

In Vitro Micropropagation of Commercial Ginseng Cultivars (Panax ginseng Meyer) via Somatic Embryogenesis Compared to Traditional Seed Production
 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Seed Production and Characteristics
2.3. Somatic Embryogenesis and Plant Regeneration
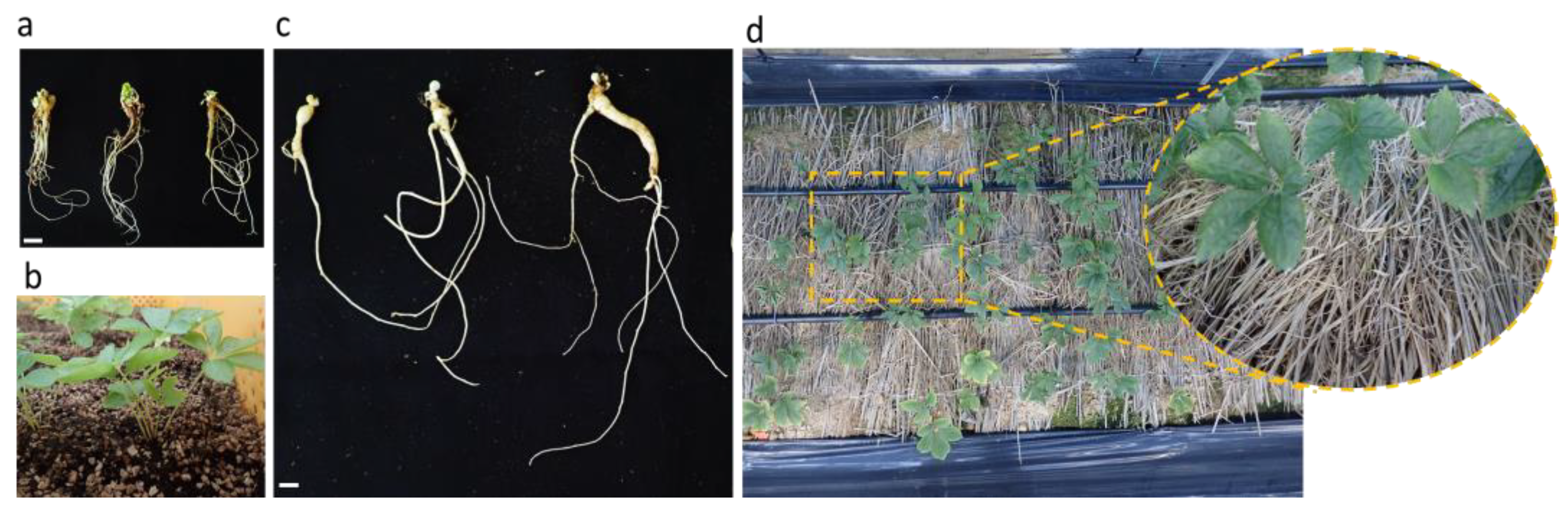
2.4. Acclimatization
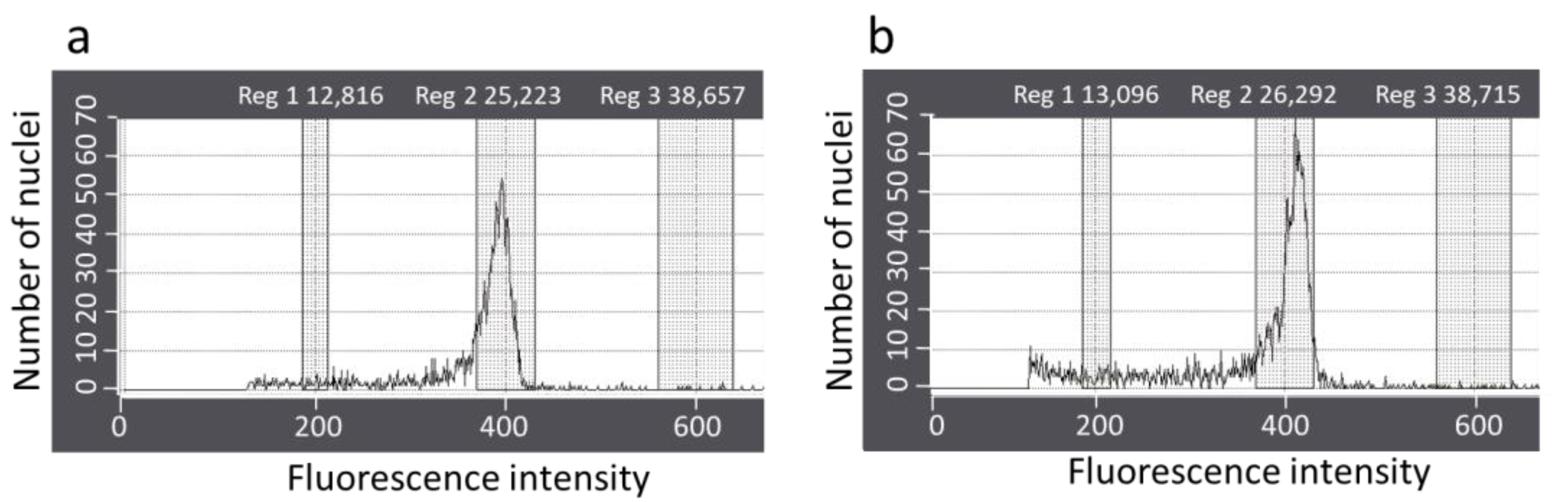
2.5. Ploidy Level Analysis
2.6. Statistical Analysis
3. Results
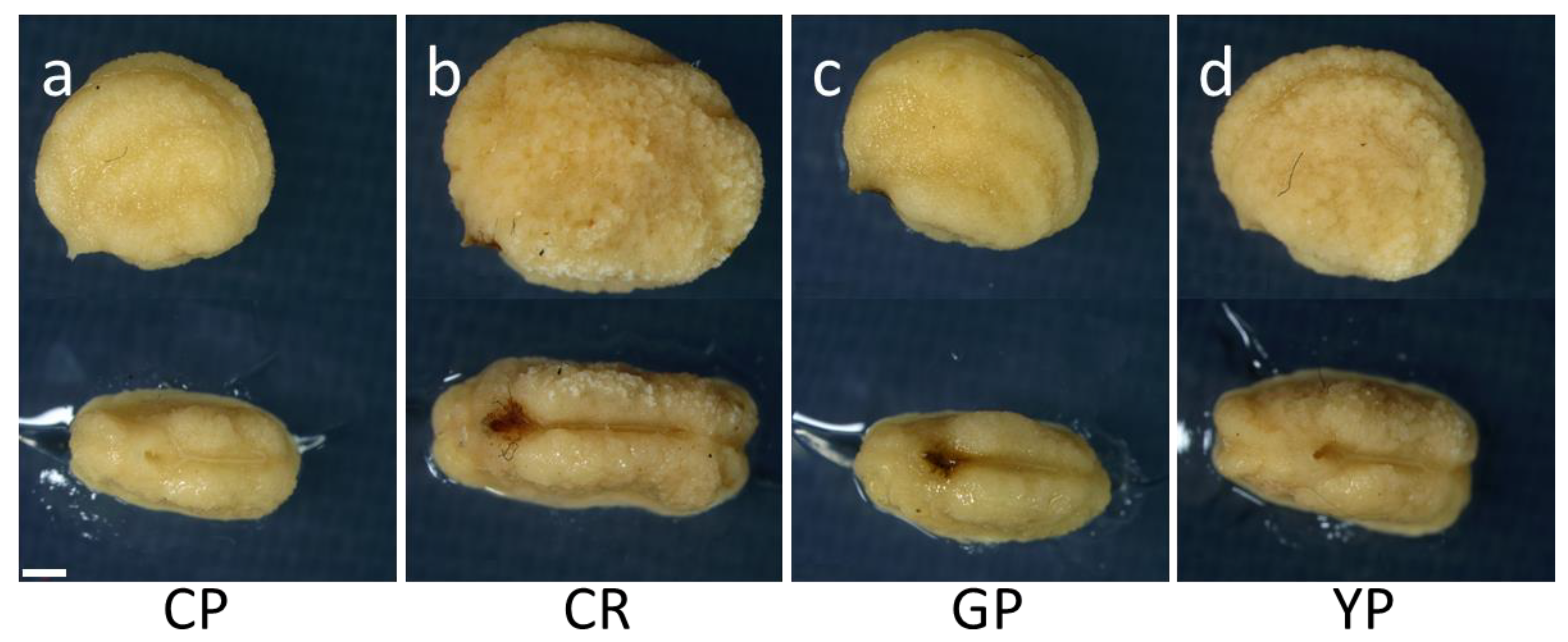
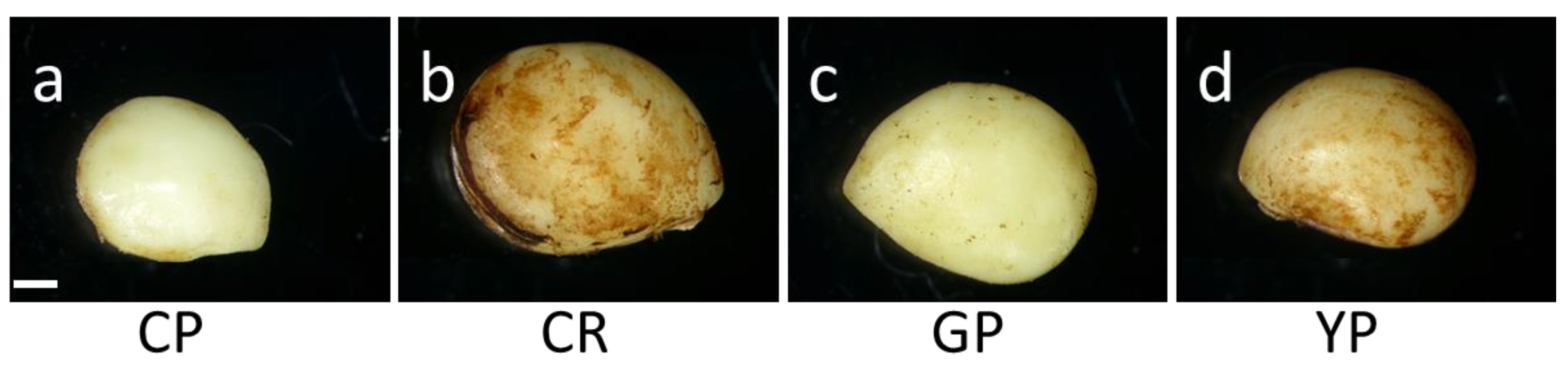
3.1. Seed Production and Characteristics
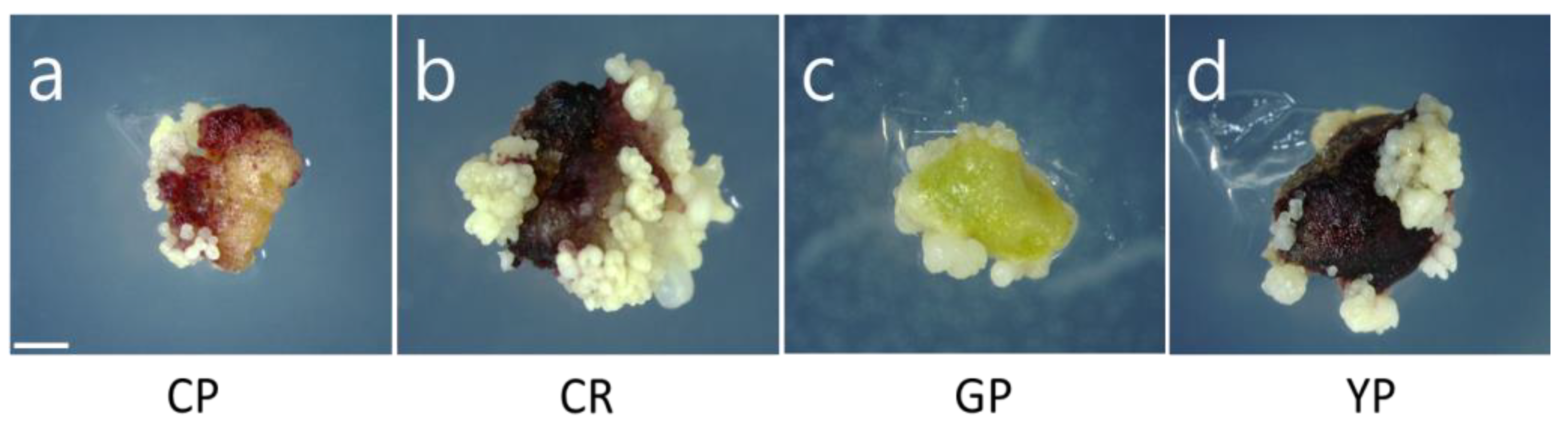
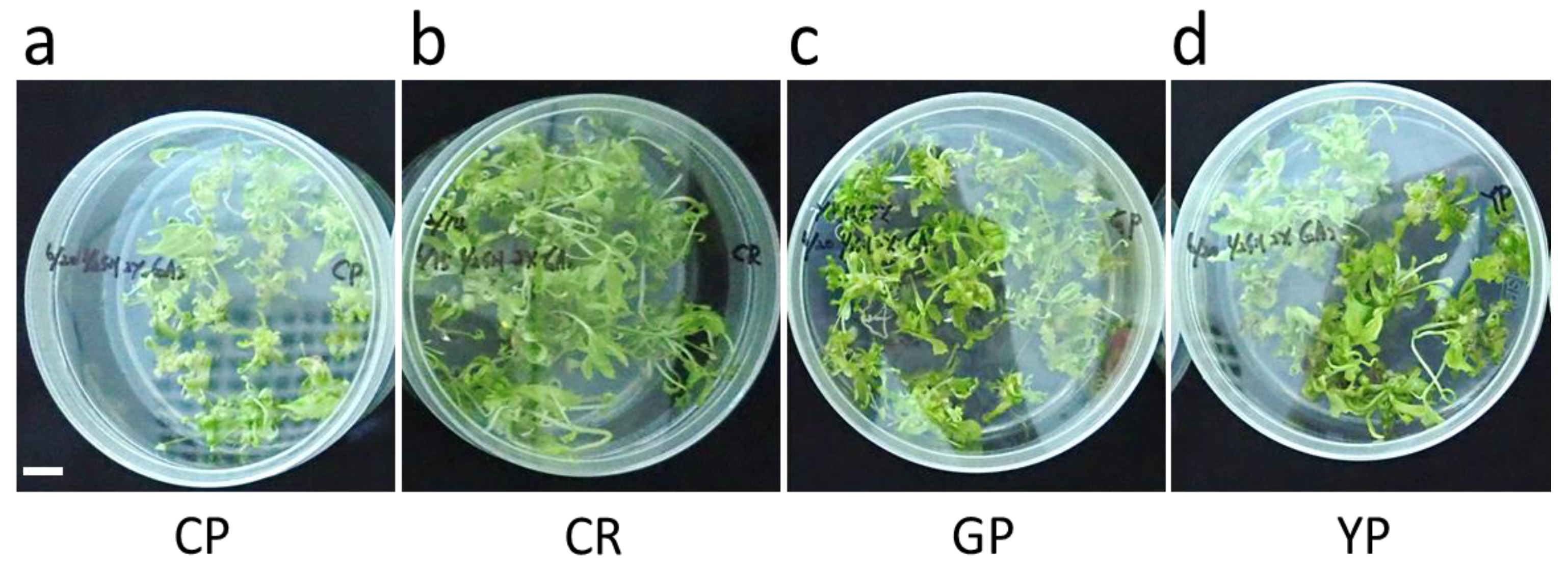
3.2. Somatic Embryogenesis and Plant Regeneration
3.3. Acclimatization and Ploidy Level Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Potenza, M.A.; Montagnani, M.; Santacroce, L.; Charitos, I.A.; Bottalico, L. Ancient herbal therapy: A brief history of Panax ginseng. J. Ginseng Res. 2022. [Google Scholar] [CrossRef]
- Luthra, R.; Roy, A.; Pandit, S.; Prasad, R. Biotechnological methods for the production of ginsenosides. S. Afr. J. Bot. 2021, 141, 25–36. [Google Scholar] [CrossRef]
- Adil, M.; Jeong, B.R. In vitro cultivation of Panax ginseng C.A. Meyer. Indus. Crop. Prod. 2018, 122, 239–251. [Google Scholar] [CrossRef]
- Zhang, H.; Abid, S.; Ahn, J.C.; Mathiyalagan, R.; Kim, Y.J.; Yang, D.C.; Wang, Y. Characteristics of Panax ginseng Cultivars in Korea and China. Molecules 2020, 25, 2635. [Google Scholar] [CrossRef]
