

Effects of Light-Emitting Diodes (LEDs) on Growth, Nitrates and Osmoprotectant Content in Microgreens of Aromatic and Medicinal Plants
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
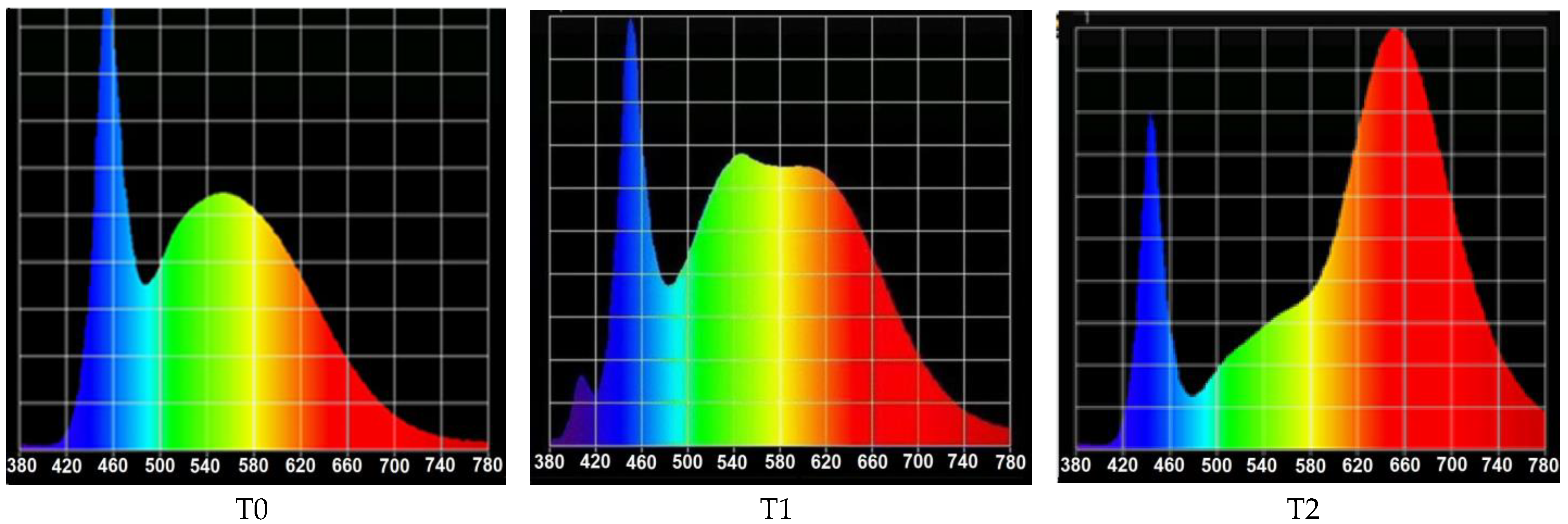
2.1. Light Devices
2.2. Plant Materials
2.2.1. Fresh Weight and Height
2.2.2. Determination of Nitrate Content
2.2.3. Chlorophyll and Carotene Determination
2.2.4. Glucose and Proline Determination
2.2.5. Soluble-Protein Determination
2.2.6. Statistical Analyses
3. Results
3.1. Fresh Weight (FW) and Height (H)
3.2. Nitrate Content
3.3. Chlorophyll and Carotenoid Contents
3.4. Glucose, Proline, and Proteins Contents
4. Discussion
4.1. Fresh Weight (FW) and Height (H)
4.2. Nitrate Content
4.3. Chlorophyll and Carotenoid Contents
4.4. Glucose, Proline, and Proteins Contents
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional Value, Bioactive Compounds and Health Benefits of Lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Luo, Y.; Wang, Q. Assessment of Vitamin and Carotenoid Concentrations of Emerging Food Products: Edible Microgreens. J. Agric. Food Chem. 2012, 60, 7644–7651. [Google Scholar] [CrossRef]
- Ghoora, M.D.; Babu, D.R.; Srividya, N. Nutrient Composition, Oxalate Content and Nutritional Ranking of Ten Culinary Microgreens. J. Food Compos. Anal. 2020, 91, 103495. [Google Scholar] [CrossRef]
- Pinto, E.; Almeida, A.A.; Aguiar, A.A.; Ferreira, I.M.P.L.V.O. Comparison between the Mineral Profile and Nitrate Content of Microgreens and Mature Lettuces. J. Food Compos. Anal. 2015, 37, 38–43. [Google Scholar] [CrossRef]
- de la Fuente, B.; López-García, G.; Máñez, V.; Alegría, A.; Barberá, R.; Cilla, A. Evaluation of the Bioaccessibility of Antioxidant Bioactive Compounds and Minerals of Four Genotypes of Brassicaceae Microgreens. Foods 2019, 8, 250. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-Scale Vegetable Production and the Rise of Microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Azizi, M. Study of Four Improved Cultivars of Matricaria chamomilla L. in Climatic Condition of Iran. Iran. J. Med. Aromat. Plants Res. 2007, 22, 386–396. [Google Scholar]
- Sangwan, R.S.; Chaurasiya, N.D.; Lal, P.; Misra, L.; Tuli, R.; Sangwan, N.S. Withanolide A Is Inherently de Novo Biosynthesized in Roots of the Medicinal Plant Ashwagandha (Withania Somnifera). Physiol. Plant. 2008, 133, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Umano, K.; Shibamoto, T.; Lee, K.-G. Identification of Volatile Components in Basil (Ocimum basilicum L.) and Thyme Leaves (Thymus vulgaris L.) and Their Antioxidant Properties. Food Chem. 2005, 91, 131–137. [Google Scholar] [CrossRef]
- Zandi, P.; Basu, S.K.; Khatibani, L.B.; Balogun, M.O.; Aremu, M.O.; Sharma, M.; Kumar, A.; Sengupta, R.; Li, X.; Li, Y.; et al. Fenugreek (Trigonella foenum-graecum L.) Seed: A Review of Physiological and Biochemical Properties and Their Genetic Improvement. Acta Physiol. Plant. 2014, 37, 1714. [Google Scholar] [CrossRef]
