
Systematic Analysis of Two Tandem GGDEF/EAL Domain Genes Regulating Antifungal Activities in Pseudomonas glycinae MS82
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Construction of PafQ and PafR Deletion Mutants
2.3. Bioassay of Fungistatic Activity
2.4. qRT-PCR Analysis
2.5. Analysis of Biofilm Formation
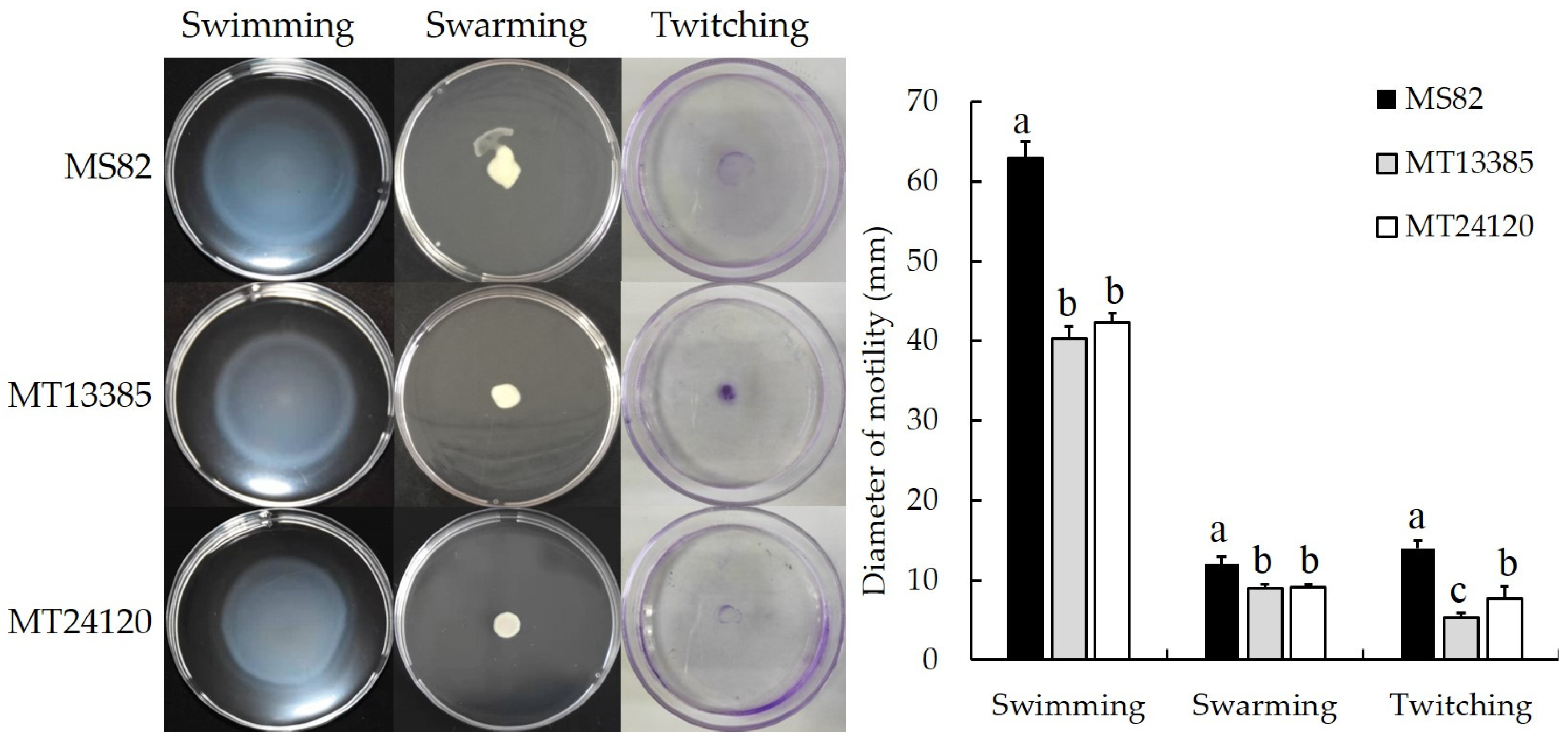
2.6. Analysis of Motility
3. Results
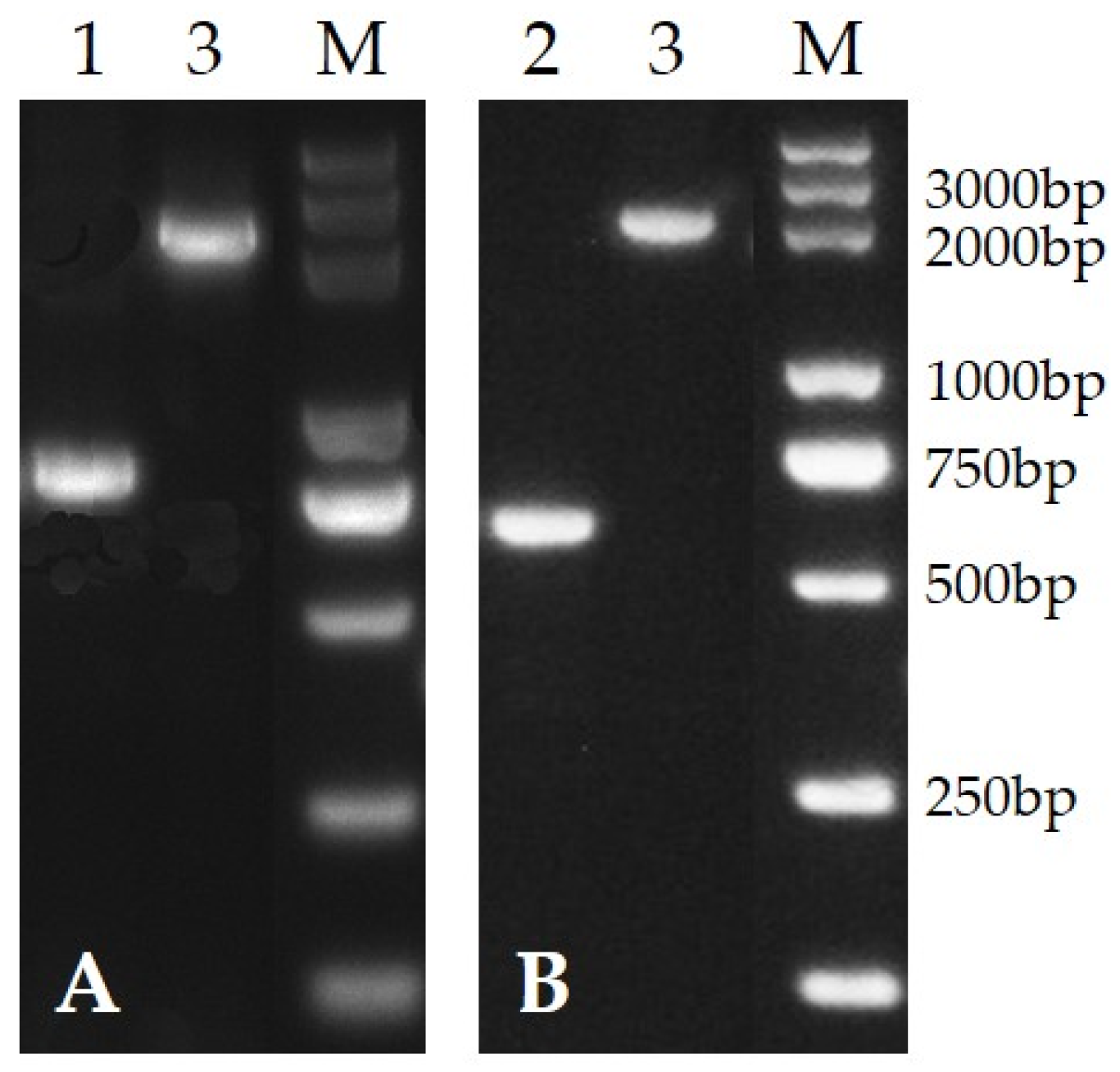
3.1. Deletion of PafQ and PafR
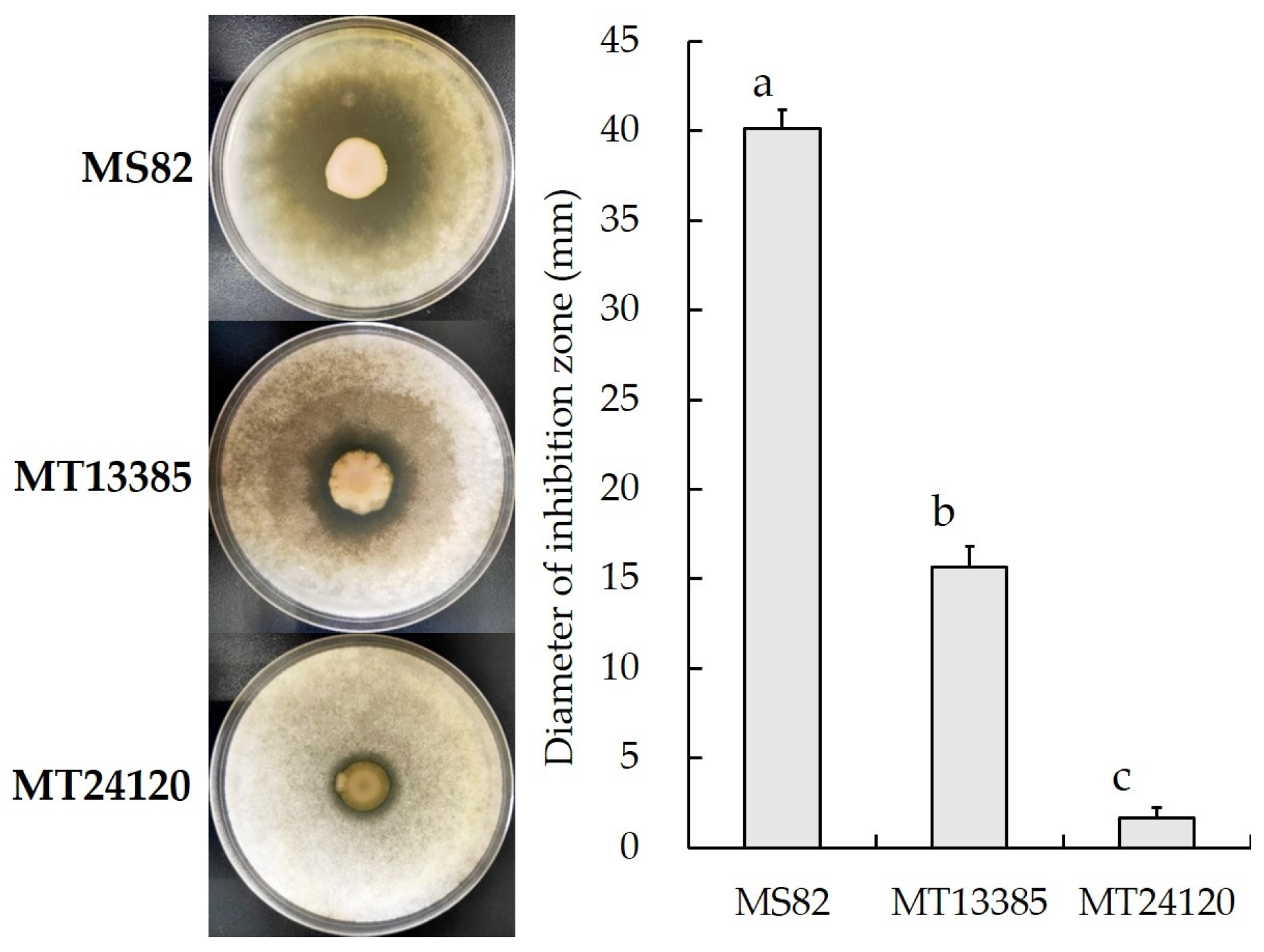
3.2. Bioassay of Fungistatic Activity
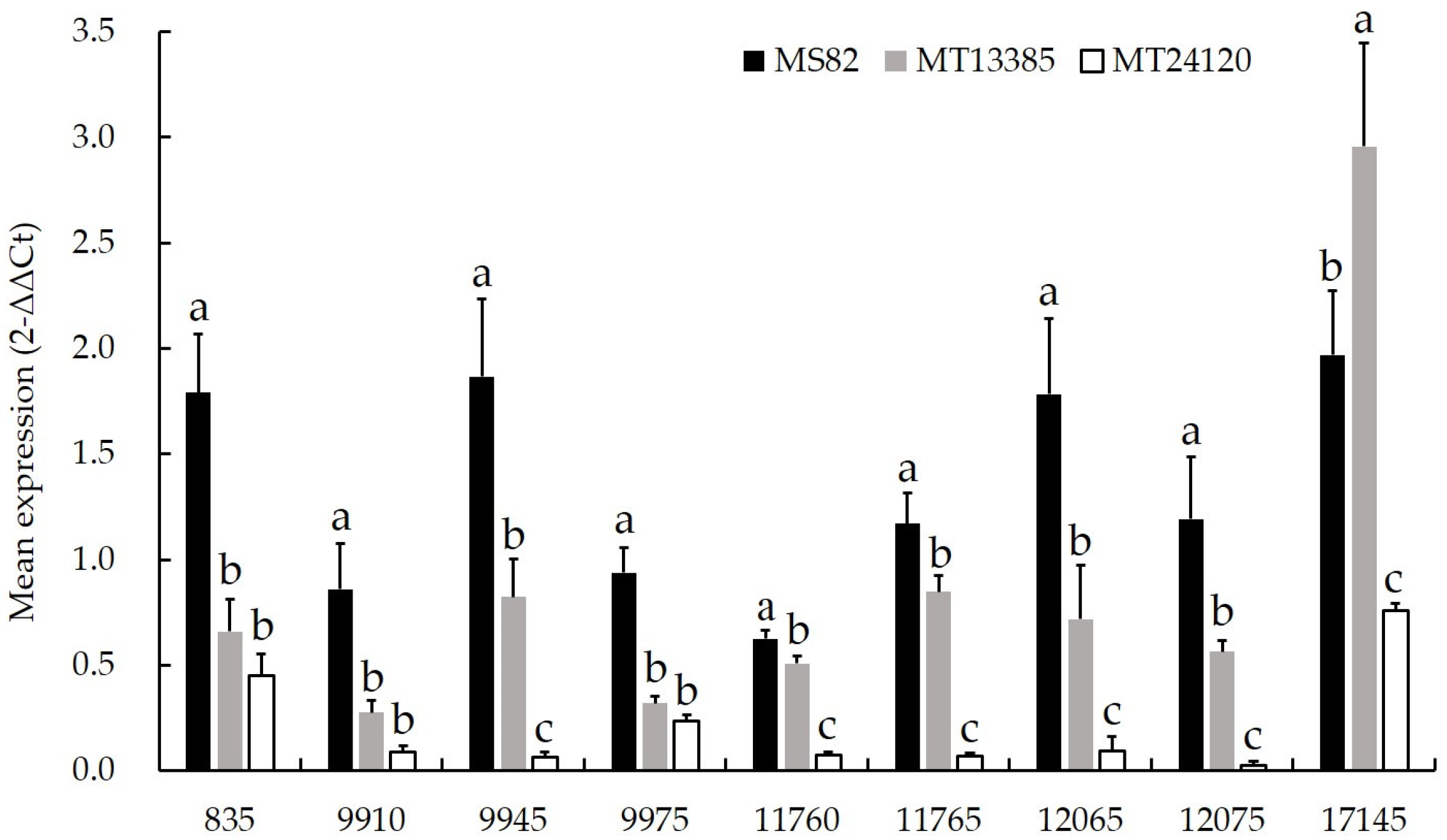
3.3. qRT-PCR Analysis
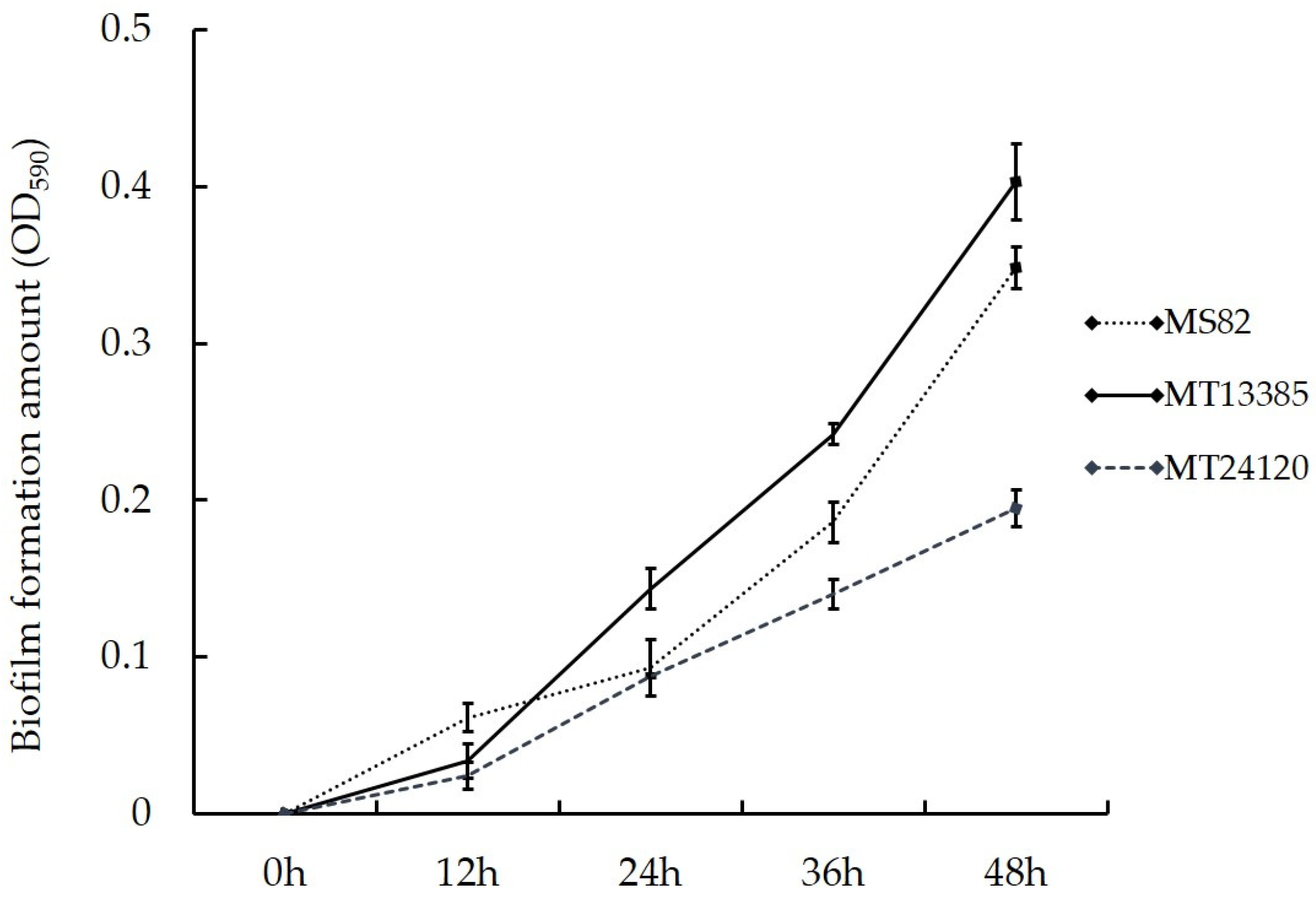
3.4. Analysis of Biofilm Formation
3.5. Analysis of Motility
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, K.; Newman, M.; McInroy, J.A.; Hu, C.H.; Kloepper, J.W. Selection and assessment of plant growth-promoting rhizobacteria for biological control of multiple plant diseases. Phytopathology 2017, 107, 928–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montelongo-Martínez, L.F.; Hernández-Méndez, C.; Muriel-Millan, L.F.; Hernández-Estrada, R.; Fabian-Del Olmo, M.J.; González-Valdez, A.; Soberón-Chávez, G.; Cocotl-Yañez, M. Unraveling the regulation of pyocyanin synthesis by RsmA through MvaU and RpoS in Pseudomonas aeruginosa ID4365. J. Basic Microbiol. 2023, 63, 51–63. [Google Scholar] [CrossRef]
- Zhang, J.; Mavrodi, D.V.; Yang, M.; Thomashow, L.S.; Weller, D.M. Pseudomonas synxantha 2-79 transformed with pyrrolnitrin biosynthesis genes has improved biocontrol activity against soilborne pathogens of wheat and canola. Phytopathology 2020, 110, 1010–1017. [Google Scholar] [CrossRef]
- Gutiérrez-García, K.; Neira-González, A.; Pérez-Gutiérrez, R.M.; Granados-Ramírez, G.; Zarraga, R.; Wrobel, K.; Barona-Gómez, F.; Flores-Cotera, B.L. Phylogenomics of 2,4-diacetylphloroglucinol-producing Pseudomonas and novel antiglycation endophytes from Piper auritum. J. Nat. Prod. 2017, 80, 1955–1963. [Google Scholar] [CrossRef]
- Zhang, Q.X.; Kong, X.W.; Li, S.Y.; Chen, X.J.; Chen, X.J. Antibiotics of Pseudomonas protegens FD6 are essential for biocontrol activity. Australas. Plant Pathol. 2020, 49, 307–317. [Google Scholar] [CrossRef]