- Kwon, W.S.; Kang, J.Y.; Lee, J.H.; Lee, M.G.; Choi, K.T. Red ginseng quality and characteristics of KG101 a promising line of Panax ginseng. J. Ginseng Res. 1998, 22, 244–251. [Google Scholar]
- Kwon, W.S.; Lee, M.G.; Choi, K.T. Breeding process and characteristics of Yunpoong, a new variety of Panax ginseng C.A. Meyer. J. Ginseng Res. 2000, 24, 1–7. [Google Scholar]
- Lee, J.H.; Lee, J.S.; Kwon, W.S.; Kang, J.Y.; Lee, D.Y.; In, J.G.; Kim, Y.S.; Seo, J.; Baeg, I.H.; Chang, I.M.; et al. Characteristics of Korean ginseng varieties of Gumpoong, Sunun, Sunpoong, Sunone, Cheongsun, and Sunhyang. J. Ginseng Res. 2015, 39, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Kim, D.H.; Bang, K.H.; Kim, J.U.; Hyun, D.Y.; Lee, S.W.; Kang, S.W.; Cha, S.W.; Kim, K.H.; Choi, J.K.; et al. A high yielding and salt resistance ginseng variety ‘Cheonryang’. Korean J. Breed. Sci. 2013, 45, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Jo, I.H.; Kim, J.U.; Hong, C.E.; Kim, Y.C.; Kim, D.H.; Park, Y.D. Improvement of seed dehiscence and germination in ginseng by stratification, gibberellin, and/or kinetin treatments. Hortic. Environ. Biotechnol. 2018, 59, 293–301. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, Y.C.; Bang, K.H.; Kim, J.U.; Lee, J.W.; Cho, I.H.; Kim, Y.B.; Lim, J.Y.; Kim, K.H. Flowering and fruits formation characteristics in major varieties of Panax ginseng. Korean Soc. Med. Crop. Sci. 2016, 24, 207–213. [Google Scholar] [CrossRef]
- Koufan, M.; Belkoura, I.; Mazri, M.A. In vitro propagation of caper (Capparis spinosa L.): A review. Horticulturae 2022, 8, 737. [Google Scholar] [CrossRef]
