
Transcriptomic Analysis Reveals the Mechanism of Lignin Biosynthesis in Fresh-Cut Cucumber


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Treatments and Storage Conditions
2.2. Measurement of Enzyme Activity
2.3. Determination of Lignin Contents
2.4. Transcriptomic Analysis
2.4.1. RNA Extraction
2.4.2. Construction of Library
2.4.3. Sequencing Method
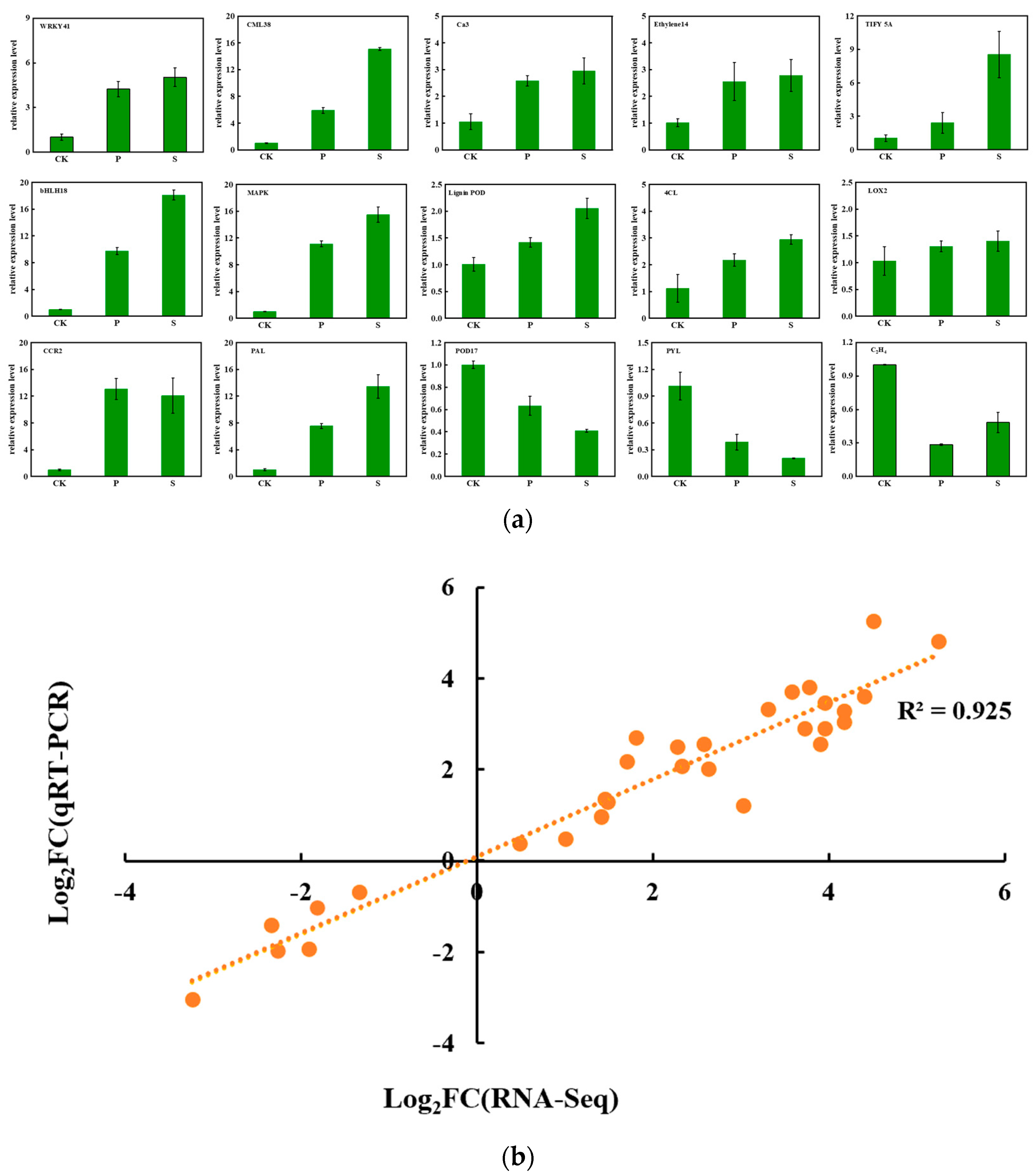
2.4.4. Verification of DEGs by qRT-PCR
2.5. Statistical Analysis
3. Results
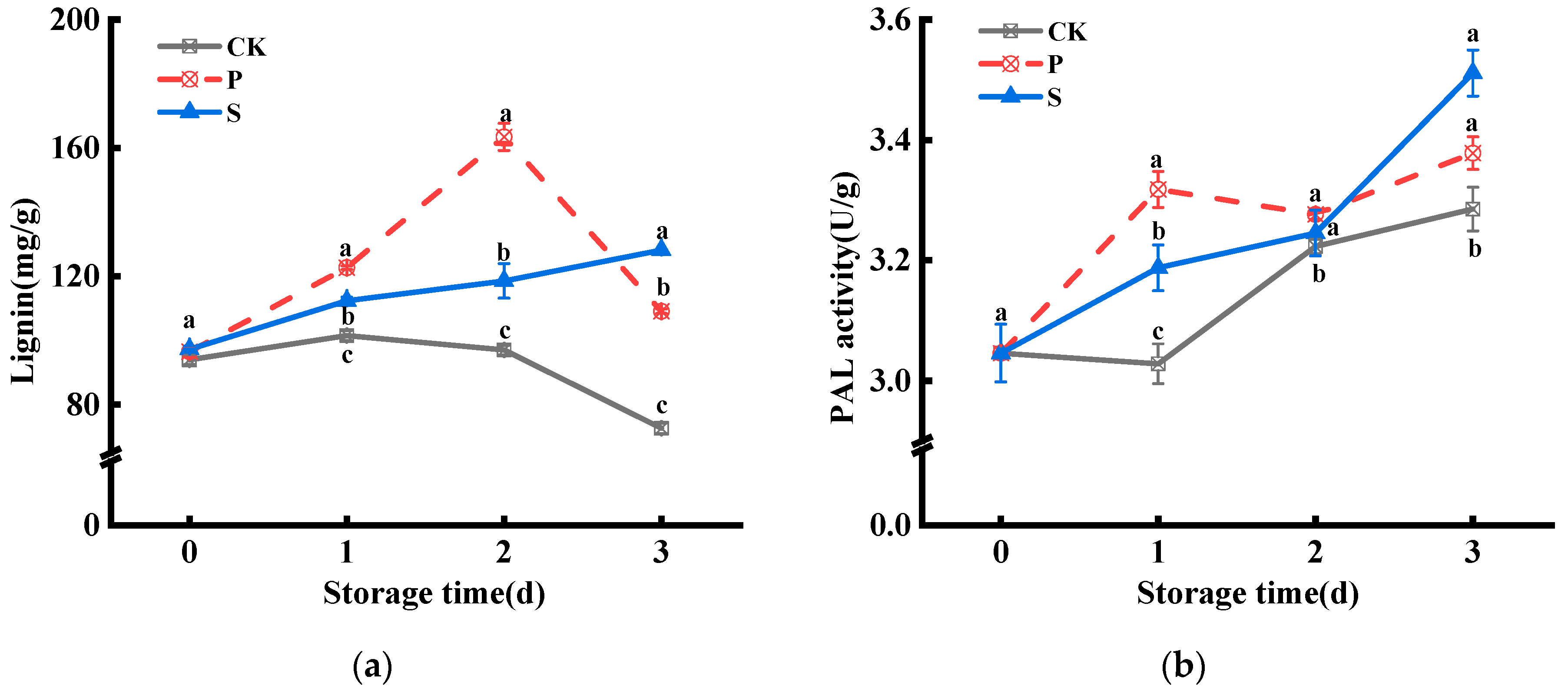
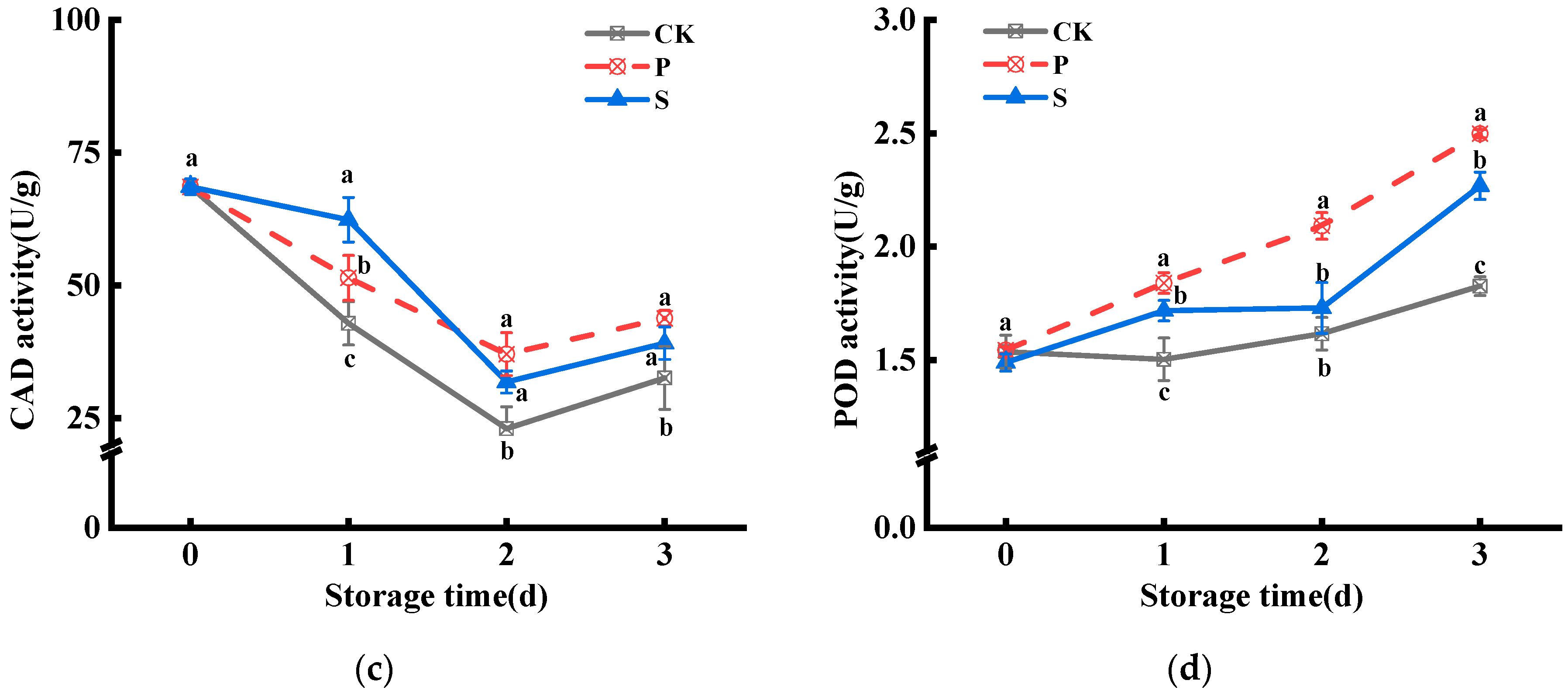
3.1. Lignin Contents and Enzyme Activity of PAL, CAD, and POD
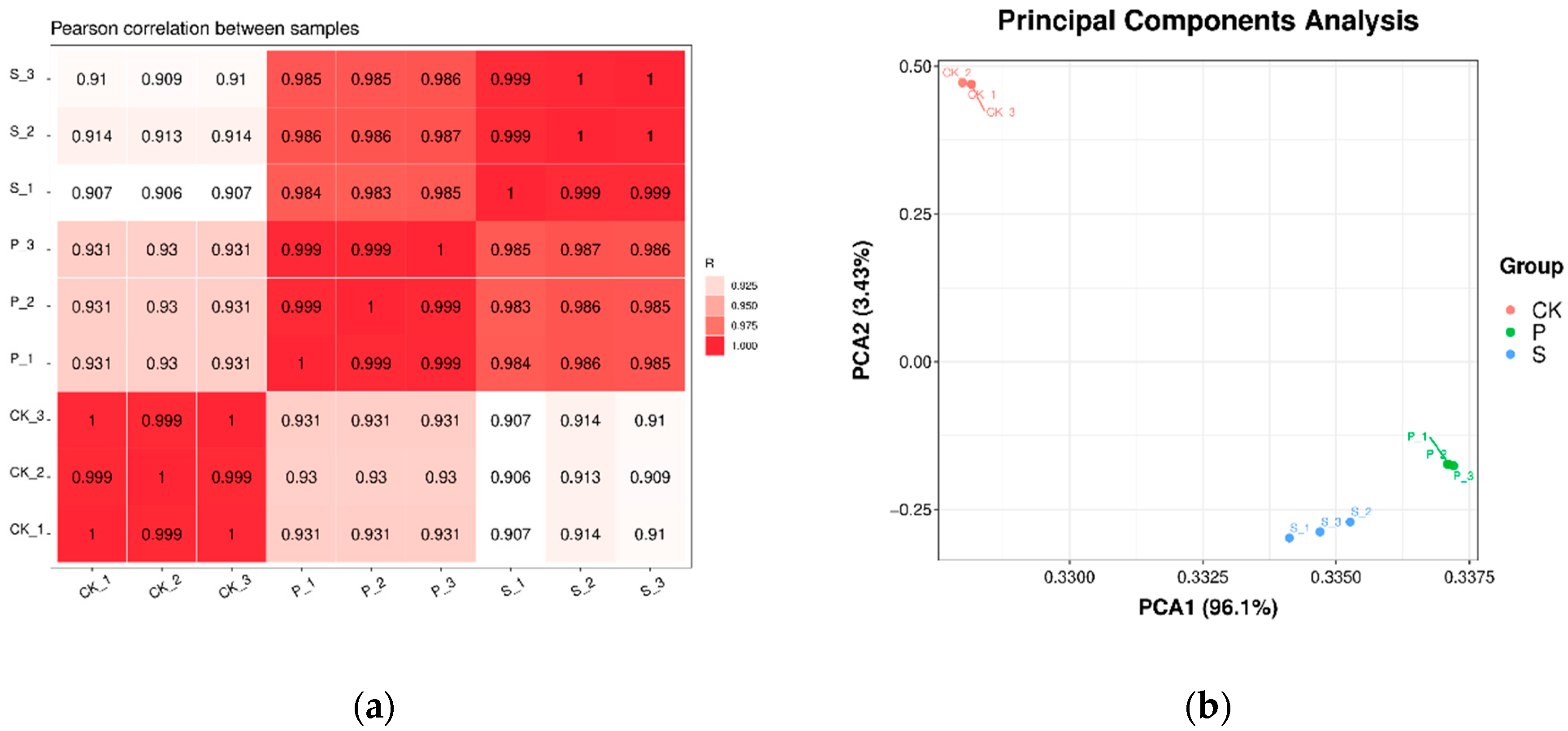
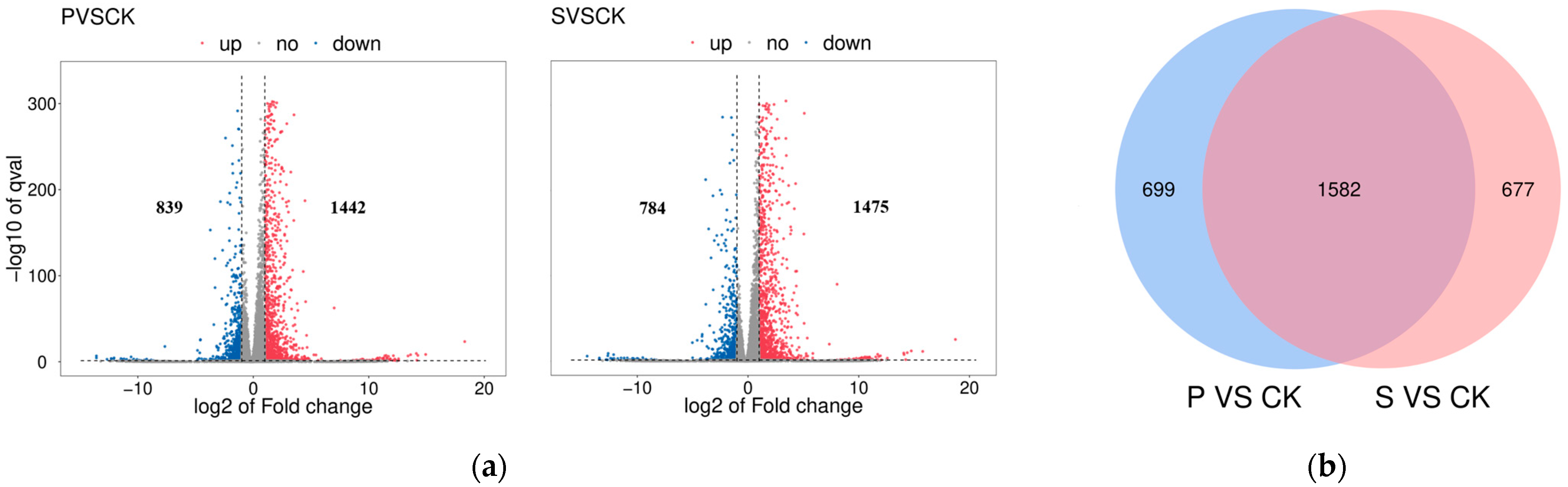
3.2. Transcriptomic Analysis
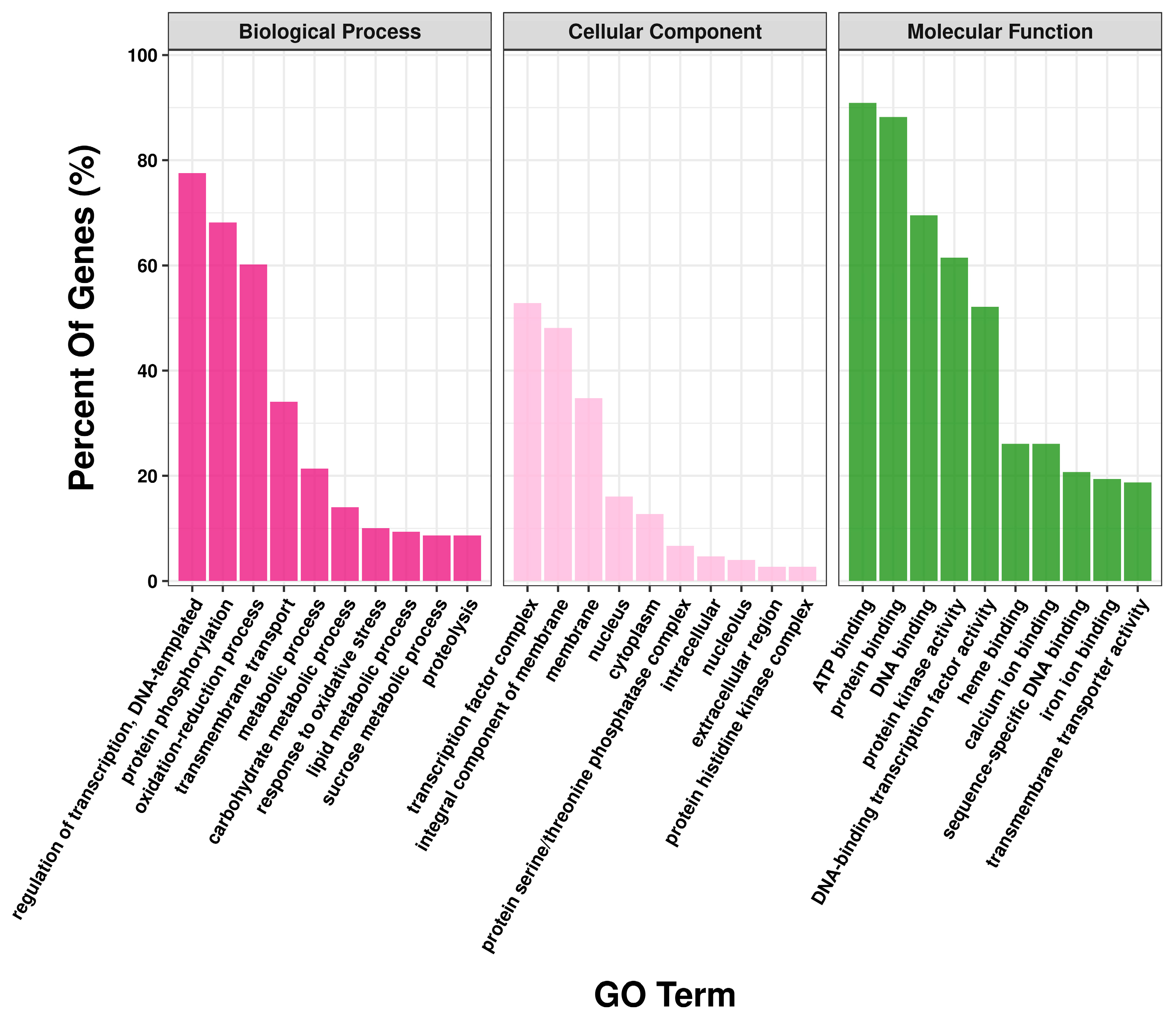
3.3. GO Annotation of Differentially Expressed Genes
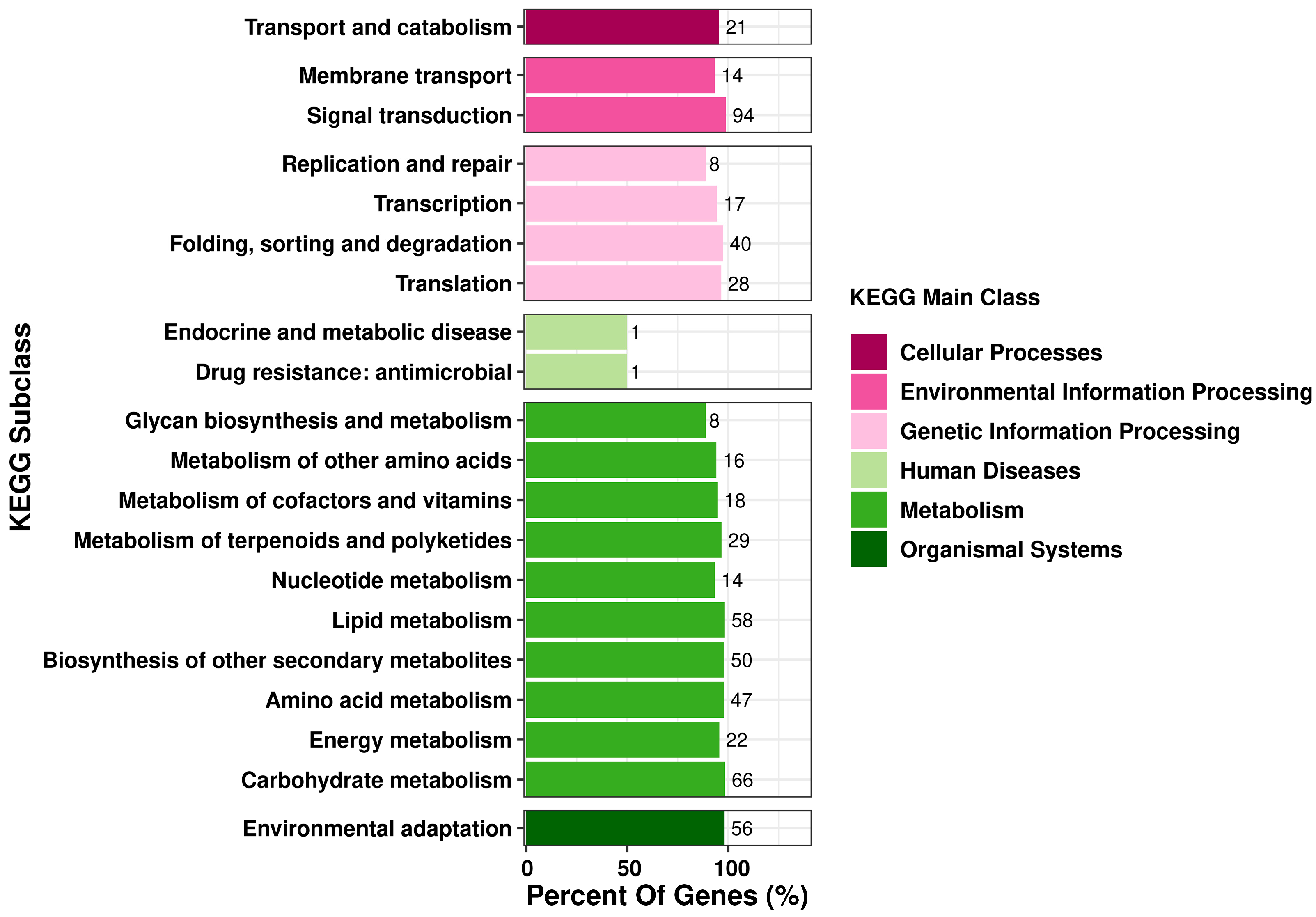
3.4. KEGG Pathway Analysis
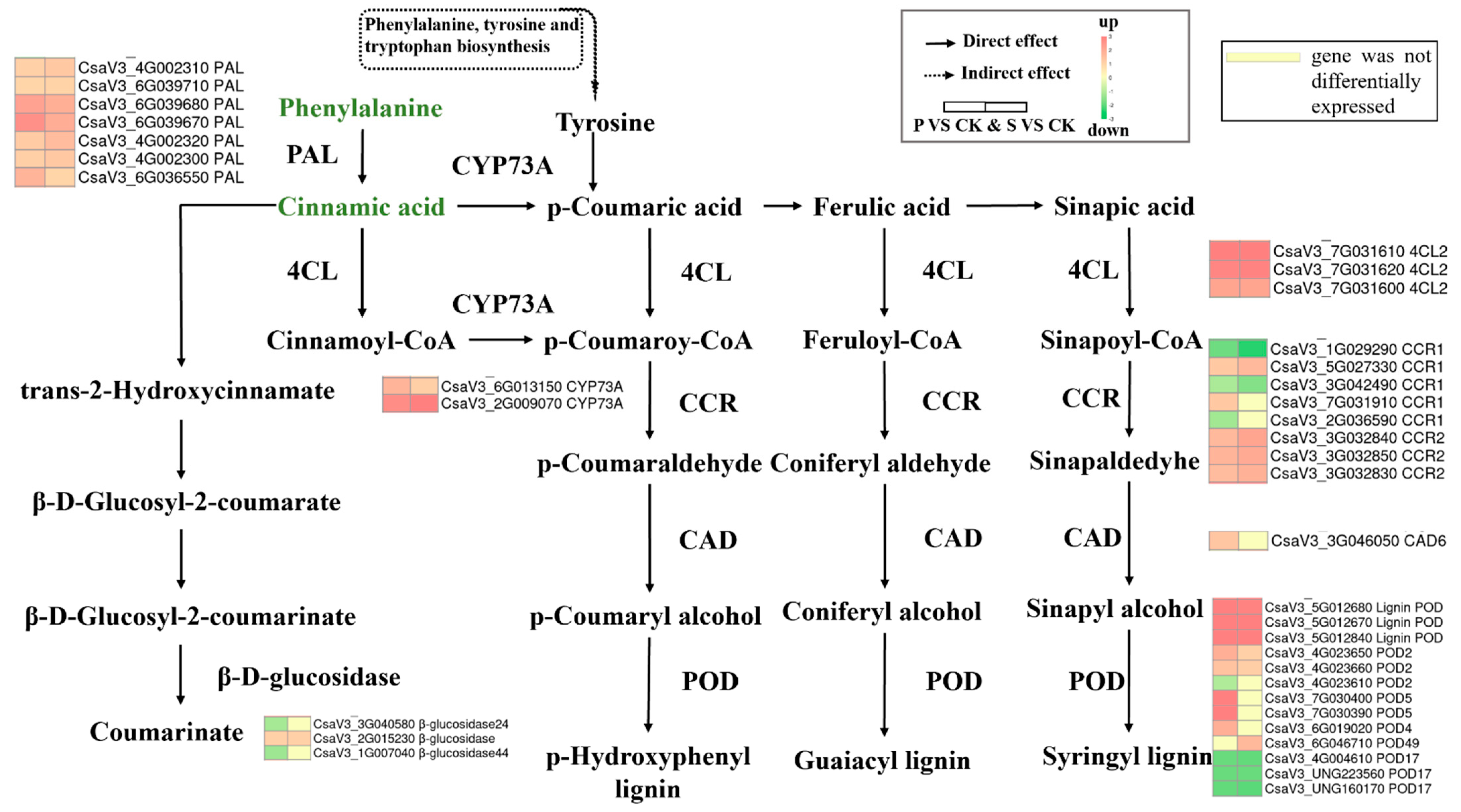
3.5. Critical Metabolic Pathways Induced in Response to Mechanical Damages in Cucumbers
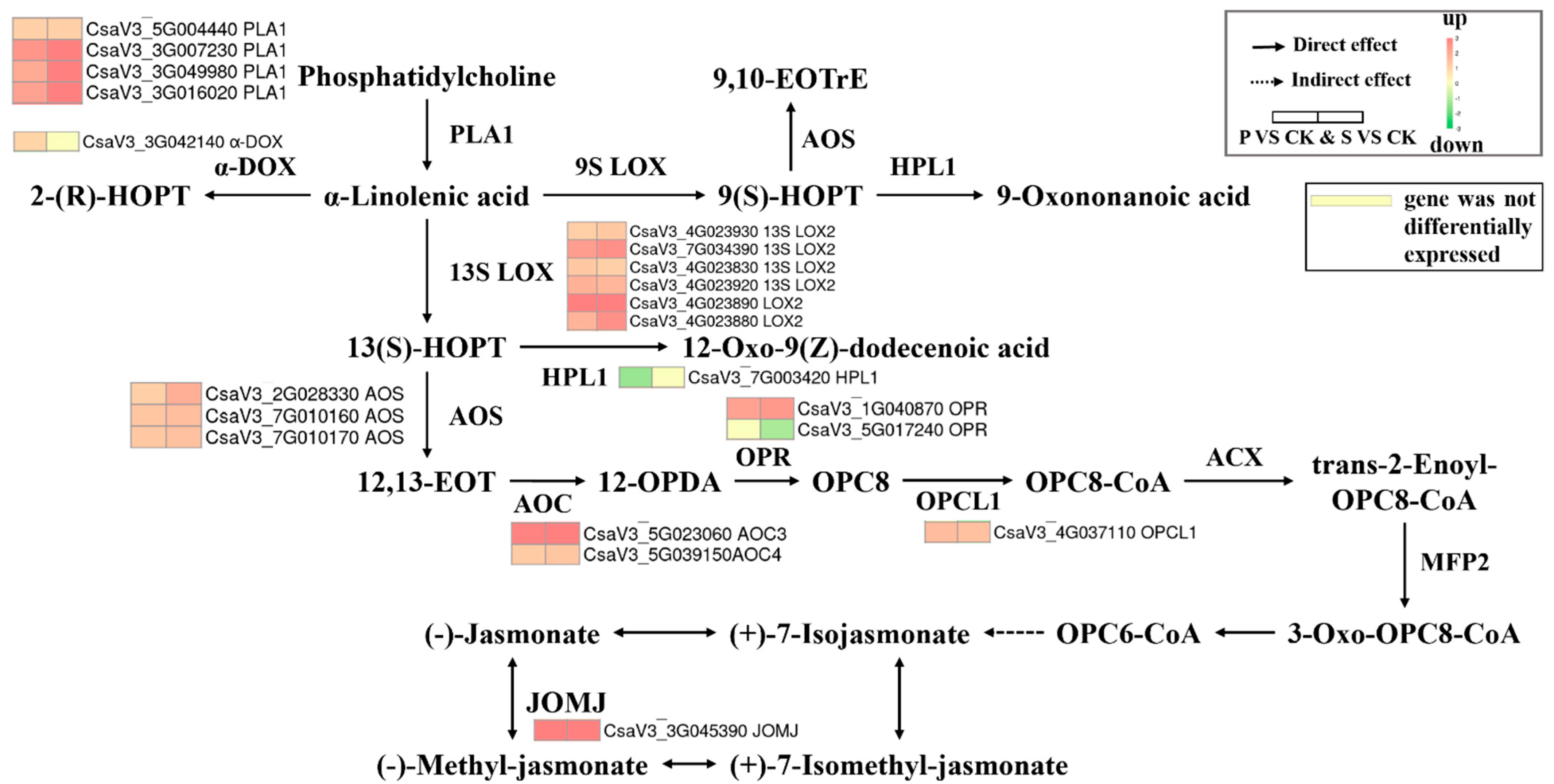
3.5.1. Lipid Metabolism
3.5.2. Amino Acid Metabolism
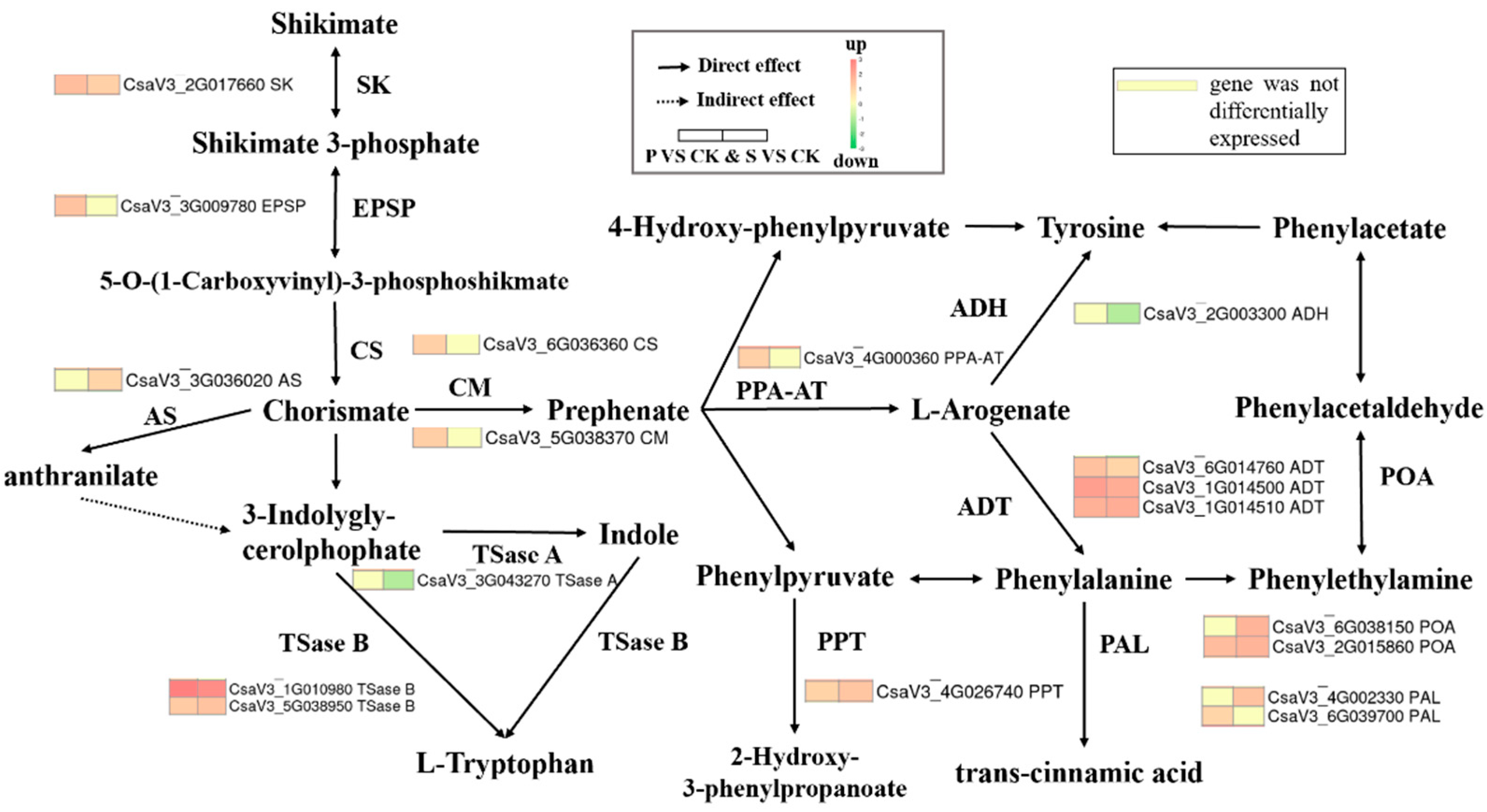
3.5.3. Phenylpropanoid Biosynthesis
3.6. Verification of DEGs by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Valverde-Miranda, D.; Díaz-Pérez, M.; Gómez-Galán, M.; Callejón-Ferre, Á.-J. Total soluble solids and dry matter of cucumber as indicators of shelf life. Postharvest Biol. Technol. 2021, 180, 111603. [Google Scholar] [CrossRef]
- Patel, C.; Panigrahi, J. Starch glucose coating-induced postharvest shelf-life extension of cucumber. Food Chem. 2019, 288, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Zhang, M.; Chen, H. Effect of Ultrasound Treatment Combined with Carbon Dots Coating on the Microbial and Physicochemical Quality of Fresh-Cut Cucumber. Food Bioprocess Technol. 2020, 13, 648–660. [Google Scholar] [CrossRef]
