in Horticultural Crops as a Tool for Designing Novel Functional Foods and Beverages with Enhanced Nutraceutical Content: Carrot Juice as a Case Study. J. Food Sci. 2019, 84, 1151–1161. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Asai, T.; Nishikawa, F.; Ikoma, Y.; Matsumoto, H. Effect of 1-methylcyclopropene on the expression of genes for ascorbate metabolism in postharvest broccoli. Postharvest Biol. Technol. 2010, 58, 121–128. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Yu, J.; Zhao, M.; Ji, Y.; Feng, K.; Sarengaowa; Yang, X. Influence of cut type on quality, antioxidant substances and antioxidant activity of fresh-cut broccoli. Int. J. Food Sci. Technol. 2020, 55, 3019–3030. [Google Scholar] [CrossRef]
- Chen, C.; Hu, W.; He, Y.; Jiang, A.; Zhang, R. Effect of citric acid combined with UV-C on the quality of fresh-cut apples. Postharvest Biol. Technol. 2016, 111, 126–131. [Google Scholar] [CrossRef]
- Li, X.; Long, Q.; Gao, F.; Han, C.; Jin, P.; Zheng, Y. Effect of cutting styles on quality and antioxidant activity in fresh-cut pitaya fruit. Postharvest Biol. Technol. 2017, 124, 1–7. [Google Scholar] [CrossRef]
- Hu, W.; Guan, Y.; Ji, Y.; Yang, X. Effect of cutting styles on quality, antioxidant activity, membrane lipid peroxidation, and browning in fresh-cut potatoes. Food Biosci. 2021, 44, 101435. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Sa, R.; Feng, K.; Zhao, M.; Yu, J.; Ji, Y.; Hou, M.; et al. Effect of Methyl Jasmonate on Phenolic Accumulation in Wounded Broccoli. Molecules 2019, 24, 3537. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Martínez-Hernández, G.B.; del C. Rodríguez, S.; Cao, C.-M.; Cisneros-Zevallos, L. Plants as Biofactories: Physiological Role of Reactive Oxygen Species on the Accumulation of Phenolic Antioxidants in Carrot Tissue under Wounding and Hyperoxia Stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Li, Y.; Hu, J.; Pang, L.; Fan, H. Browning inhibition and quality preservation of fresh-cut romaine lettuce exposed to high intensity light. Innov. Food Sci. Emerg. Technol. 2012, 14, 70–76. [Google Scholar] [CrossRef]
- Oliu, G.O.; Aguiló-Aguayo, I.; Martín-Belloso, O.; Soliva-Fortuny, R. Effects of pulsed light treatments on quality and antioxidant properties of fresh-cut mushrooms (Agaricus bisporus). Postharvest Biol. Technol. 2010, 56, 216–222. [Google Scholar] [CrossRef]
- Berno, N.D.; Tezotto-Uliana, J.V.; Dias, C.T.D.S.; Kluge, R.A. Storage temperature and type of cut affect the biochemical and physiological characteristics of fresh-cut purple onions. Postharvest Biol. Technol. 2014, 93, 91–96. [Google Scholar] [CrossRef]
- Viña, S.Z.; Chaves, A.R. Antioxidant responses in minimally processed celery during refrigerated storage. Food Chem. 2006, 94, 68–74. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, Z.; Zhang, X.; Zheng, S.; Wang, J.; Mo, J. Alleviating effects of exogenous melatonin on salt stress in cucumber. Sci. Hortic. 2020, 262, 109070. [Google Scholar] [CrossRef]