- Pandey, S.; Gupta, S. Evaluation of Pseudomonas sp. for its multifarious plant growth promoting potential and its ability to alleviate biotic and abiotic stress in tomato (Solanum lycopersicum) plants. Sci. Rep. 2020, 10, 20951. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z. Analysis of the complete genome sequence of a rhizosphere-derived Pseudomonas sp. HN3-2 leads to the characterization of a cyclic lipopeptide-type antibiotic bananamide C. 3 Biotech 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Soler-Rivas, C.; Arpin, N.; Olivier, J.M.; Wichers, H.J. WLIP, a lipodepsipeptide of Pseudomonas ‘reactans’, as inhibitor of the symptoms of the brown blotch disease of Agaricus bisporus. J. Appl. Microbiol. 1999, 86, 635–641. [Google Scholar] [CrossRef]
- Tajalipour, S.; Hassanzadeh, N.; Jolfaee, H.K.; Heydarib, A.; Ghasemib, A. Biological control of mushroom brown blotch disease using antagonistic bacteria. Biocontrol Sci. Technol. 2014, 24, 473–484. [Google Scholar] [CrossRef]
- Ma, L.; Wang, X.; Deng, P.; Baird, S.M.; Liu, Y.; Qu, S.; Lu, S. The PafR gene is required for antifungal activity of strain MS82 against Mycogone perniciosa. Adv. Microbiol. 2017, 7, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Qu, S.; Lin, J.; Jia, J.; Baird, S.M.; Jiang, N.; Li, H.; Hou, L.; Lu, S. The complete genome of the antifungal bacterium Pseudomonas sp. strain MS82. J. Plant Dis. Protect. 2019, 126, 153–160. [Google Scholar] [CrossRef]
- Tagua, V.G.; Molina-Henares, M.A.; Travieso, M.L.; Nisa-Martínez, R.; Quesada, J.M.; Espinosa-Urgel, M.; Ramos-González, M.I. C-di-GMP and biofilm are regulated in Pseudomonas putida by the CfcA/CfcR two-component system in response to salts. Environ. Microbiol. 2022, 24, 158–178. [Google Scholar] [CrossRef] [PubMed]
- Cutruzzolà, F.; Paiardini, A.; Rossi, C.S.; Spizzichino, S.; Paone, A.; Giardina, G.; Rinaldo, S. A conserved scaffold with heterogeneous metal ion binding site: The multifaceted example of HD-GYP proteins. Coordin. Chem. Rev. 2022, 450, 214228. [Google Scholar] [CrossRef]
- Verma, R.K.; Biswas, A.; Kakkar, A.; Lomada, S.K.; Pradhan, B.B.; Chatterjee, S. A bacteriophytochrome mediates interplay between light sensing and the second messenger cyclic di-GMP to control social behavior and virulence. Cell Rep. 2020, 32, 108202. [Google Scholar] [CrossRef]
- Ribbe, J.; Baker, A.E.; Euler, S.; O’Toole, G.A.; Maier, B. Role of cyclic di-GMP and exopolysaccharide in type IV pilus dynamics. J. Bacteriol. 2017, 199, e00859-16. [Google Scholar] [CrossRef] [Green Version]