- Olawuyi, I.F.; Park, J.J.; Lee, J.J.; Lee, W.Y. Combined effect of chitosan coating and modified atmosphere packaging on fresh-cut cucumber. Food Sci. Nutr. 2019, 7, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, H.; Rizwan, H.M.; Wang, N.; Jiang, J.; She, W.; Zheng, G.; Pan, H.; Guo, Z.; Pan, D.; et al. Transcriptome Analysis Reveals Candidate Lignin-Related Genes and Transcription Factors during Fruit Development in Pomelo (Citrus maxima). Genes 2022, 13, 845. [Google Scholar] [CrossRef]
- Wu, J.-L.; Pan, T.-F.; Guo, Z.-X.; Pan, D.-M. Specific Lignin Accumulation in Granulated Juice Sacs of Citrus maxima. J. Agric. Food Chem. 2014, 62, 12082–12089. [Google Scholar] [CrossRef]
- Chauhan, O.P.; Raju, P.S.; Ravi, N.; Singh, A.; Bawa, A.S. Effectiveness of ozone in combination with controlled atmosphere on quality characteristics including lignification of carrot sticks. J. Food Eng. 2011, 102, 43–48. [Google Scholar] [CrossRef]
- Cai, C.; Xu, C.; Li, X.; Ferguson, I.; Chen, K. Accumulation of lignin in relation to change in activities of lignification enzymes in loquat fruit flesh after harvest. Postharvest Biol. Technol. 2005, 40, 163–169. [Google Scholar] [CrossRef]
- Hou, D.; Lu, H.; Zhao, Z.; Pei, J.; Yang, H.; Wu, A.; Yu, X.; Lin, X. Integrative transcriptomic and metabolomic data provide insights into gene networks associated with lignification in postharvest Lei bamboo shoots under low temperature. Food Chem. 2022, 368, 130822. [Google Scholar] [CrossRef]
