
Influence of the 135 bp Intron on Stilbene Synthase VaSTS11 Transgene Expression in Cell Cultures of Grapevine and Different Plant Generations of Arabidopsis thaliana

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
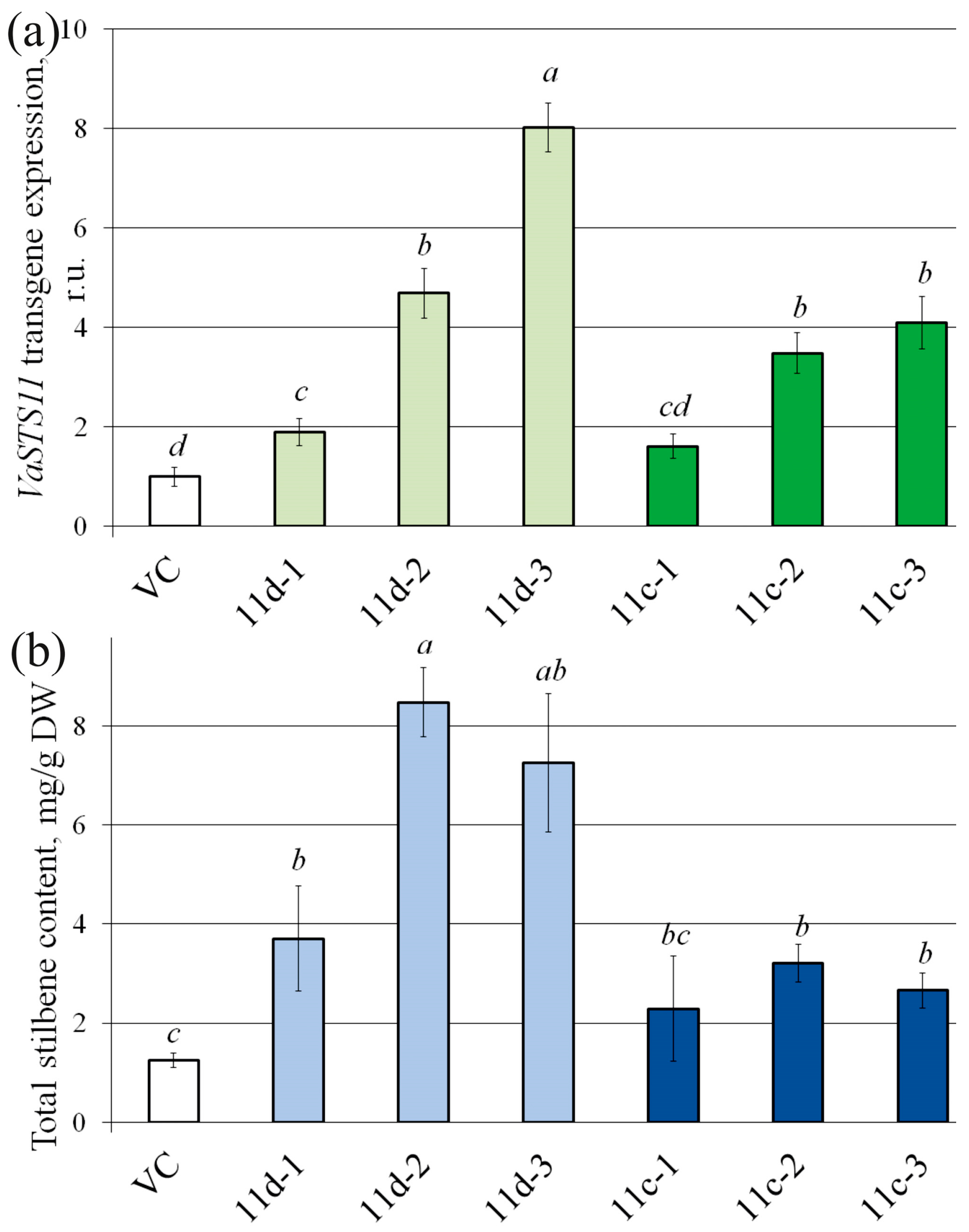
2.1. VaSTS11 Transgene Expression in the VaSTS11-Transgenic A. thaliana
2.2. Stilbene Content in the VaSTS11-Transgenic A. thaliana
2.3. VaSTS11 Cytosine Methylation in Transgenic A. thaliana
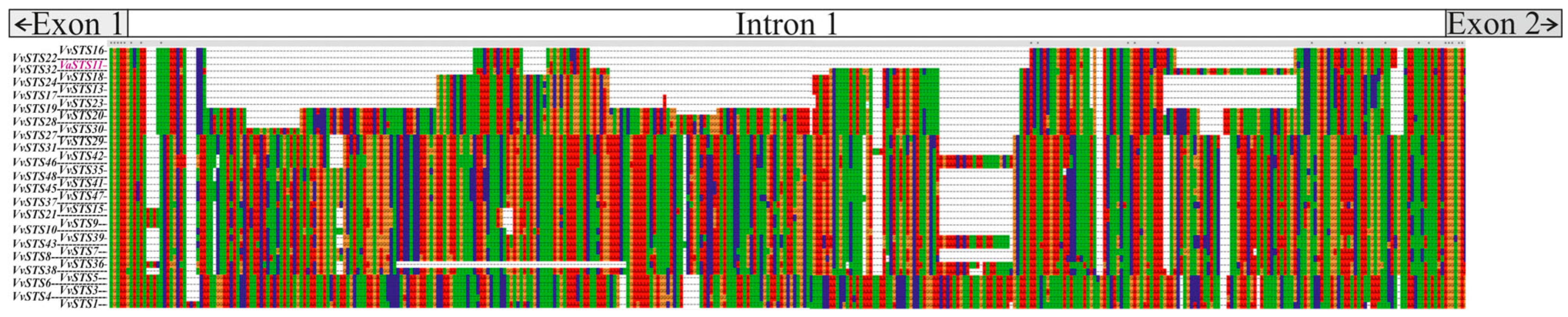
2.4. Search for Consensus Patterns of Plant Regulatory Sequences in the VaSTS11 Intron Using NSITE-PL
2.5. Stilbene and Biomass Accumulation in the Grapevine VaSTS11-Transgenic Cell Lines
3. Conclusions
4. Materials and Methods
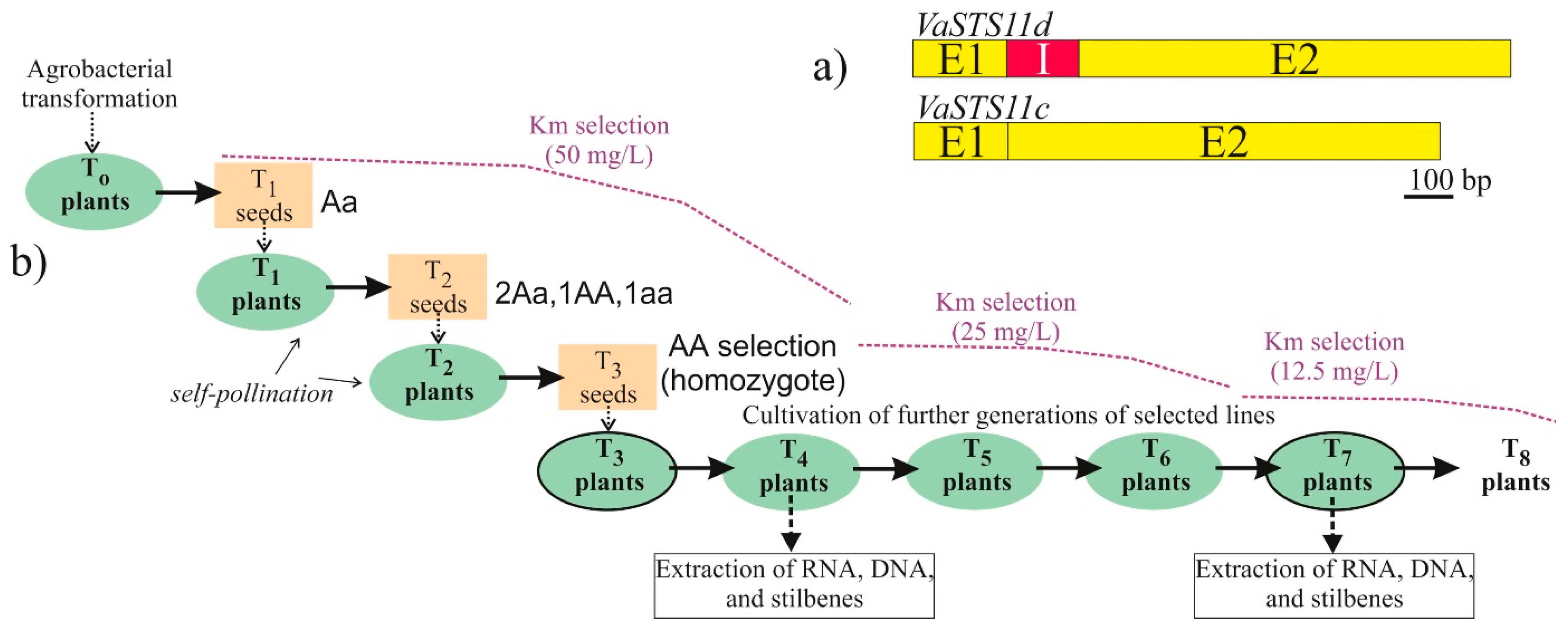
4.1. Plant Material and Cell Cultures
4.2. Overexpression of VaSTS11c and VaSTS11d in Arabidopsis Plants and Cell Cultures of V. amurensis
4.3. HPLC and Mass Spectrometry Stilbene Analysis
4.4. Nucleic Acid Purification and RT-qPCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends Plant Sci. 2008, 13, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.; Brosnan, C.A.; Rothnagel, J.A.; Carroll, B.J. RNA decay and RNA silencing in plants: Competition or collaboration? Front. Plant Sci. 2011, 2, 99. [Google Scholar] [CrossRef]
- Dalakouras, A.; Lauter, A.; Bassler, A.; Krczal, G.; Wassenegger, M. Transient expression of intron-containing transgenes generates non-spliced aberrant pre-mRNAs that are processed into siRNAs. Planta 2019, 249, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.; Carroll, B.J. SERRATE is required for intron suppression of RNA silencing in Arabidopsis. Plant Signal Behav. 2011, 6, 2035–2037. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Solovyev, V.V.; Shahmuradov, I.A.; Salamov, A.A. Identification of promoter regions and regulatory sites. Methods Mol. Biol. 2010, 674, 57–83. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.-J. Cancer chemopreventive and therapeutic potential of resveratrol: Mechanistic perspectives. Cancer Lett. 2008, 269, 243–261. [Google Scholar] [CrossRef]
- Vingtdeux, V.; Dreses-Werringloer, U.; Zhao, H.; Davies, P.; Marambaud, P. Therapeutic potential of resveratrol in Alzheimer’s disease. BMC Neurosci. 2008, 9, S6. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering, and biological functions of stilbene phytoalexins in plants. Biofactors 2010, 36, 331–341. [Google Scholar] [CrossRef]