- Gdaniec, B.G.; Allard, P.-M.; Queiroz, E.F.; Wolfender, J.-L.; van Delden, C.; Köhler, T. Surface sensing triggers a broad-spectrum antimicrobial response in Pseudomonas aeruginosa. Environ. Microbiol. 2020, 22, 3572–3587. [Google Scholar] [CrossRef]
- Blanco-Romero, E.; Garrido-Sanz, D.; Durán, D.; Rivilla, R.; Redondo-Nieto, M.; Martín, M. Regulation of extracellular matrix components by AmrZ is mediated by c-di-GMP in Pseudomonas ogarae F113. Sci. Rep. 2022, 12, 11914. [Google Scholar] [CrossRef] [PubMed]
- Christen, M.; Christen, B.; Folcher, M.; Schauerte, A.; Jenal, U. Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J. Biol. Chem. 2005, 280, 30829–30837. [Google Scholar] [CrossRef] [Green Version]
- Römling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef] [Green Version]
- Muriel, C.; Blanco-Romero, E.; Trampari, E.; Arrebola, E.; Durán, D.; Redondo-Nieto, M.; Malone, J.G.; Martín, M.; Rivilla, R. The diguanylate cyclase AdrA regulates flagellar biosynthesis in Pseudomonas fluorescens F113 through SadB. Sci. Rep. 2019, 9, 8096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Xia, J.; Su, Z.; Xu, G.; Gomelsky, M.; Qian, G.; Liu, F. Lysobacter PilR, the regulator of type IV pilus synthesis, controls antifungal antibiotic production via a cyclic di-GMP pathway. Appl. Environ. Microbiol. 2017, 83, e03397-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennox, E.S. Transduction of linked genetic characters of the host by bacteriophage P1. Virology 1955, 1, 190–206. [Google Scholar] [CrossRef]
- Jia, J.; Wang, X.; Deng, P.; Ma, L.; Baird, M.S.; Li, X.; Lu, S.E. Pseudomonas glycinae sp. nov. isolated from the soybean rhizosphere. MicrobiologyOpen 2020, 9, e1101. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qu, S.; Luo, X.; Lu, S.-E.; Liu, Y.; Li, H.; Hou, L.; Lin, J.; Jiang, N.; Ma, L. PafS containing GGDEF-domain regulates life activities of Pseudomonas glycinae MS82. Microorganisms 2022, 10, 2342. [Google Scholar] [CrossRef]
- Wang, X.Q.; Liu, A.X.; Guerrero, A.; Liu, J.; Yu, X.Q.; Deng, P.; Ma, L.; Baird, S.M.; Smith, L.; Li, X.D.; et al. Occidiofungin is an important component responsible for the antifungal activity of Burkholderia pyrrocinia strain Lyc2. J. Appl. Microbiol. 2015, 120, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Huertas-Rosales, Ó.; Romero, M.; Heeb, S.; Espinosa-Urgel, M.; Camara, M.; Ramos-Gonzalez, M.I. The Pseudomonas putida CsrA/RsmA homologues negatively affect c-di-GMP pools and biofilm formation through the GGDEF/EAL response regulator CfcR. Envion. Microbiol. 