- Guan, Y.; Li, S.G.; Fan, X.F.; Su, Z.H. Application of somatic embryogenesis in woody plants. Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaj, M.D. Direct somatic embryogenesis as a rapid and efficient system for in vitro regeneration of Arabidopsis thaliana. Plant Cell Tiss. Organ Cult. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Zhou, S.; Brown, D.C.W. High efficiency plant production of North American ginseng via somatic embryogenesis from cotyledon explants. Plant Cell Rep. 2006, 25, 166–173. [Google Scholar] [CrossRef]
- Raza, G.; Singh, M.B.; Bhalla, P.L. Somatic embryogenesis and plant regeneration from commercial soybean cultivars. Plants 2020, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Kim, M.K.; Shim, J.S.; Pulla, R.K.; Yang, D.C. Somatic embryogenesis of two new Panax ginseng cultivars, Yun-Poong and Chun-Poong. Russ J. Plant Physiol. 2010, 57, 283–289. [Google Scholar] [CrossRef]
- Lee, J.; Jo, I.; Hong, C.; Bang, K.; Kim, J. Effect of gamma-irradiation on seed dehiscence, development, survival, and growth in Panax ginseng. Korean J. Breed. Sci. 2020, 52, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Do, G.R.; Jo, I.H.; Hong, C.E.; Bang, K.H.; Kim, J.U.; Park, Y.D. Zygotic embryo culture is an efficient way to optimize in vitro growth in Panax ginseng. Indus. Crop. Prod. 2021, 167, 1134971. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.H.; Kim, Y.C.; Kim, K.H.; Han, J.Y.; Choi, Y.E. In vitro grown thickened taproots, a new type of soil transplanting source in Panax ginseng. J. Ginseng Res. 2016, 40, 409–414. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Nagahama, N.; Manifestod, M.M.; Fortunato, R.H. Vegetative propagation and proposal for sustainable management of Valeriana carnosa Sm., a traditional medicinal plant from Patagonia. J. Appl. Res. Med. Aromat. Plants. 2019, 14, 100218. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, Y.Y.; Kim, Y.S.; Balaraju, K.; Mok, Y.S.; Yoo, S.J.; Jeon, Y. Enhancement of seed germination and microbial disinfection on ginseng by cold plasma treatment. J. Ginseng Res. 2021, 45, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Kim, Y.C.; Kim, J.U.; Jo, I.H.; Kim, K.H.; Kim, D.H. Effects of gibberellic acid and alternating temperature on breaking seed dormancy of Panax ginseng C.A. Meyer. Korean Soc. Med. Crop. Sci. 2016, 24, 284–293. [Google Scholar] [CrossRef]
- Min, J.E.; Long, N.P.; Hong, J.Y.; Kim, S.J.; Anh, N.H.; Wang, D.; Wang, X.; Park, J.H.; Kwon, S.W.; Lee, S.J. The dehiscence process in Panax ginseng seeds and the stigmasterol biosynthesis pathway in terms of metabolomics. J. Ginseng Res. 2022, 46, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Shim, C.K.; Kim, Y.K.; Hong, S.J.; Park, J.H.; Han, E.J.; Kim, S.C. Enhancement of Seed Dehiscence by Seed Treatment with Talaromyces flavus GG01 and GG04 in ginseng (Panax ginseng). Plant Pathol. J. 2017, 33, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, D.; Mariz-Ponte, N.; Loureiro, J.; Castro, S.; Debiasi, C.; Domingues, J.; Santos, C.; Gonçalves, J.C. A protocol for micropropagation of the medicinal species Tuberaria lignosa provides ploidy true-to-type plants with high antioxidant capacity. Plant Cell Tiss. Organ Cult. 2022, 150, 599–609. [Google Scholar] [CrossRef]