- Kiselev, K.V. Perspectives for production and application of resveratrol. Appl. Microbiol. Biotechnol. 2011, 90, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Langcake, P.; Pryce, R.J. A new class of phytoalexins from grapevines. Experientia 1977, 33, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Dubrovina, A.S.; Kiselev, K.V. Regulation of stilbene biosynthesis in plants. Planta 2017, 346, 597–623. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.B.; Bowman, M.E.; Ferrer, J.-L.; Schröder, J.; Noel, J.P. An Aldol Switch Discovered in Stilbene Synthases Mediates Cyclization Specificity of Type III Polyketide Synthases. Chem. Biol. 2004, 11, 1179–1194. [Google Scholar] [CrossRef]
- Schmidlin, L.; Poutaraud, A.; Claudel, P.; Mestre, P.; Prado, E.; Santos-Rosa, M.; Wiedemann-Merdinoglu, S.; Karst, F.; Merdinoglu, D.; Hugueney, P. A stress-inducible resveratrol O-methyltransferase involved in the biosynthesis of pterostilbene in grapevine. Plant Physiol. 2008, 148, 1630–1639. [Google Scholar] [CrossRef]
- Hall, D.; De Luca, V. Mesocarp localization of a bi-functional resveratrol/hydroxycinnamic acid glucosyltransferase of Concord grape (Vitis labrusca). Plant J. 2007, 49, 579–591. [Google Scholar] [CrossRef]
- Dry, I.B.; Robinson, S. Molecular cloning and characterisation of grape berry polyphenol oxidase. Plant Mol. Biol. 1994, 26, 495–502. [Google Scholar] [CrossRef]
- Zhang, X.; Henriques, R.; Lin, S.-S.; Niu, Q.-W.; Chua, N.-H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Dubrovina, A.S. Overexpression of stilbene synthase genes to modulate the properties of plants and plant cell cultures. Biotechnol. Appl. Biochem. 2020, 68, 13–19. [Google Scholar] [CrossRef]
- Yu, C.K.Y.; Lam, C.N.W.; Springob, K.; Schmidt, J.; Chu, I.K.; Lo, C. Constitutive Accumulation of cis -piceid in Transgenic Arabidopsis Overexpressing a Sorghum Stilbene Synthase Gene. Plant Cell Physiol. 2006, 47, 1017–1021. [Google Scholar] [CrossRef]
- Lo, C.; Le Blanc, J.C.Y.; Yu, C.K.Y.; Sze, K.H.; Ng, D.C.M.; Chu, I.K. Detection, characterization, and quantification of resveratrol glycosides in transgenic Arabidopsis over-expressing a sorghum stilbene synthase gene by liquid chromatog-raphy/tandem mass spectrometry. Rapid Commun. Mass. Spectrom. 2007, 21, 4101–4108. [Google Scholar] [CrossRef] [PubMed]