- Veronico, P.; Paciolla, C.; Pomar, F.; De Leonardis, S.; García-Ulloa, A.; Melillo, M.T. Changes in lignin biosynthesis and monomer composition in response to benzothiadiazole and root-knot nematode Meloidogyne incognita infection in tomato. J. Plant Physiol. 2018, 230, 40–50. [Google Scholar] [CrossRef]
- Guo, Y.; Li, M.; Han, H.; Cai, J. Salmonella enterica serovar Choleraesuis on fresh-cut cucumber slices after reduction treatments. Food Control 2016, 70, 20–25. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, Y.; Guan, H.; Liang, H.; Zhao, X.; Wang, D. Adhesion mechanism and biofilm formation of Escherichia coli O157:H7 in infected cucumber (Cucumis sativus L.). Food Microbiol. 2022, 105, 103885. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, C.; Jiang, A.; Zhao, Q.; Zhang, Y.; Hu, W. miRNA and Degradome Sequencing Identify miRNAs and Their Target Genes Involved in the Browning Inhibition of Fresh-Cut Apples by Hydrogen Sulfide. J. Agric. Food Chem. 2020, 68, 8462–8470. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Cao, Y.; Jiang, C.Z.; Li, G.; Liu, P.; Liu, S.; Wang, Q. Cysteine Protease Inhibitors Reduce Enzymatic Browning of Potato by Lowering the Accumulation of Free Amino Acids. J. Agric. Food Chem. 2020, 68, 2467–2476. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Jin, P.; Li, M.; Wang, L.; Zheng, Y. Physiological and Transcriptomic Analysis Validates Previous Findings of Changes in Primary Metabolism for the Production of Phenolic Antioxidants in Wounded Carrots. J. Agric. Food Chem. 2017, 65, 7159–7167. [Google Scholar] [CrossRef] [PubMed]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Biosynthesis of phenolic antioxidants in carrot tissue increases with wounding intensity. Food Chem. 2012, 134, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Ji, Y.; Yang, X.; Pang, L.; Cheng, J.; Lu, X.; Zheng, J.; Yin, L.; Hu, W. Antioxidant activity and microbial safety of fresh-cut red cabbage stored in different packaging films. LWT 2023, 175, 114478. [Google Scholar] [CrossRef]
- Au-Foster, C.E.; Au-Martin, T.M.; Au-Pauly, M. Comprehensive Compositional Analysis of Plant Cell Walls (Lignocellulosic biomass) Part II: Carbohydrates. JoVE 2010, 12, e1837. [Google Scholar]
- Barnes, W.J.; Anderson, C.T. Acetyl Bromide Soluble Lignin (ABSL) Assay for Total Lignin Quantification from Plant Biomass. Bio-Protocol 2017, 7, e2149. [Google Scholar] [CrossRef]
- Fugate, K.K.; Finger, F.L.; Lafta, A.M.; Dogramaci, M.; Khan, M.F.R. Wounding rapidly alters transcription factor expression, hormonal signaling, and phenolic compound metabolism in harvested sugarbeet roots. Front. Plant Sci. 2022, 13, 1070247. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Xu, Y.; Sarengaowa; Ji, Y.; Yang, X.; Feng, K. Proteomic analysis validates previous findings on wounding-responsive plant hormone signaling and primary metabolism contributing to the biosynthesis of secondary metabolites based on metabolomic analysis in harvested broccoli (Brassica oleracea L. var. italica). Food Res. Int. 2021, 145, 110388. [Google Scholar] [PubMed]
- Cheong, J.J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. TIG 2003, 19, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Onohata, T.; Gomi, K. Overexpression of jasmonate-responsive OsbHLH034 in rice results in the induction of bacterial blight resistance via an increase in lignin biosynthesis. Plant Cell Rep. 2020, 39, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Salazar, R.; Pollmann, S.; Morales-Quintana, L.; Herrera, R.; Caparrós-Ruiz, D.; Ramos, P. In seedlings of Pinus radiata, jasmonic acid and auxin are differentially distributed on opposite sides of tilted stems affecting lignin monomer biosynthesis and composition. Plant Physiol. Biochem. 