- Choi, Y.E.; Yang, D.C.; Park, J.C.; Soh, W.Y.; Choi, K.T. Regenerative ability of somatic single and multiple embryos from cotyledons of Korean ginseng on hormone-free medium. Plant Cell Rep. 1998, 17, 544–551. [Google Scholar] [CrossRef]
- Choi, Y.E.; Yang, D.C.; Yoon, E.S.; Choi, K.T. Plant regeneration via adventitious bud formation from cotyledon explants of Panax ginseng C.A. Meyer. Plant Cell Rep. 1998, 17, 731–736. [Google Scholar] [CrossRef]
- Silva, R.C.; Luis, Z.G.; Scherwinski-Pereira, J.E. Differential responses to somatic embryogenesis of different genotypes of Brazilian oil palm (Elaeis guineensis Jacq.). Plant Cell Tiss. Organ Cult. 2012, 111, 59–67. [Google Scholar] [CrossRef]
- Kim, J.Y.; Adhikari, P.B.; Ahn, C.H.; Kim, D.H.; Chang Kim, Y.C.; Han, J.Y.; Kondeti, S.; Choi, Y.E. High frequency somatic embryogenesis and plant regeneration of interspecific ginseng hybrid between Panax ginseng and Panax quinquefolius. J. Ginseng Res. 2019, 43, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Martínez, T.; Corredoira, E.; Valladares, S.; Jorquera, L.; Vieitez, A.M. Germination and conversion of somatic embryos derived from mature Quercus robur trees: The effects of cold storage and thidiazuron. Plant Cell Tiss. Organ Cult. 2008, 95, 341–351. [Google Scholar] [CrossRef]