2017, 19, 3551–3566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smet, J.D.; Wagemans, J.; Hendrix, H.; Staes, I.; Visnapuu, A.; Horemans, B.; Aertsen, A.; Lavigne, R. Bacteriophage mediated interference of the c-di-GMP signaling pathway in Pseudomonas aeruginosa. Microb. Biotechnol. 2020, 14, 967–978. [Google Scholar] [CrossRef]
- Cai, Y.; Hutchin, A.; Craddock, J.; Walsh, M.A.; Webb, J.S.; Tews, I. Differential impact on motility and biofilm dispersal of closely related phosphodiesterases in Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 6232. [Google Scholar] [CrossRef] [Green Version]
- Meissner, A.; Wild, V.; Simm, R.; Rohde, M.; Erck, C.; Bredenbruch, F.; Morr, M.; Römling, U.; Hussler, S. Pseudomonas aeruginosa cupA-encoded fimbriae expression is regulated by a GGDEF and EAL domain-dependent modulation of the intracellular level of cyclic diguanylate. Envion. Microbiol. 2007, 9, 2475–2485. [Google Scholar] [CrossRef]
- Valentini, M.; Filloux, A. Biofilms and cyclic di-GMP (c-di-GMP) signaling: Lessons from Pseudomonas aeruginosa and other bacteria. J. Biol. Chem. 2016, 291, 12547–12555. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Zhang, B.; Yang, Q.; Zhang, Y.; Zheng, D.; Zhang, L.Q.; Yan, Q.; Wu, X. Cyclic di-GMP regulates the quorum-sensing system and biocontrol activity of Pseudomonas fluorescens 2P24 through the RsmA and RsmE proteins. Appl. Environ. Microbiol. 2020, 86, e02016-20. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y. Functional Analysis of the GGDEF/EAL Domain Protein PigX in Serratia plymuthica G3. Ph.D. Thesis, Jiangsu University, Zhenjiang, China, 2016. [Google Scholar]
- Fineran, P.C.; Williamson, N.R.; Lilley, K.S.; Salmond, G.P.C. Virulence and prodigiosin antibiotic biosynthesis in Serratia are regulated pleiotropically by the GGDEF/EAL domain protein, PigX. J. Bacteriol. 2007, 189, 7653–7662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, C.; Christensen, M.N.; Kovács, T. Molecular aspects of plant growth promotion and protection by Bacillus subtilis. Mol. Plant Microbe Interact. 2021, 34, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Allard-Massicotte, R.; Tessier, L.; Lecuyer, F.; Lakshmanan, V.; Lucier, J.-F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involves multiple chemotaxis receptors. mBio 2016, 7, e01664-16. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.E.; Diepold, A.; Kuchma, S.L.; Scott, J.E.; Ha, D.G.; Orazi, G.; Armitage, J.P.; O’Toole, G.A. PilZ domain protein FlgZ mediates cyclic di-GMP-dependent swarming motility control in Pseudomonas aeruginosa. J. Bacteriol. 2016, 198, 1837–1846. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Xin, L.; Zeng, Y.; Yam, J.K.H.; Ding, Y.; Venkataramani, P.; Cheang, Q.W.; Yang, X.; Tang, X.; Zhang, L.-H.; et al. A cyclic di-GMP-binding adaptor protein interacts with a chemotaxis methyltransferase to control flagellar motor switching. Sci. Signal. 2016, 9, ra102/1–ra102/12. [Google Scholar] [CrossRef]
- Yan, X.-F.; Xin, L.; Yen, J.T.; Zeng, Y.; Jin, S.; Cheang, Q.W.; Fong, R.A.C.Y.; Chiam, K.-H.; Liang, Z.-X.; Gao, Y.-G. Structural analyses unravel the molecular mechanism of cyclic di-GMP regulation of bacterial chemotaxis via a PilZ adaptor protein. J. Biol. Chem. 2018, 293, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Deepa, N.; Chauhan, S.; Kumari, P.; Rai, A.K.; Tandon, S.; Singh, A. Linalool reduces the virulence of Pseudomonas syringae pv. tomato DC 3000 by modulating the PsyI/PsyR quorum-sensing system. Microb. Pathog. 2022, 173, 105884. [Google Scholar] [CrossRef]
- Bense, S.; Witte, J.; Preuße, M.; Koska, M.; Pezoldt, L.; Dröge, A.; Hartmann, O.; Müsken, M.; Schulze, J.; Fiebig, T.; et al. Pseudomonas aeruginosa post-translational responses to elevated c-di-GMP levels. Mol. Microbiol. 2022, 117, 1213–1226. [Google Scholar] [CrossRef]
- Jain, R.; Sliusarenko, O.; Kazmierczak, B.I. Interaction of the cyclic-di-GMP binding protein FimX and the Type 4 pilus assembly ATPase promotes pilus assembly. PLoS Pathog. 2017, 13, e1006594. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, G.B.; Marmont, L.S.; Ostaszewski, A.; Rich, J.D.; Whitney, J.C.; Parsek, M.R.; Harrison, J.J.; Howell, P.L. Pel polysaccharide biosynthesis requires an inner membrane complex comprised of PelD, PelE, PelF, and PelG. J. Bacteriol. 2020, 202, e00684-19. [Google Scholar] [CrossRef]