- Kaur, V.; Kaur, R.; Bhardwaj, U. A Review on Dill Essential Oil and Its Chief Compounds as Natural Biocide. Flavour Fragr. J. 2021, 36, 412–431. [Google Scholar] [CrossRef]
- Hendawy, S.F.; Hussein, M.S.; El-Gohary, A.E.; Soliman, W.S. Chemical Constituents of Essential Oil in Chervil (Anthriscus cerefolium L. Hoffm.) Cultivated in Different Locations. J. Essent. Oil Bear. Plants 2019, 22, 264–272. [Google Scholar] [CrossRef]
- Delian, E.; Chira, A.; Bădulescu, L.; Chira, L. Insights into Microgreens Physiology. Sci. Pap.–Ser. B Hortic. 2015, 59, 447–454. [Google Scholar]
- Avercheva, O.V.; Berkovich, Y.A.; Erokhin, A.N.; Zhigalova, T.V.; Pogosyan, S.I.; Smolyanina, S.O. Growth and Photosynthesis of Chinese Cabbage Plants Grown under Light-Emitting Diode-Based Light Source. Russ. J. Plant Physiol. 2009, 56, 14–21. [Google Scholar] [CrossRef]
- Zhang, X.; Bian, Z.; Yuan, X.; Chen, X.; Lu, C. A Review on the Effects of Light-Emitting Diode (LED) Light on the Nutrients of Sprouts and Microgreens. Trends Food Sci. Technol. 2020, 99, 203–216. [Google Scholar] [CrossRef]
- Morrow, R.C. LED Lighting in Horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef] [Green Version]
- Ying, Q.; Kong, Y.; Jones-Baumgardt, C.; Zheng, Y. Responses of Yield and Appearance Quality of Four Brassicaceae Microgreens to Varied Blue Light Proportion in Red and Blue Light-Emitting Diodes Lighting. Sci. Hortic. 2020, 259, 108857. [Google Scholar] [CrossRef]
- Urrestarazu, M.; Nájera, C.; del Mar Gea, M. Effect of the Spectral Quality and Intensity of Light-Emitting Diodes on Several Horticultural Crops. HortScience 2016, 51, 268–271. [Google Scholar] [CrossRef] [Green Version]
- Sonneveld, C.; Straver, N. Voedingsoplossingen Voor Groenten en Bloemen Geteeld in Water of Substraten = Nutrient Solutions for Vegetables and Flowers Grown in Water or Substrates; No. 8; Proefstation voor Tuinbouw onder Glas: Naaldwijk, The Netherlands, 1994. [Google Scholar]
- Lichtenthaler, H.K. [34] Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Plant Cell Membranes; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water Stress Induced Changes in Concentrations of Proline and Total Soluble Sugars in Nodulated Alfalfa (Medicago sativa) Plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Paquin, R.; Lechasseur, P. Observations Sur Une Méthode de Dosage de La Proline Libre Dans Les Extraits de Plantes. Can. J. Bot. 1979, 57, 1851–1854. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the Role of Red:Blue LED Lights on Resource Use Efficiency and Nutritional Properties of Indoor Grown Sweet Basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrón-Carrillo, F.; Guil-Guerrero, J.L.; González-Fernández, M.J.; Lyashenko, S.; Battafarano, F.; da Cunha-Chiamolera, T.P.L.; Urrestarazu, M. LED Enhances Plant Performance and Both Carotenoids and Nitrates Profiles in Lettuce. Plant Foods Hum. Nutr. 2021, 76, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Nájera, C.; Urrestarazu, M. Effect of the Intensity and Spectral Quality of LED Light on Yield and Nitrate Accumulation in Vegetables. HortScience 2019, 54, 1745–1750. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-L.; Chang, K.-P. The Growth Response of Leaf Lettuce at Different Stages to Multiple Wavelength-Band Light-Emitting Diode Lighting. Sci. Hortic. 2014, 179, 78–84. [Google Scholar] [CrossRef]
- Ouzounis, T.; Heuvelink, E.; Ji, Y.; Schouten, H.J.; Visser, R.G.F.; Marcelis, L.F.M. Blue and Red LED Lighting Effects on Plant Biomass, Stomatal Conductance, and Metabolite Content in Nine Tomato Geno-types. VIII International Symposium on Light in Horticulture 1134. 2016. Available online: https://www.ishs.org/ishs-article/1134_34 (accessed on 1 January 2023).
- Anjana, S.U.; Iqbal, M. Nitrate Accumulation in Plants, Factors Affecting the Process, and Human Health Implications. A Review. Agron. Sustain. Dev. 2007, 27, 45–57. [Google Scholar] [CrossRef]
- Gruda, N. Impact of Environmental Factors on Product Quality of Greenhouse Vegetables for Fresh Consumption. Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Cantliffe, D.J. Nitrate Accumulation in Spinach Grown Under Different Light Intensities1,2. J. Am. Soc. Hortic. Sci. 1972, 97, 152–154. [Google Scholar] [CrossRef]
- Aslam, M.; Oaks, A.; Huffaker, R.C. Effect of Light and Glucose on the Induction of Nitrate Reductase and on the Distribution of Nitrate in Etiolated Barley Leaves 1. Plant Physiol. 1976, 58, 588–591. [Google Scholar] [CrossRef] [Green Version]
- Blom-Zandstra, M.; Lampe, J.E.M.; Ammerlaan, F.H.M. C and N Utilization of Two Lettuce Genotypes during Growth under Non-Varying Light Conditions and after Changing the Light Intensity. Physiol. Plant. 1988, 74, 147–153. [Google Scholar] [CrossRef]
- Blom-Zandstra, M.; Lampe, J.E.M. The Role of Nitrate in the Osmoregulation of Lettuce (Lactuca sativa L.) Grown at Different Light Intensities. J. Exp. Bot. 1985, 36, 1043–1052. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Takase, M.; Kon, N.; Fujiwara, K.; Kurata, K. Effect of Light Quality on Growth and Vegetable Quality in Leaf Lettuce, Spinach and Komatsuna. Environ. Control Biol. 2007, 45, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of Light Quality on CO2 Assimilation, Chlorophyll-Fluorescence Quenching, Expression of Calvin Cycle Genes and Carbohydrate Accumulation in Cucumis Sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef]