- Fan, K.; Zhang, M.; Jiang, F. Ultrasound treatment to modified atmospheric packaged fresh-cut cucumber: Influence on microbial inhibition and storage quality. Ultrason. Sonochem. 2019, 54, 162–170. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Lei, X.; Tian, W.; Cao, J.; Jiang, W.; Wang, M. Effects of Wax Coating on the Moisture Loss of Cucumbers at Different Storage Temperatures. J. Food Qual. 2018, 2018, 9351821. [Google Scholar] [CrossRef]
- Han, C.; Li, J.; Jin, P.; Li, X.; Wang, L.; Zheng, Y. The effect of temperature on phenolic content in wounded carrots. Food Chem. 2017, 215, 116–123. [Google Scholar] [CrossRef]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Biosynthesis of phenolic antioxidants in carrot tissue increases with wounding intensity. Food Chem. 2012, 134, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Sarengaowa; Hu, W.; Jiang, A.; Xiu, Z.; Feng, K. Effect of thyme oil-alginate-based coating on quality and microbial safety of fresh-cut apples. J. Sci. Food Agric. 2018, 98, 2302–2311. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Xu, D.; Liu, C.; Chen, C.; Tian, M.; Jiang, A. Ascorbic acid treatment inhibits wound healing of fresh-cut potato strips by controlling phenylpropanoid metabolism. Postharvest Biol. Technol. 2021, 181, 111644. [Google Scholar] [CrossRef]
- Hu, W.; Jiang, A.; Tian, M.; Liu, C.; Wang, Y. Effect of ethanol treatment on physiological and quality attributes of fresh-cut eggplant. J. Sci. Food Agric. 2010, 90, 1323–1326. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; Toivonen, P.M. Quality of fresh-cut fruits and vegetables as affected by exposure to abiotic stress. Postharvest Biol. Technol. 2008, 48, 155–162. [Google Scholar] [CrossRef]
- Lai, Y.; Wang, W.; Zhao, J.; Tu, S.; Yin, Y.; Ye, L. Chitosan Na-montmorillonite films incorporated with citric acid for prolonging cherry tomatoes shelf life. Food Packag. Shelf Life 2022, 33, 100879. [Google Scholar] [CrossRef]
- Han, C.; Ji, Y.; Li, M.; Li, X.; Jin, P.; Zheng, Y. Influence of wounding intensity and storage temperature on quality and antioxidant activity of fresh-cut Welsh onions. Sci. Hortic. 2016, 212, 203–209. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, Y.; Jia, R.; Wang, Q.; Lou, X.; Shi, J. Sodium chloride combined with polypropylene film can maintain the quality of fresh-cut ginger. Food Packag. Shelf Life 2020, 25, 100541. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, H.; Zhang, J.; Sheng, Z.; Cao, J.; Jiang, W. Different molecular weights chitosan coatings delay the senescence of postharvest nectarine fruit in relation to changes of redox state and respiratory pathway metabolism. Food Chem. 2020, 289, 160–168. [Google Scholar] [CrossRef]
- Wang, D.; Ma, Q.; Li, D.; Li, W.; Li, L.; Aalim, H.; Luo, Z. Moderation of respiratory cascades and energy metabolism of fresh-cut pear fruit in response to high CO2 controlled atmosphere. Postharvest Biol. Technol. 2021, 172, 111379. [Google Scholar] [CrossRef]