- Roelofs, K.G.; Jones, C.J.; Helman, S.R.; Shang, X.; Orr, M.W.; Goodson, J.R.; Galperin, M.Y.; Yildiz, F.H.; Lee, V.T. Systematic identification of cyclic-di-GMP binding proteins in Vibrio cholerae reveals a novel class of cyclic-di-GMP-binding ATPases associated with type II secretion systems. PLoS Pathog. 2015, 27, e1005232. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Ahmad, I.; Blanka, A.; Schottkowski, M.; Cimdins, A.; Galperin, M.Y.; Römling, U.; Gomelsky, M. GIL, a new c-di-GMP-binding protein domain involved in regulation of cellulose synthesis in enterobacteria. Mol. Microbiol. 2014, 93, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, E.; Seminara, A.B.; Kim, S.; Hall, C.L.; Wang, Y.; Lee, V.T. Thiol-benzotriazolo-quinazolinone inhibits Alg44 binding to c-di-GMP and reduces alginate production by Pseudomonas aeruginosa. ACS Chem. Biol. 2017, 12, 3076–3085. [Google Scholar] [CrossRef] [PubMed]
- Molina-Henares, M.A.; Ramos-González, M.I.; Daddaoua, A.; Fernández-Escamilla, A.M.; Espinosa-Urgel, M. FleQ of Pseudomonas putida KT2440 is a multimeric cyclic diguanylate binding protein that differentially regulates expression of biofilm matrix components. Res. Microbiol. 2017, 168, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Chuah, M.L.C.; Dong, X.; Xie, K.; Luo, Z.; Tang, K.; Liang, Z.X. Binding of cyclic diguanylate in the non-catalytic EAL domain of FimX induces a long-range conformational change. J. Biol. Chem. 2011, 286, 2910–2917. [Google Scholar] [CrossRef] [Green Version]
- Nie, H.; Xiao, Y.; He, J.; Liu, H.; Nie, L.; Chen, W.; Huang, Q. Phenotypic–genotypic analysis of GGDEF/EAL/HD-GYP domain-encoding genes in Pseudomonas putida. Environ. Microbiol. Rep. 2020, 12, 38–48. [Google Scholar] [CrossRef]
- Tal, R.; Wong, H.C.; Calhoon, R.; Gelfand, D.; Benziman, M. Three cdg operons control cellular turnover of cyclic di-GMP in Acetobacter xylinum: Genetic organization and occurrence of conserved domains in isoenzymes. J. Bacteriol. 1998, 180, 4416–4425. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Wang, Y.; Cen, C.; Fu, L.; Wang, Y. A tandem GGDEF-EAL domain protein-regulated c-di-GMP signal contributes to spoilage-related activities of Shewanella baltica OS155. Appl. Microbiol. Biotechnol. 2020, 104, 2205–2216. [Google Scholar] [CrossRef]
- Wang, Y.; Hay, I.D.; Rehman, Z.U.; Rehm, B.H.A. Membrane-anchored MucR mediates nitrate-dependent regulation of alginate production in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2015, 99, 7253–7265. [Google Scholar] [CrossRef]