- Rupprich, N.; Hildebrand, H.; Kindl, H. Substrate specificity in vivo and in vitro in the formation of stilbenes. Biosynthesis of rhaponticin. Arch. Biochem. Biophys. 1980, 200, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Shumakova, O.A.; Manyakhin, A.Y.; Kiselev, K.V. Resveratrol Content and Expression of Phenylalanine Ammonia-lyase and Stilbene Synthase Genes in Cell Cultures of Vitis amurensis Treated with Coumaric Acid. Appl. Biochem. Biotechnol. 2011, 165, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Ogneva, Z.; Aleynova, O.; Suprun, A.; Karetin, Y.; Dubrovina, A.; Kiselev, K. Tolerance of Arabidopsis thaliana plants overexpressing grapevine VaSTS1 or VaSTS7 genes to cold, heat, drought, salinity, and ultraviolet irradiation. Biol. Plant 2021, 65, 111–117. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V. Age-associated alterations in the somatic mutation and DNA methylation levels in plants. Plant Biol. 2015, 18, 185–196. [Google Scholar] [CrossRef]
- Kwon, H.B.; Park, S.C.; Peng, H.P.; Goodman, H.M.; Dewdney, J.; Shih, M.C. Identification of a Light-Responsive Region of the Nuclear Gene Encoding the B Subunit of Chloroplast Glyceraldehyde 3-Phosphate Dehydrogenase from Arabidopsis thaliana. Plant Physiol. 1994, 105, 357–367. [Google Scholar] [CrossRef]
- Shih, M.-C.; Heinrich, P.; Goodman, H.M. Cloning and chromosomal mapping of nuclear genes encoding chloroplast and cytosolic glyceraldehyde-3-phosphate-dehydrogenase from Arabidopsis thaliana. Gene 1991, 104, 133–138. [Google Scholar] [CrossRef]
- Hao, D.; Jin, L.; Wen, X.; Yu, F.; Xie, Q.; Guo, H. The RING E3 ligase SDIR1 destabilizes EBF1/EBF2 and modulates the ethylene response to ambient temperature fluctuations in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2024592118. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Li, K.; Liu, H.; Lin, C. Multiple bHLH Proteins form Heterodimers to Mediate CRY2-Dependent Regulation of Flowering-Time in Arabidopsis. PLoS Genet. 2013, 9, e1003861. [Google Scholar] [CrossRef]
- Klein, P.; Seidel, T.; Stöcker, B.; Dietz, K.J. The membrane-tethered transcription factor ANAC089 serves as redox-dependent suppressor of stromal ascorbate peroxidase gene expression. Front. Plant Sci. 2012, 3, 247. [Google Scholar] [CrossRef]