- Coleman, G.D.; Ernst, S.G. In vitro shoot regeneration of Populus deltoides: Effect of cytokinin and genotype. Plant Cell Rep. 1989, 8, 459–462. [Google Scholar] [CrossRef]
- Bhatia, P.; Ashwath, N.; Midmore, D.J. Effects of genotype, explant orientation, and wounding on shoot regeneration in tomato. In Vitro Cell Dev. Biol-Plant. 2005, 41, 457–464. [Google Scholar] [CrossRef]
- Griga, M. Direct somatic embryogenesis from shoot apical meristems of pea, and thidiazuron-induced high conversion rate of somatic embryos. Biol. Plant. 1998, 41, 481–495. [Google Scholar] [CrossRef]
- Wetzstein, H.Y.; Baker, C.M. The relationship between somatic embryo morphology and conversion in peanut (Arachis hypogaea L.). Plant Sci. 1993, 92, 81–89. [Google Scholar] [CrossRef]
- Malik, M.Q.; Mujib, A.; Gulzar, B.; Zafar, N.; Syeed, R.; Mamgain, J.; Ejaz, B. Genome size analysis of field grown and somatic embryo regenerated plants in Allium sativum L. J. Appl. Genet. 2020, 61, 25–35. [Google Scholar] [CrossRef]
- Syeed, R.; Mujib, A.; Malik, M.Q.; Mamgain, J.; Ejaz, B.; Gulzar, B.; Zafar, N. Mass propagation through direct and indirect organogenesis in three species of genus Zephyranthes and ploidy assessment of regenerants through flow cytometry. Mol. Biol. Rep. 2021, 48, 513–526. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Mean Number of Fruits/Individual Plant *** | Mean Weight of Fruit (g) *** | Mean Number of Seeds/Individual Plant *** |
|---|---|---|---|
| CP | 32.2 ± 10.2 b | 0.20 ± 0.05 c | 45.8 ± 16.9 b |
| CR | 16.5 ± 6.3 c | 0.28 ± 0.04 a | 23.1 ± 9.2 c |