- Hay, I.D.; Remminghorst, U.; Rehm, B.H.A. MucR, a novel membrane-associated regulator of alginate biosynthesis in Pseudomonas aeruginosa. Appl. Environ. Microb. 2009, 75, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Heine, S.; Entian, M.; Sauer, K.; Frankenberg-Dinkel, N. NO-induced biofilm dispersion in Pseudomonas aeruginosa is mediated by an MHYT domain-coupled phosphodiesterase. J. Bacteriol. 2013, 195, 3531–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phippen, C.W.; Mikolajek, H.; Schlaefli, H.G.; Keevil, C.W.; Webb, J.S.; Tews, I. Formation and dimerization of the phosphodiesterase active site of the Pseudomonas aeruginosa MorA, a bi-functional c-di-GMP regulator. FEBS Lett. 2014, 588, 4631–4636. [Google Scholar] [CrossRef] [Green Version]
- Choy, W.-K.; Zhou, L.; Syn, C.K.-C.; Zhang, L.-H.; Swarup, S. MorA defines a new class of regulators affecting flagellar development and biofilm formation in diverse Pseudomonas species. J. Bacteriol. 2004, 186, 7221–7228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichandran, A.; Ramachandran, M.; Suriyanarayanan, T.; Wong, C.C.; Swarup, S. Global regulator MorA affects virulence-associated protease secretion in Pseudomonas aeruginosa PAO1. PLoS ONE 2015, 10, e0123805. [Google Scholar] [CrossRef] [PubMed]
- Katharios-Lanwermeyer, S.; Whitfield, G.B.; Howell, P.L.; O’Toole, G.A. Pseudomonas aeruginosa uses c-di-GMP phosphodiesterases RmcA and MorA to regulate biofilm maintenance. mBio 2021, 12, e03384-20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Genotype or Phenotype 1 | Source |
|---|---|---|
| Strains | ||
| E. coli | ||
| DH5α | supE44 ΔlacU169 (Φ80 lacZΔM15) hsdR17 recA1 endA1 gyrA96 thi-1 relA1 | TSINGKE, Beijing, China |
| S17-1λ | RP4-2(Km::Tn7,Tc::Mu-1), pro-82, LAMpir, recA1, endA1, thiE1, hsdR17, creC510 | WEIDI, Shanghai, China |
| P. gylcinae | ||
| MS82 | Ampr 1, Wild type | [23] |
| MT13385 | Ampr, PafQ deletion mutant derived from MS82 | This work |
| MT24120 | Ampr, PafR deletion mutant derived from MS82 | This work |
| T. virens | ||
| NJ1 | A fungal pathogen collected from Pleurotus ostreatus substrate | Lab stock |
| Plasmids and vectors | ||
| pEX18GM | Gmr, Suicide vector, SacB | FENGHUI, Changsha, China |
| pEX18-13385 | Gmr, up- and down-stream region of PafQ in PEX18GM | This work |
| pEX18-24120 | Gmr, up- and down-stream region of PafR in PEX18GM | This work |
| Primer Name | Sequence 5′→3′ | Restriction Enzyme Site 1 | Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| 13385-F1 13385-R1 | CGGAATTCGAATACCTCAACAGACACTC 1 GGGGTACCCCTTGGCAGTAGCAATGCGC | Eco RI Kpn I | 387 | 55 |