- Aleynova-Shumakova, O.A.; Dubrovina, A.S.; Manyakhin, A.Y.; Karetin, Y.A.; Kiselev, K.V. VaCPK20 gene overexpression significantly increased resveratrol content and expression of stilbene synthase genes in cell cultures of Vitis amurensis Rupr. Appl. Microbiol. Biotechnol. 2014, 98, 5541–5549. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Aleynova, O.A. Influence of overexpression of stilbene synthase VaSTS7 gene on resveratrol production in transgenic cell cultures of grape Vitis amurensis Rupr. Appl. Biochem. Microbiol. 2016, 52, 56–60. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Grigorchuk, V.P.; Dubrovina, A.S.; Rybin, V.G.; Kiselev, K.V. Stilbene accumulation in cell cultures of Vitis amurensis Rupr. overexpressing VaSTS1, VaSTS2, and VaSTS7 genes. Plant Cell Tissue Organ Cult. 2016, 125, 329–339. [Google Scholar] [CrossRef]
- Chu, M.; Pedreño, M.; Alburquerque, N.; Faize, L.; Burgos, L.; Almagro, L. A new strategy to enhance the biosynthesis of trans-resveratrol by overexpressing stilbene synthase gene in elicited Vitis vinifera cell cultures. Plant Physiol. Biochem. 2017, 113, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Suprun, A.R.; Ogneva, Z.V.; Dubrovina, A.S.; Kiselev, K.V. Effect of spruce PjSTS1a, PjSTS2, or PjSTS3 gene overexpression on stilbene biosynthesis in callus cultures of Vitis amurensis Rupr. Biotechnol. Appl. Biochem. 2020, 67, 234–239. [Google Scholar] [CrossRef]
- Gelvin, S.B. Agrobacterium-mediated plant transformation: The biology behind the “gene-jockeying” tool. Microbiol. Mol. Biol. Rev. 2003, 67, 16–37. [Google Scholar] [CrossRef]
- Tyunin, A.P.; Suprun, A.R.; Nityagovsky, N.N.; Manyakhin, A.Y.; Karetin, Y.A.; Dubrovina, A.S.; Kiselev, K.V. The effect of explant origin and collection season on stilbene biosynthesis in cell cultures of Vitis amurensis Rupr. Plant Cell Tissue Organ Cult. 2019, 136, 189–196. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Manyakhin, A.Y.; Zhuravlev, Y.N.; Kiselev, K.V. Resveratrol content and expression of phenylalanine ammonia-lyase and stilbene synthase genes in rolC transgenic cell cultures of Vitis amurensis. Appl. Microbiol. Biotechnol. 2010, 88, 727–736. [Google Scholar] [CrossRef]