- Sood, S.; Gupta, V.; Tripathy, B.C. Photoregulation of the Greening Process of Wheat Seedlings Grown in Red Light*. Plant Mol. Biol. 2005, 59, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.-X.; Xu, Z.-G.; Liu, X.-Y.; Tang, C.-M.; Wang, L.-W.; Han, X. Effects of Light Intensity on the Growth and Leaf Development of Young Tomato Plants Grown under a Combination of Red and Blue Light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Goins, G.D.; Yorio, N.C.; Sanwo, M.M.; Brown, C.S. Photomorphogenesis, Photosynthesis, and Seed Yield of Wheat Plants Grown under Red Light-Emitting Diodes (LEDs) with and without Supplemental Blue Lighting. J. Exp. Bot. 1997, 48, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, B.C.; Brown, C.S. Root-Shoot Interaction in the Greening of Wheat Seedlings Grown under Red Light. Plant Physiol. 1995, 107, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagi, T.; Okamoto, K.; Takita, S. Effects of blue, red, and blue/red lights of two different PPF levels on growth and morphogenesis of lettuce plants. Acta Hortic. 1996, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving Spinach, Radish, and Lettuce Growth under Red Light-Emitting Diodes (LEDs) with Blue Light Supplementation. HortScience 2001, 36, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Yorio, N.C.; Wheeler, R.M.; Goins, G.D.; Sanwo-Lewandowski, M.M.; Mackowiak, C.L.; Brown, C.S.; Sager, J.C.; Stutte, G.W. Blue light requirements for crop plants used in bioregenerative life support systems. Life Support Biosph. Sci. 1998, 5, 119–128. [Google Scholar]
- Li, H.; Xu, Z.; Tang, C. Effect of Light-Emitting Diodes on Growth and Morphogenesis of Upland Cotton (Gossypium hirsutum L.) Plantlets in Vitro. Plant Cell Tiss. Organ. Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of Different Light Sources on the Growth of Non-Heading Chinese Cabbage (Brassica campestris L.). JAS 2012, 4, 262. [Google Scholar] [CrossRef] [Green Version]
- Lekkham, P.; Srilaong, V.; Pongprasert, N.; Kondo, S. Anthocyanin Concentration and Antioxidant Activity in Light-Emitting Diode (LED)-Treated Apples in a Greenhouse Environmental Control System. Fruits 2016, 71, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current Status and Recent Achievements in the Field of Horticulture with the Use of Light-Emitting Diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Viršilė, A.; Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Miliauskienė, J.; Jankauskienė, J.; Novičkovas, A.; Laužikė, K.; Samuolienė, G. The Distinct Impact of Multi-Color LED Light on Nitrate, Amino Acid, Soluble Sugar and Organic Acid Contents in Red and Green Leaf Lettuce Cultivated in Controlled Environment. Food Chem. 2020, 310, 125799. [Google Scholar] [CrossRef]
- Makowski, W.; Tokarz, B.; Banasiuk, R.; Królicka, A.; Dziurka, M.; Wojciechowska, R.; Tokarz, K.M. Is a Blue–Red Light a Good Elicitor of Phenolic Compounds in the Family Droseraceae? A Comparative Study. J. Photochem. Photobiol. B Biol. 2019, 201, 111679. [Google Scholar] [CrossRef]
- Gyugos, M.; Ahres, M.; Gulyás, Z.; Szalai, G.; Darkó, É.; Mednyánszky, Z.; Dey, N.; Kar, R.K.; Simon-Sarkadi, L.; Kocsy, G. Light Spectrum Modifies the Drought-Induced Changes of Glutathione and Free Amino Acid Levels in Wheat. Acta Physiol. Plant. 2021, 43, 90. [Google Scholar] [CrossRef]