2019, 135, 215–223. [Google Scholar] [CrossRef]
- Abreu, I.N.; Johansson, A.I.; Sokołowska, K.; Niittylä, T.; Sundberg, B.; Hvidsten, T.R.; Street, N.R.; Moritz, T. A metabolite roadmap of the wood-forming tissue in Populus tremula. New Phytol. 2020, 228, 1559–1572. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Lafuente, M.T.; Forment, J.; Gadea, J.; De Vos, R.C.H.; Bovy, A.G.; GonzÁLez-Candelas, L. Transcriptomic profiling of citrus fruit peel tissues reveals fundamental effects of phenylpropanoids and ethylene on induced resistance. Mol. Plant Pathol. 2011, 12, 879–897. [Google Scholar] [CrossRef]
- Ding, Z.; Fu, L.; Tie, W.; Yan, Y.; Wu, C.; Dai, J.; Zhang, J.; Hu, W. Highly dynamic, coordinated, and stage-specific profiles are revealed by a multi-omics integrative analysis during tuberous root development in cassava. J. Exp. Bot. 2020, 71, 7003–7017. [Google Scholar] [CrossRef]
- Li, X.; Li, B.; Min, D.; Ji, N.; Zhang, X.; Li, F.; Zheng, Y. Transcriptomic analysis reveals key genes associated with the biosynthesis regulation of phenolics in fresh-cut pitaya fruit (Hylocereus undatus). Postharvest Biol. Technol. 2021, 181, 111684. [Google Scholar] [CrossRef]
- Woolfson, K.N.; Zhurov, V.; Wu, T.; Kaberi, K.M.; Wu, S.; Bernards, M.A. Transcriptomic analysis of wound-healing in Solanum tuberosum (potato) tubers: Evidence for a stepwise induction of suberin-associated genes. Phytochemistry 2023, 206, 113529. [Google Scholar] [CrossRef]