- Tzfira, T.; Tian, G.-W.; Lacroix, B.; Vyas, S.; Li, J.; Leitner-Dagan, Y.; Krichevsky, A.; Taylor, T.; Vainstein, A.; Citovsky, V. pSAT vectors: A modular series of plasmids for autofluorescent protein tagging and expression of multiple genes in plants. Plant Mol. Biol. 2005, 57, 503–516. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr., mediates cold and drought stress tolerance. J. Plant Physiol. 2015, 185, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Suprun, A.R.; Aleynova, O.A.; Ogneva, Z.V.; Kalachev, A.V.; Dubrovina, A.S. External dsRNA Downregulates Anthocyanin Biosynthesis-Related Genes and Affects Anthocyanin Accumulation in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 6749. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, A.; Giancaspro, A.; Cardone, M.F.; Blanco, A. Real-time PCR for the detection of precise transgene copy number in durum wheat. Cell. Mol. Biol. Lett. 2011, 16, 652–668. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Aleynova, O.A.; Grigorchuk, V.P.; Dubrovina, A.S. Stilbene accumulation and expression of stilbene biosynthesis pathway genes in wild grapevine Vitis amurensis Rupr. Planta 2017, 245, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.; Dubrovina, A.; Tyunin, A. The methylation status of plant genomic DNA influences PCR efficiency. J. Plant Physiol. 2015, 175, 59–67. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Ogneva, Z.V.; Suprun, A.R.; Grigorchuk, V.P.; Dubrovina, A.S. Action of ultraviolet-C radiation and p-coumaric acid on stilbene accumulation and expression of stilbene biosynthesis-related genes in the grapevine Vitis amurensis Rupr. Acta Physiol. Plant. 2019, 41, 28. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Shumakova, O.A.; Manyakhin, A.Y. Effects of the Calmodulin Antagonist W7 on Resveratrol Biosynthesis in Vitis amurensis Rupr. Plant Mol. Biol. Rep. 2013, 31, 1569–1575. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Suprun, A.R.; Ananev, A.A.; Nityagovsky, N.N.; Ogneva, Z.V.; Dubrovina, A.S.; Kiselev, K.V. Effect of cal-modulin-like gene (CML) overexpression on stilbene biosynthesis in cell cultures of Vitis amurensis Rupr. Plants 2022, 11, 171. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RE Name | RE Registration Number in the NSITE-PL Database | Gene | RE Binding Factors |