- Chen, L.; Wang, H.; Gong, X.; Zeng, Z.; Xue, X.; Hu, Y. Transcriptome Analysis Reveals Effects of Red and Blue Light-Emitting Diodes (LEDs) on the Growth, Chlorophyll Fluorescence and Endogenous Plant Hormones of Potato (Solanum tuberosum L.) Plantlets Cultured in Vitro. J. Integr. Agric. 2021, 20, 2914–2931. [Google Scholar] [CrossRef]


{kind=link}
| Treatment | Lamp LED | PPFD | Illuminance |
|---|---|---|---|
| T0 | L18 T8 Roblan® | 150 ± 7 a | 6447 ± 186 a |
| T1 | L18 NS1 Valoya® | 141 ± 19 b | 5161 ± 229 b |
| T2 | L18 AP67 Valoya® | 107 ± 7 c | 3837 ± 358 c |
| Fresh Weight (g m−2) | Height (cm) | |||||||
|---|---|---|---|---|---|---|---|---|
| Basil | Fenugreek | Dill | Chervil | Basil | Fenugreek | Dill | Chervil | |
| T0 | 231.3 ± 12 byBz | 150.4 ± 29 cC | 107.9 ± 23 bD | 258 ± 05 bA | 4.9 ± 0.4 bB | 4.5 ± 0.7 cB | 5.9 ± 0.5 cA | 5.5 ± 0.5 bA |
| T1 | 390.7 ± 20 aA | 156.9 ± 20 bC | 076.5 ± 08 cD | 237 ± 06 bB | 6.0 ± 0.4 aA | 5.0 ± 0.3 bB | 5.2 ± 0.3 bB | 5.2 ± 0.3 bB |
| T2 | 225 ± 16 bB | 235.4 ± 23 aB | 144.0 ± 20 aC | 613 ± 12 aA | 5.2 ± 0.2 bC | 6.2 ± 0.4 aB | 7.4 ± 0.6 aA | 7.3 ± 1.0 aA |
| Treatment | Nitrate Content (mg 100 g−1 FW) | |||
|---|---|---|---|---|
| Basil | Fenugreek | Dill | Chervil | |
| T0 | 505.7 ± 43.5 ayAz | 407.1 ± 26.9 aB | 360 ± 13.1 aB | 304.3 ± 12.7 aD |
| T1 | 447.1 ± 24.3 bA | 328.6 ± 12.2 bB | 225.7 ± 7.7 bD | 240 ± 8.2 bC |
| T2 | 458.7 ± 11.3 bA | 331.2 ± 13.6 bC | 355.7 ± 45.4 aB | 220 ± 22.0 cD |
| Chlorophyll (mg g−1 FW) | Carotenoid (mg g−1 FW) | |||||||
|---|---|---|---|---|---|---|---|---|
| Basil | Fenugreek | Dill | Chervil | Basil | Fenugreek | Dill | Chervil | |
| T0 | 2.6 ± 0.08 byBz | 3 ± 0.23 bA | 1.1 ± 0.03 bC | 2.5 ± 0.1 bB | 0.023 ± 0.007 cB | 0.029 ± 0.005 cA | 0.015 ± 0.003 cC | 0.016 ± 0.004 cC |
| T1 | 2.8 ± 0.07 aB | 3.6 ± 0.14 aA | 1.3 ± 0.07 aC | 2.6 ± 0.1 abB | 0.035 ± 0.007 bB | 0.046 ± 0.009 bA | 0.028 ± 0.006 bC | 0.031 ± 0.008 bBC |
| T2 | 2.9 ± 0.04 aB | 3.8 ± 0.15 aA | 1.2 ± 0.04 abC | 2.8 ± 0.07 aB | 0.052 ± 0.005 aAB | 0.058 ± 0.007 aA | 0.045 ± 0.01 aC | 0.046 ± 0.008 aBC |
| Glucose (mg g−1 FW) | Proline (mg g−1 FW) | |||||||
|---|---|---|---|---|---|---|---|---|
| Basil | Fenugreek | Dill | Chervil | Basil | Fenugreek | Dill | Chervil | |
| T0 | 2.8 ± 0.06 byAz | 3.2 ± 0.05 cA | 0.9 ± 0.025 bC | 1.3 ± 0.014 cB | 84.6 ± 10.2 cB | 88.7 ± 12.6 bA | 72.7 ± 9.3 cC | 60.4 ± 5.78 cD |
| T1 | 3 ± 0.023 bB | 3.8 ± 0.03 bA | 1.16 ± 0.086 bD | 2.3 ± 0.043 b C | 90.4 ± 11 bA | 92.9 ± 11.8 bA | 89.6 ± 7.011 bA | 64.7 ± 8.45 bB |
| T2 | 5.6 ± 0.04 aB | 6.2 ± 0.033 aA | 5.4 ± 0.094 aB | 5.7 ± 0.026 aB | 95 ± 9.9 aC | 102.4 ± 14.3 aB | 111.3 ± 4.3 aA | 70.9 ± 6.05 aD |
| Treatment | Protein Content (mg g−1 FW) | |||
|---|---|---|---|---|
| Basil | Fenugreek | Dill | Chervil | |