| GP | 38.8 ± 10.4 a | 0.23 ± 0.04 b | 58.8 ± 16.6 a |
| YP | 37.6 ± 14.1 ab | 0.23 ± 0.04 b | 53.4 ± 20.8 ab |
| Cultivar | Mean Weight of Indehiscent Seed (mg) *** | Mean Diameter of Indehiscent Seed (mm) *** | Mean Length of Indehiscent Seed (mm) *** | Mean Width of Indehiscent Seed (mm) *** |
|---|---|---|---|---|
| CP | 23.2 ± 3.5 d | 1.82 ± 0.21 c | 4.69 ± 0.26 c | 3.62 ± 0.22 c |
| CR | 34.1 ± 3.4 a | 2.26 ± 0.16 a | 5.21 ± 0.25 a | 4.13 ± 0.23 a |
| GP | 27.6 ± 3.3 c | 2.17 ± 0.18 b | 4.94 ± 0.29 b | 3.72 ± 0.20 bc |
| YP | 31.0 ± 5.0 b | 2.27 ± 0.25 a | 4.84 ± 0.28 b | 3.79 ± 0.25 b |
| Cultivar | Mean Weight of Dehiscent Seed (mg) *** | Mean Diameter of Dehiscent Seed (mm) *** | Mean Length of Dehiscent Seed (mm) ** | Width of Dehiscent Seed (mm) *** |
|---|---|---|---|---|
| CP | 56.3 ± 7.1 c | 2.57 ± 0.23 c | 5.47 ± 0.31 ab | 4.66 ± 0.35 bc |
| CR | 74.4 ± 9.0 a | 2.99 ± 0.38 a | 5.58 ± 0.46 a | 4.90 ± 0.41 a |
| GP | 56.4 ± 5.8 c | 2.80 ± 0.29 b | 5.39 ± 0.39 b | 4.57 ± 0.33 c |
| YP | 67.7 ± 8.2 b | 2.97 ± 0.26 a | 5.58 ± 0.48 a | 4.75 ± 0.40 b |
| Cultivar | Somatic Embryo Induction Rate (%) ns | Mean Number of Somatic Embryos/Explant * | Mean Number of Somatic Embryos/Seed *** |
|---|---|---|---|
| CP | 99.3 ± 1.0 | 9.3 ± 1.3 b | 76.4 ± 22.3 b |
| CR | 100 ± 0.0 | 14.4 ± 3.7 a | 100.2 ± 23.3 a |
| GP | 100 ± 0.0 | 7.3 ± 0.6 b | 76.9 ± 24.8 b |
| YP | 100 ± 0.0 | 8.7 ± 2.1 b | 71.3 ± 20.2 b |
| Cultivar | Mean Number of Shoots/Seed * | Shoot Conversion Rate (%) ns |
|---|---|---|
| CP | 50.7 ± 2.2 b | 66.4 ± 1.9 |