|---|---|---|---|
| Gap box 2 | 1027, RSP01020 | GapB | GAPF |
| EIN3 BS2 | 2013, RSP01979 | EBF2 | EIN3 |
| E-box | 2473, RSP02439 | FT | CIB1; CIB2, CIB3, CIB4, CIB5 |
| E-box | 2475, RSP02441 | FT | CIB2; CIB4; CIB5 |
| E-box | 2645, RSP02611 | At3g14205 | bHLH122 |
| E-box | 2647, RSP02613 | ERF6 (At4g17490) | bHLH122 |
| E-box | 2649, RSP02615 | ERF6 (At4g17490) | bHLH122 |
| ANAC089 BS 2 | 2664, RSP02630 | sAPX | ANAC089 |
| Cell Line | Overexpressed STS Gene | Fresh Weight, g/L | Dry Weight, g/L | Total Stilbene Content, mg/g DW | Total Stilbene Production, mg/L |
|---|---|---|---|---|---|
| KA0 | - | 222.1 ± 12.2 b | 9.77 ± 0.84 a | 1.24 ± 0.17 c | 12.1 ± 2.6 d |
| 11d-1 | VaSTS11d | 260.1 ± 14.7 a | 10.98 ± 0.77 a | 3.71 ± 1.07 b | 40.7 ± 7.5 b |
| 11d-2 | 262.2 ± 15.1 a | 10.81 ± 0.92 a | 8.49 ± 1.25 a | 91.8 ± 8.9 a | |
| 11d-3 | 232.4 ± 13.3 ab | 10.64 ± 0.65 a | 7.26 ± 1.40 ab | 77.3 ± 7.7 a | |
| 11c-1 | VaSTS11c | 253.6 ± 15.2 ab | 9.81 ± 0.74 a | 2.28 ± 1.02 bc | 22.4 ± 5.0 cd |
| 11c-2 | 247.4 ± 15.5 ab | 9.88 ± 0.98 a | 3.21 ± 0.38 b | 31.7 ± 5.3 bc | |
| 11c-3 | 238.7 ± 16.6 ab | 9.84 ± 0.75 a | 2.66 ± 0.37 b | 26.2 ± 6.1 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiselev, K.V.; Ogneva, Z.V.; Aleynova, O.A.; Suprun, A.R.; Ananev, A.A.; Nityagovsky, N.N.; Dubrovina, A.S. Influence of the 135 bp Intron on Stilbene Synthase VaSTS11 Transgene Expression in Cell Cultures of Grapevine and Different Plant Generations of Arabidopsis thaliana. Horticulturae 2023, 9, 513. https://doi.org/10.3390/horticulturae9040513
Kiselev KV, Ogneva ZV, Aleynova OA, Suprun AR, Ananev AA, Nityagovsky NN, Dubrovina AS. Influence of the 135 bp Intron on Stilbene Synthase VaSTS11 Transgene Expression in Cell Cultures of Grapevine and Different Plant Generations of Arabidopsis thaliana. Horticulturae. 2023; 9(4):513. https://doi.org/10.3390/horticulturae9040513
Chicago/Turabian StyleKiselev, Konstantin V., Zlata V. Ogneva, Olga A. Aleynova, Andrey R. Suprun, Alexey A. Ananev, Nikolay N. Nityagovsky, and Alexandra S. Dubrovina. 2023. "Influence of the 135 bp Intron on Stilbene Synthase VaSTS11 Transgene Expression in Cell Cultures of Grapevine and Different Plant Generations of Arabidopsis thaliana" Horticulturae 9, no. 4: 513. https://doi.org/10.3390/horticulturae9040513