| T0 | 12.6 ± 1.005 cyBz | 15.3 ± 1.63 cA | 9.4 ± 1.02 aC | 10.3 ± 0.94 bC |
| T1 | 16.3 ± 1.95 bB | 18.4 ± 1.86 bA | 7.8 ± 1.36 bD | 12.6 ± 1.57 aC |
| T2 | 18.6 ± 1.87 aB | 22.5 ± 2.07 aA | 4.9 ± 1.03 cD | 13.2 ± 1.05 aC |
| Reference | Species | Lighting Conditions | Glucose (mg g−1 FW) | Proteins (mg g−1 FW) | Proline (mg g−1 FW) |
|---|---|---|---|---|---|
| This study | Ocimum basilicum L. | Artificial white light | 2.8 ± 0.06 byAz | 12.6 ± 1.005 cyBz | 84.6 ± 10.2 cB |
| Blue LED light | 3 ± 0.023 bB | 16.3 ± 1.95 bB | 90.4 ± 11 bA | ||
| Red LED light | 5.6 ± 0.04 aB | 18.6 ± 1.87 aB | 95 ± 9.9 aC | ||
| Trigonella foenum-graecum | Artificial white light | 3.2 ± 0.05 cA | 15.3 ± 1.63 cA | 88.7 ± 12.6 bA | |
| Blue LED light | 3.8 ± 0.03 bA | 18.4 ± 1.86 bA | 92.9 ± 11.8 bA | ||
| Red LED light | 6.2 ± 0.033 aA | 22.5 ± 2.07 aA | 102.4 ± 14.3 aB | ||
| Anethum graveolens | Artificial white light | 0.9 ± 0.025 bC | 9.4 ± 1.02 aC | 72.7 ± 9.3 cC | |
| Blue LED light | 1.16 ± 0.086 bD | 7.8 ± 1.36 bD | 89.6 ± 7.011 bA | ||
| Red LED light | 5.4 ± 0.094 aB | 4.9 ± 1.03 cD | 111.3 ± 4.3 aA | ||
| Anthriscus cerefolium | Artificial white light | 1.3 ± 0.014 cB | 10.3 ± 0.94 bC | 60.4 ± 5.78 cD | |
| Blue LED light | 2.3 ± 0.043 b C | 12.6 ± 1.57 aC | 64.7 ± 8.45 bB | ||
| Red LED light | 5.7 ± 0.026 aB | 13.2 ± 1.05 aC | 70.9 ± 6.05 aD | ||
| [45] | Brassica campestris L. | Artificial white light | 13.3 c | 12.5 b | |
| Blue LED light | 37.5 b | 8.6 bc | |||
| Red LED light | 56.7 a | 17.5 a | |||
| [46] | Malus x domestica | Artificial white light | 20.2 b | ||
| Blue LED light | 24.4 ab | ||||
| Red LED light | 30.8 a | ||||
| [47] | Solanum lycopersicum | Artificial white light | 1.7 b | ||
| Blue LED light | 1.25 c | ||||
| Red LED light | 1.8 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Haddaji, H.; Akodad, M.; Skalli, A.; Moumen, A.; Bellahcen, S.; Elhani, S.; Urrestarazu, M.; Kolar, M.; Imperl, J.; Petrova, P.; et al. Effects of Light-Emitting Diodes (LEDs) on Growth, Nitrates and Osmoprotectant Content in Microgreens of Aromatic and Medicinal Plants. Horticulturae 2023, 9, 494. https://doi.org/10.3390/horticulturae9040494
El Haddaji H, Akodad M, Skalli A, Moumen A, Bellahcen S, Elhani S, Urrestarazu M, Kolar M, Imperl J, Petrova P, et al. Effects of Light-Emitting Diodes (LEDs) on Growth, Nitrates and Osmoprotectant Content in Microgreens of Aromatic and Medicinal Plants. Horticulturae. 2023; 9(4):494. https://doi.org/10.3390/horticulturae9040494
Chicago/Turabian StyleEl Haddaji, Hamza, Mustapha Akodad, Ali Skalli, Abdelmajid Moumen, Said Bellahcen, Sliman Elhani, Miguel Urrestarazu, Mitja Kolar, Jernej Imperl, Petranka Petrova, and et al. 2023. "Effects of Light-Emitting Diodes (LEDs) on Growth, Nitrates and Osmoprotectant Content in Microgreens of Aromatic and Medicinal Plants" Horticulturae 9, no. 4: 494. https://doi.org/10.3390/horticulturae9040494