- Kong, J.-Q. Phenylalanine ammonia-lyase, a key component used for phenylpropanoids production by metabolic engineering. RSC Adv. 2015, 5, 62587–62603. [Google Scholar] [CrossRef]
- Dong, N.-Q.; Lin, H.-X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Postharvest UV-C irradiation increased the flavonoids and anthocyanins accumulation, phenylpropanoid pathway gene expression, and antioxidant activity in sweet cherries (Prunus avium L.). Postharvest Biol. Technol. 2021, 175, 111490. [Google Scholar] [CrossRef]
- Bunsiri, A.; Paull, R.E.; Ketsa, S. Increased activities of phenyalanine ammonia lyase, peroxidase, and cinnamyl alcohol dehydrogenase in relation to pericarp hardening after physical impact in mangosteen (Garcinia mangostana L.). J. Hortic. Sci. Biotechnol. 2012, 87, 231–236. [Google Scholar] [CrossRef]
- Wei, x.; Guan, w.; Yang, y.; Shao, y.; Mao, l. Methyl jasmonate promotes wound healing by activation of phenylpropanoid metabolism in harvested kiwifruit. Postharvest Biol. Technol. 2021, 175, 111472. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Xu, Y.; Yang, X.; Ji, Y.; Feng, K.; Sarengaowa. Metabolomics and physiological analyses validates previous findings on the mechanism of response to wounding stress of different intensities in broccoli. Food Res. Int. 2021, 140, 110058. [Google Scholar] [CrossRef]
- Song, Z.; Li, F.; Guan, H.; Xu, Y.; Fu, Q.; Li, D. Combination of nisin and ε-polylysine with chitosan coating inhibits the white blush of fresh-cut carrots. Food Control 2017, 74, 34–44. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, B.; Ma, L.; Zheng, X.; Gong, D.; Xue, H.; Bi, Y.; Wang, Y.; Zhang, Z.; Prusky, D. Benzo-(1, 2, 3)-thiadiazole-7-carbothioic acid s-methyl ester (BTH) promotes tuber wound healing of potato by elevation of phenylpropanoid metabolism. Postharvest Biol. Technol. 2019, 153, 125–132. [Google Scholar] [CrossRef]