| CR | 61.3 ± 4.3 a | 61.4 ± 6.0 |
| GP | 57.1 ± 4.8 ab | 75.4 ± 9.5 |
| YP | 52.0 ± 2.6 b | 73.6 ± 6.9 |
| Cultivar | Mean Number of IGRs/Seed ns | IGRs Conversion Rate (%) ns |
|---|---|---|
| CP | 39.6 ± 10.6 | 51.8 ± 13.4 |
| CR | 41.0 ± 4.1 | 41.6 ± 8.4 |
| GP | 41.1 ± 4.4 | 58.7 ± 12.9 |
| YP | 37.1 ± 2.0 | 49.5 ± 10.5 |
| Number of Transplanted IGRs | Number of Sprouted IGRs | Sprouting Rate (%) |
|---|---|---|
| 1200 | 1055 | 87.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-W.; Kwon, N.; Kim, J.-U.; Bang, K.-H.; Jung, S.M.; Lee, S.W.; Kim, D.-H.; Kim, Y.-C.; Jo, I.-H.; Park, Y.-D. In Vitro Micropropagation of Commercial Ginseng Cultivars (Panax ginseng Meyer) via Somatic Embryogenesis Compared to Traditional Seed Production. Horticulturae 2023, 9, 435. https://doi.org/10.3390/horticulturae9040435
Lee J-W, Kwon N, Kim J-U, Bang K-H, Jung SM, Lee SW, Kim D-H, Kim Y-C, Jo I-H, Park Y-D. In Vitro Micropropagation of Commercial Ginseng Cultivars (Panax ginseng Meyer) via Somatic Embryogenesis Compared to Traditional Seed Production. Horticulturae. 2023; 9(4):435. https://doi.org/10.3390/horticulturae9040435
Chicago/Turabian StyleLee, Jung-Woo, Nayeong Kwon, Jang-Uk Kim, Kyong-Hwan Bang, Sung Min Jung, Sung Woo Lee, Dong-Hwi Kim, Young-Chang Kim, Ick-Hyun Jo, and Young-Doo Park. 2023. "In Vitro Micropropagation of Commercial Ginseng Cultivars (Panax ginseng Meyer) via Somatic Embryogenesis Compared to Traditional Seed Production" Horticulturae 9, no. 4: 435. https://doi.org/10.3390/horticulturae9040435