| 13385-F2 13385-R2 | GGGGTACCGGAAATCACCGAAACCACCG GCTCTAGAATGACCTGCGATTAGCGGCT | Kpn I Xba I | 425 | 55 |
| 24120-F1 24120-R1 | CGGAATTCCAGCCAGACCGCAGGATTAC GGGGTACCGCTGTCGCTGAGGATTTTTC | Eco RI Kpn I | 404 | 55 |
| 24120-F2 24120-R2 | GGGGTACCGCGACGAGGTTCAGGGTTAT GCTCTAGAATCGGGGGCAGAAAAGGGG | Kpn I Xba I | 302 | 55 |
| Gene Locus_tag 1 | Gene Function | Primer Name | Primer Sequence 5′→3′ |
|---|---|---|---|
| DBV33_00020 | DNA gyrase subunit B | gyrB-F | CGGCACCCAGATTCACTT |
| gyrB-R | GGAGTTGAGGAAGGACAGTT | ||
| DBV33_00835 | adenylyl-sulfate kinase | 835-F | AGTCGTGGTCTGCAAAGTGT |
| 835-R | CTGCACCAGACCTCGCAATA | ||
| DBV33_09910 | non-ribosomal peptide synthetase (NRPS) | 9910-F | CGTCAGACTGCTCAACACCT |
| 9910-R | TTGACCGATCGGCATTGTCA | ||
| DBV33_09945 | membrane dipeptidase | 9945-F | ATCGGGTTCAAGGACAACCC |
| 9945-R | TCCTTGTCGACTTCGTTCCC | ||
| DBV33_09975 | ornithine monooxygenase | 9975-F | CAACAATGCCACCGGTGAAG |
| 9975-R | AAGCCCTGCATGTACAGACC | ||
| DBV33_11760 | NRPS | 11760-F | CTGGCAGCGATCCATGTGTA |
| 11760-R | TGAATGACAACTCGCGACCA | ||
| DBV33_11765 | NRPS | 11765-F | AGCATCTGGACGAACCTGTG |
| 11765-R | CAGGTCGAGGCGGAAGTATC | ||
| DBV33_12065 | NRPS | 12065-F | GTTGAAGTGTGGCCGTTGTC |
| 12065-R | CGTTGCTGATCCGGACGATA | ||
| DBV33_12075 | NRPS | 12075-F | GGTATCGGGCCAATCCTGAG |
| 12075-R | ACCGCTGCCACTCAAATACA | ||
| DBV33_17145 | (2Fe-2S)-binding protein | 17145-F | CAATGCACTGCCTAGAAAGAACC |
| 17145-R | TGAGCACGGTTTCGCCAATAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, J.; Qu, S.; Chen, X.; Li, H.; Hou, L.; Lu, S.-E.; Xu, P.; Jiang, N.; Ma, L. Systematic Analysis of Two Tandem GGDEF/EAL Domain Genes Regulating Antifungal Activities in Pseudomonas glycinae MS82. Horticulturae 2023, 9, 446. https://doi.org/10.3390/horticulturae9040446
Lin J, Qu S, Chen X, Li H, Hou L, Lu S-E, Xu P, Jiang N, Ma L. Systematic Analysis of Two Tandem GGDEF/EAL Domain Genes Regulating Antifungal Activities in Pseudomonas glycinae MS82. Horticulturae. 2023; 9(4):446. https://doi.org/10.3390/horticulturae9040446
Chicago/Turabian StyleLin, Jinsheng, Shaoxuan Qu, Xianyi Chen, Huiping Li, Lijuan Hou, Shi-En Lu, Ping Xu, Ning Jiang, and Lin Ma. 2023. "Systematic Analysis of Two Tandem GGDEF/EAL Domain Genes Regulating Antifungal Activities in Pseudomonas glycinae MS82" Horticulturae 9, no. 4: 446. https://doi.org/10.3390/horticulturae9040446