- Li, H.; Suo, J.; Han, Y.; Liang, C.; Jin, M.; Zhang, Z.; Rao, J. The effect of 1-methylcyclopropene, methyl jasmonate and methyl salicylate on lignin accumulation and gene expression in postharvest ‘Xuxiang’ kiwifruit during cold storage. Postharvest Biol. Technol. 2017, 124, 107–118. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Data | Valid Data | Valid Ratio (%) | Q20% | Q30% | GC% | ||
|---|---|---|---|---|---|---|---|---|
| Read | Base | Read | Base | |||||
| CK | 40,771,589 | 6.11 G | 37,754,237 | 5.68 G | 93.02 | 99.96 | 96.80 | 42.50 |
| P | 39,870,298 | 5.98 G | 39,146,899 | 5.54 G | 93.11 | 99.96 | 96.74 | 42.50 |
| S | 41,079,656 | 6.03 G | 38,250,758 | 5.73 G | 93.11 | 99.96 | 96.67 | 42.50 |
| Lipid Metabolism | P vs. CK | S vs. CK | ||
|---|---|---|---|---|
| DEGs | p | DEGs | p | |
| Alpha-linolenic acid metabolism | 20 | 0.0001 | 20 | 0.0001 |
| Fatty acid elongation | 5 | 0.2169 | 7 | 0.0350 |
| Linoleic acid metabolism | 10 | 0.0023 | 7 | 0.0615 |
| Ether lipid metabolism | 7 | 0.1773 | 8 | 0.0821 |
| Secondary bile acid biosynthesis | 1 | 0.1918 | 1 | 0.1894 |
| Glycerophospholipid metabolism | 14 | 0.2693 | 14 | 0.2536 |
| Arachidonic acid metabolism | 5 | 0.1801 | 4 | 0.3503 |
| Glycerolipid metabolism | 13 | 0.3768 | 13 | 0.3589 |
| Biosynthesis of unsaturated fatty acids | 4 | 0.7434 | 5 | 0.5479 |
| Cutin, suberine, and wax biosynthesis | 7 | 0.1255 | 4 | 0.6144 |
| Sphingolipid metabolism | 9 | 0.4929 | 8 | 0.6222 |
| Steroid biosynthesis | 1 | 0.9634 | 2 | 0.8297 |
| Fatty acid degradation | 3 | 0.9573 | 4 | 0.8786 |