- Li, F.; Min, D.; Ren, C.; Dong, L.; Shu, P.; Cui, X.; Zhang, X. Ethylene altered fruit cuticular wax, the expression of cuticular wax synthesis-related genes and fruit quality during cold storage of apple (Malus domestica Borkh. c.v. Starkrimson) fruit. Postharvest Biol. Technol. 2019, 149, 58–65. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; González-Agüero, M.; Cisneros-Zevallos, L. Cross-talk between signaling pathways: The link between plant secondary metabolite production and wounding stress response. Sci. Rep. 2015, 5, 8608. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, C.; Moraes, K.; Pérez-Gago, M.; Palou, L.; Maraschin, M.; Monteiro, A. Effect of active modified atmosphere and cold storage on the postharvest quality of cherry tomatoes. Postharvest Biol. Technol. 2015, 109, 73–81. [Google Scholar] [CrossRef]
- Zhou, F.; Jiang, A.; Feng, K.; Gu, S.; Xu, D.; Hu, W. Effect of methyl jasmonate on wound healing and resistance in fresh-cut potato cubes. Postharvest Biol. Technol. 2019, 157, 110958. [Google Scholar] [CrossRef]
- Barrett, D.M.; Beaulieu, J.; Shewfelt, R. Color, Flavor, Texture, and Nutritional Quality of Fresh-Cut Fruits and Vegetables: Desirable Levels, Instrumental and Sensory Measurement, and the Effects of Processing. Crit. Rev. Food Sci. Nutr. 2010, 50, 369–389. [Google Scholar] [CrossRef]
- Zhao, P.; Li, W.; Zhen, C.; Wang, K.; Qin, Z.; Gao, H. Transcriptomic analysis of the effects of γ-aminobutyric acid treatment on browning and induced disease resistance in fresh-cut apples. Postharvest Biol. Technol. 2021, 181, 111686. [Google Scholar] [CrossRef]
- Nishikawa, F.; Kato, M.; Hyodo, H.; Ikoma, Y.; Sugiura, M.; Yano, M. Ascorbate metabolism in harvested broccoli. J. Exp. Bot. 2003, 54, 2439–2448. [Google Scholar] [CrossRef]
- Xia, F.; Cheng, H.; Chen, L.; Zhu, H.; Mao, P.; Wang, M. Influence of exogenous ascorbic acid and glutathione priming on mitochondrial structural and functional systems to alleviate aging damage in oat seeds. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef]
- Li, Z.; Li, B.; Li, M.; Fu, X.; Zhao, X.; Min, D.; Li, F.; Li, X.; Zhang, X. Hot air pretreatment alleviates browning of fresh-cut pitaya fruit by regulating phenylpropanoid pathway and ascorbate-glutathione cycle. Postharvest Biol. Technol. 2022, 190, 111954. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Come, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiol. Plant. 2010, 97, 104–110. [Google Scholar] [CrossRef]
- Mori, T.; Terai, H.; Yamauchi, N.; Suzuki, Y. Effects of postharvest ethanol vapor treatment on the ascorbate–glutathione cycle in broccoli florets. Postharvest Biol. Technol. 2009, 52, 134–136. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, S.; Ma, Y.; Zhao, W.; Wang, P.; Zhao, S.; Wang, D. Ascorbic acid prevents yellowing of fresh-cut yam by regulating pigment biosynthesis and energy metabolism. Food Res. Int. 2022, 157, 111424. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, X.; Li, L.; Aghdam, M.S.; Wei, X.; Liu, J.; Xu, Y.; Luo, Z. Elevated CO2 delayed the chlorophyll degradation and anthocyanin accumulation in postharvest strawberry fruit. Food Chem. 2019, 285, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zhu, Y.; Li, Z.; Bi, Y.; Yang, J.; Zhang, J.; Prusky, D. Preharvest multiple fungicide stroby sprays promote wound healing of harvested potato tubers by activating phenylpropanoid metabolism. Postharvest Biol. Technol. 2021, 171, 111328. [Google Scholar] [CrossRef]
- Zheng, X.; Jiang, H.; Bi, Y.; Wang, B.; Wang, T.; Li, Y.; Gong, D.; Wei, Y.; Li, Z.; Prusky, D. Comparison of wound healing abilities of four major cultivars of potato tubers in China. Postharvest Biol. Technol. 2020, 164, 111167. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Zhao, M.; Yu, J.; Ji, Y.; Sarengaowa; Yang, X.; Feng, K. The effect of cutting style on the biosynthesis of phenolics and cellular antioxidant capacity in wounded broccoli. Food Res. Int. 2020, 137, 109565. [Google Scholar] [CrossRef]
- Villarreal-García, D.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as Biofactories: Postharvest Stress-Induced Accumulation of Phenolic Compounds and Glucosinolates in Broccoli Subjected to Wounding Stress and Exogenous Phytohormones. Front. Plant Sci. 2016, 7, 45. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Xu, Y.; Sarengaowa; Ji, Y.; Yang, X.; Feng, K. Proteomic analysis validates previous findings on wounding-responsive plant hormone signaling and primary metabolism contributing to the biosynthesis of secondary metabolites based on metabolomic analysis in harvested broccoli (Brassica oleracea L. var. italica). Food Res. Int. 2021, 145, 110388. [Google Scholar] [CrossRef]
- Han, C.; Jin, P.; Li, M.; Wang, L.; Zheng, Y. Physiological and Transcriptomic Analysis Validates Previous Findings of Changes in Primary Metabolism for the Production of Phenolic Antioxidants in Wounded Carrots. J. Agric. Food Chem. 2017, 65, 7159–7167. [Google Scholar] [CrossRef]
- Odriozola-Serrano, I.; Soliva-Fortuny, R.; Martín-Belloso, O. Effect of minimal processing on bioactive compounds and color attributes of fresh-cut tomatoes. LWT-Food Sci. Technol. 2008, 41, 217–226. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Xu, Y.; Yang, X.; Ji, Y.; Feng, K. Sarengaowa Metabolomics and physiological analyses validates previous findings on the mechanism of response to wounding stress of different intensities in broccoli. Food Res. Int. 2020, 140, 110058. [Google Scholar] [CrossRef] [PubMed]
- Zha, Z.; Tang, R.; Wang, C.; Li, Y.-L.; Liu, S.; Wang, L.; Wang, K. Riboflavin inhibits browning of fresh-cut apples by repressing phenolic metabolism and enhancing antioxidant system. Postharvest Biol. Technol. 2022, 187, 111867. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hernández, G.B.; Artés-Hernández, F.; Gomez, P.; Formica, A.C.; Artés, F. Combination of electrolysed water, UV-C and superatmospheric O2 packaging for improving fresh-cut broccoli quality. Postharvest Biol. Technol. 2013, 76, 125–134. [Google Scholar] [CrossRef]
- Guan, Y.; Ji, Y.; Yang, X.; Pang, L.; Cheng, J.; Lu, X.; Zheng, J.; Yin, L.; Hu, W. Antioxidant activity and microbial safety of fresh-cut red cabbage stored in different packaging films. LWT Food Sci. Technol. 2023, 175, 114478. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Time (Day) | Treatment | Total Soluble Solid (%) | Weight Loss (%) | Respiration Rate (mg kg−1 h−1) | Firmness (N) | Ascorbic Acid (mg kg−1) | Glutathione (mg kg−1) |
|---|---|---|---|---|---|---|---|
| 0 | Whole | 0.81 ± 0.03 a | 0 | 51.64 ± 1.68 a | 21.76 ± 0.90 a | 19.97 ± 1.04 a | 0.11 ± 0.004 a |
| Stripped | 0.82 ± 0.05 a | 0 | 53.36 ± 0.98 a | 21.83 ± 0.84 a | 20.81 ± 0.94 a | 0.12 ± 0.007 a | |