| Fatty acid biosynthesis | 1 | 0.9961 | 1 | 0.9958 |
| Amino Acid Metabolism | P vs. CK | S vs. CK | ||
|---|---|---|---|---|
| DEGs | p | DEGs | p | |
| Phenylalanine metabolism | 16 | 0.0028 | 16 | 0.0025 |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | 11 | 0.0098 | 10 | 0.0236 |
| Glycine, serine, and threonine metabolism | 13 | 0.1175 | 15 | 0.0316 |
| Alanine, aspartate and glutamate metabolism | 7 | 0.1186 | 6 | 0.5433 |
| Arginine biosynthesis | 7 | 0.1773 | 4 | 0.6864 |
| Valine, leucine, and isoleucine biosynthesis | 2 | 0.7133 | 2 | 0.7063 |
| Lysine biosynthesis | 1 | 0.7497 | 1 | 0.7450 |
| Tryptophan metabolism | 3 | 0.9007 | 4 | 0.7623 |
| Tyrosine metabolism | 4 | 0.797 | 4 | 0.7884 |
| Cysteine and methionine metabolism | 11 | 0.7487 | 10 | 0.8269 |
| Histidine metabolism | - | - | 1 | 0.8493 |
| Valine, leucine, and isoleucine degradation | - | - | 3 | 0.9717 |
| Arginine and proline metabolism | 3 | 0.0423 | 3 | 0.9777 |
| Lysine degradation | - | - | 1 | 0.9852 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Yuan, N.; Guan, Y.; Chen, C.; Hu, W. Transcriptomic Analysis Reveals the Mechanism of Lignin Biosynthesis in Fresh-Cut Cucumber. Horticulturae 2023, 9, 500. https://doi.org/10.3390/horticulturae9040500
Wang Y, Yuan N, Guan Y, Chen C, Hu W. Transcriptomic Analysis Reveals the Mechanism of Lignin Biosynthesis in Fresh-Cut Cucumber. Horticulturae. 2023; 9(4):500. https://doi.org/10.3390/horticulturae9040500
Chicago/Turabian StyleWang, Yi, Ning Yuan, Yuge Guan, Chen Chen, and Wenzhong Hu. 2023. "Transcriptomic Analysis Reveals the Mechanism of Lignin Biosynthesis in Fresh-Cut Cucumber" Horticulturae 9, no. 4: 500. https://doi.org/10.3390/horticulturae9040500