| Pieced | 0.77 ± 0.09 a | 0 | 52.09 ± 2.31 a | 20.10 ± 1.21 a | 21.93 ± 0.80 a | 0.12 ± 0.008 a | |
| Sliced | 0.82 ± 0.05 a | 0 | 52.66 ± 0.75 a | 20.64 ± 0.95 a | 20.44 ± 1.31 a | 0.11 ± 0.004 a | |
| 1 | Whole | 0.75 ± 0.05 b | 0.09 ± 0.01 c | 47.69 ± 1.39 b | 19.84 ± 0.47 a | 20.17 ± 0.75 a | 0.10 ± 0.002 c |
| Stripped | 0.98 ± 0.04 a | 0.29 ± 0.05 a | 36.11 ± 1.20 c | 17.05 ± 0.61 b | 21.05 ± 1.41 a | 0.13 ± 0.002 b | |
| Pieced | 0.98 ± 0.04 a | 0.24 ± 0.05 ab | 44.80 ± 2.44 b | 16.83 ± 1.02 b | 20.64 ± 1.51 a | 0.13 ± 0.005 b | |
| Sliced | 0.98 ± 0.05 a | 0.12 ± 0.08 b | 56.18 ± 0.92 a | 14.88 ± 1.49 c | 15.62 ± 0.97 b | 0.15 ± 0.007 a | |
| 2 | Whole | 0.78 ± 0.08 c | 0.16 ± 0.02 d | 55.30 ± 0.66 b | 18.71 ± 0.76 a | 18.69 ± 0.91 a | 0.11 ± 0.003 c |
| Stripped | 1.03 ± 0.03 ab | 0.60 ± 0.07 a | 40.07 ± 0.70 c | 15.82 ± 0.80 b | 10.26 ± 0.81 c | 0.13 ± 0.004 b | |
| Pieced | 1.09 ± 0.04 a | 0.46 ± 0.05 b | 51.81 ± 2.25 b | 15.09 ± 0.90 b | 15.04 ± 1.32 b | 0.14 ± 0.005 a | |
| Sliced | 0.98 ± 0.06 b | 0.31 ± 0.01 c | 59.47 ± 1.86 a | 11.93 ± 0.89 c | 16.02 ± 1.41 b | 0.15 ± 0.004 a | |
| 3 | Whole | 0.73 ±0.04 d | 0.25 ± 0.04 d | 36.32 ± 0.94 c | 19.49 ± 0.94 d | 19.34 ± 1.07 a | 0.11 ± 0.004 b |
| Stripped | 0.88 ± 0.04 c | 0.83 ± 0.09 a | 59.49 ± 1.02 b | 33.84 ± 0.83 a | 14.91 ± 0.90 b | 0.13 ± 0.005 a | |
| Pieced | 1.08 ± 0.07 a | 0.65 ± 0.03 b | 62.31 ± 1.40 b | 32.04 ± 0.65 b | 15.00 ± 1.06 b | 0.13 ± 0.006 a | |
| Sliced | 0.92 ± 0.07 b | 0.43 ± 0.09 c | 72.30 ± 0.98 a | 25.68 ± 0.84 c | 16.20 ± 1.29 b | 0.13 ± 0.007 a | |
| 4 | Whole | 0.82 ± 0.07 b | 0.33 ± 0.04 c | 30.15 ± 0.90 c | 20.35 ± 1.28 b | 18.61 ± 1.21 a | 0.11 ± 0.004 c |
| Stripped | 1.01 ± 0.03 a | 1.01 ± 0.10 a | 59.04 ± 1.28 a | 24.17 ± 0.67 a | 15.43 ± 0.73 b | 0.13 ± 0.003 b | |
| Pieced | 1.01 ± 0.04 a | 0.90 ± 0.07 a | 54.20 ± 1.09 b | 16.04 ± 0.84 c | 16.58 ± 1.41 ab | 0.12 ± 0.004 b | |
| Sliced | 0.93 ± 0.06 a | 0.57 ± 0.09 b | 59.62 ± 2.06 a | 15.36 ± 1.19 c | 18.17 ± 0.98 a | 0.14 ± 0.006 a | |
| 5 | Whole | 0.78 ± 0.04 c | 0.38 ± 0.03 d | 32.17 ± 0.93 c | 19.34 ± 1.23 a | 17.85 ± 1.47 a | 0.10 ± 0.002 d |
| Stripped | 1.10 ± 0.03 a | 1.26 ± 0.03 a | 35.87 ± 1.72 b | 15.76 ± 1.12 b | 16.99 ± 1.03 a | 0.11 ± 0.003 c | |
| Pieced | 1.01 ± 0.04 b | 1.13 ± 0.03 b | 37.65 ± 0.92 b | 12.69 ± 0.68 c | 15.86 ± 0.85 a | 0.12 ± 0.005 b | |
| Sliced | 0.93 ± 0.08 b | 0.72 ± 0.01 c | 66.46 ± 2.16 a | 11.54 ± 1.00 c | 17.76 ± 1.22 a | 0.13 ± 0.004 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, Y.; Hu, W.; Wang, L.; Yang, B. Different Cutting Methods Affect the Quality of Fresh-Cut Cucumbers by Regulating ROS Metabolism. Horticulturae 2023, 9, 514. https://doi.org/10.3390/horticulturae9040514
Guan Y, Hu W, Wang L, Yang B. Different Cutting Methods Affect the Quality of Fresh-Cut Cucumbers by Regulating ROS Metabolism. Horticulturae. 2023; 9(4):514. https://doi.org/10.3390/horticulturae9040514
Chicago/Turabian StyleGuan, Yuge, Wenzhong Hu, Lei Wang, and Bailu Yang. 2023. "Different Cutting Methods Affect the Quality of Fresh-Cut Cucumbers by Regulating ROS Metabolism" Horticulturae 9, no. 4: 514. https://doi.org/10.3390/horticulturae9040514