Genome-Wide Identification and Expression Analysis of the fw2.2-like Gene Family in Pear

College of Horticulture, Xinjiang Agricultural University, Urumqi 830052, China
*
Authors to whom correspondence should be addressed.
Horticulturae 2023, 9(4), 429; https://doi.org/10.3390/horticulturae9040429
Submission received: 25 February 2023
/
Revised: 23 March 2023
/
Accepted: 24 March 2023
/
Published: 25 March 2023
(This article belongs to the Section Genetics, Genomics, Breeding, and Biotechnology (G2B2))
Abstract
:Fruit size is a major factor determining yield, quality, and consumer acceptability. fw2.2 (fruit weight-2.2) is a primary quantitative trait locus that was the first to be cloned, accounting for 30% of the variation in tomato fruit size. The various homologs of fw2.2 (fw2.2-like) have been identified in many plants and belong to a large family. To date, there has been no report that has carried out a comprehensive identification of fw2.2-like members in pear. In this study, a total of 14 fw2.2-like genes were identified in the pear (Pyrus bretschneideri Rehd) genome and designated as PbFWL1-14. All of the PbFWL genes were unevenly distributed on nine chromosomes, and each chromosome contained between one and four genes. All PbFWL proteins contained more than two conserved motifs, and PbFWL genes contained more than one intron, and the genes of the same subfamily seemed to have a similar intron gene structure. According to the neighbor-joining phylogenetic tree, a total of 78 fw2.2/CNR (cell number regulator) from five plant species, including pear, maize, tomato, peach, rice, and physalis, could be divided into seven subgroups, and PbFWL proteins were mainly distributed in subgroups 1, 3, 4, 5, and 6. The Ka/Ks analysis also revealed that the fw2.2-like gene family of pear may have been subjected to strong purifying selection pressure during its evolution. A cis-element analysis found that many cis-elements responsive to hormones and stress were discovered in promotion regions for all PbFWLs. When combining real-time quantitative PCR analysis detection results, PbFWL1/2/5 were found to be the most likely candidate genes for regulating pear fruit size.
1. Introduction
Fruit size is one major factor determining yield, quality, and consumer acceptability. Genes participate in fruit weight or size gain under suitable growth conditions by regulating cell number and size [1,2]. At present, some genes are known to affect plant and organ size by regulating cell numbers or size [3,4,5,6,7]. Tomato fruit-weight2.2 (fw2.2) was the first major quantitative trait locus (QTLs) to be cloned, and it regulates tomato fruit size and weight by controlling the carpel cell number, accounting for 30% of fruit size variation, with negative regulation of fruit size [8]. Alterations in fruit size, imparted by fw2.2 alleles, are due to a variation in gene expression and not due to a different sequence and structure of the encoded protein [3].
Various homologs of fw2.2 proteins can be found throughout the plant kingdom as well as in mammals (fw2.2-like, also referred to as CNR or FWL (fw2.2-like) in the literature) [9]. Thibivilliers et al. used fw2.2 for sequence homology comparison and found 134 FWL genes in 13 plant species. Most plant FWL proteins consist of 100 to 250 amino acids, some being greater than 400 amino acids, and amino acid sequence comparisons between fw2.2 protein homologs revealed that they all have cysteine-rich PLAC8 structural domains [10]. They have functions in regulating organ size, maintaining metal ion homeostasis in vivo, and controlling calcium ion uptake [11]. Among them, the function of regulating organ size is universal in plants. For example, the maize gene cell number regulator1 (Zea mays; ZmCNR1) negatively controls the size of the whole plant [9]. The biological function of other fw2.2-like genes that negatively regulate fruit development has also been described in husk tomato (Physalis floridana; PfCNR1) [12], sweet cherry (Prunus avium; PavCNR12 and PavCNR20) [13], avocado (Persea americana; Pafw2.2-like) [14], and loquat (Eriobotrya japonica; EjFWL1 and EjFWL2) [15]. The homologs of fw2.2 seem to belong to a large family of the given organisms. The fw2.2-like gene family has been studied in a variety of plants, including arabidopsis [9], oilseed rape [16], rice [17], maize [9], and pomegranate [18].
Pear, belonging to the genus Pyrus in the Rosaceae family, is one of the most important fresh fruits in the world; at least 22 different species are maintained worldwide [19]. These accessions have abundant fruit size variability, and the weight of a single fruit ranges from a few grams to several hundred grams [20]. In the production and cultivation of fruit trees, the size of the fruit is the main determinant of fruit grading and the economic efficiency of the orchard. For example, the Korla fragrant pear (Pyrus sinkiangensis Yü) is a well-received variety growing in the Bayingolin Mongol Autonomous Prefecture and the Aksu region in China. The Korla fragrant pear cultivate with a single fruit weight of 120 g is 3~4 times more expensive than the Korla fragrant pear cultivar with a single fruit weight of 80 g [21]. Therefore, breeding new large-fruited cultivars is a very major breeding objective. However, the long juvenile period of pears, which can last 5 years or even longer, greatly hampers the rapid phenotypic evaluation of fruit quality traits. Therefore, understanding the markers and genes associated with pear fruit size will greatly improve the efficiency of breeding large-fruited cultivars, as it would allow for the early elimination of seedlings that are likely to produce fruit that does not meet the breeding target. Pear fruit weight is a quantitative trait, like weight in tomatoes. Many QTL loci associated with fruit size have been detected in pear, and these loci are mostly concentrated in the LG7 and LG11 linkage groups [22,23]. The fw2.2-like gene family provides a good source of candidate genes for studying the molecular mechanism of pear fruit size. However, the fw2.2-like gene family genome-wide identification has not been reported in pear. In this study, we carried out a genome-wide analysis of Chinese white pear (Pyrus bretschneideri) fw2.2-like genes and selected ‘Duli’ pear (small-fruited), Korla fragrant pear (medium-fruited), ‘Zaomeixiang’ pear (medium-fruited, which is the large-fruited bud varietal of Korla fragrant pear), and ‘Yali’ pear (large-fruited) as research materials, focusing on the two most vigorous cell division periods, those of flowering and young fruit expansion, to explore the pear fw2.2-like gene family and investigated their potential functions in controlling fruit size.
2. Materials and Methods
2.1. Identification of the fw2.2-like Family in Pear
The protein sequence data of Chinese white pear (Pyrus bretschneideri) are available in the pear genome project (http://peargenome.njau.edu.cn/ accessed on 4 January 2022) [20]. We used a combination of two methods to search for members of the fw2.2-like family in Pyrus bretschneideri: an HMM search with the PLAC8 domain HMM profile (PF04749) and BLASTp searches, each using fw2.2 (Solyc02G090730.2.1) protein sequences from tomato as queries. First, the HMM profiles of PLAC8 were retained from the Pfam database (http://pfam.xfam.org/ accessed on 4 January 2022) and were used to identify the putative fw2.2-like proteins with the best domain E-value cutoffs of ≤1 × 10−4 [18]. The tomato fw2.2 sequences were used as the query with which to perform a BLAST search in Pyrus bretschneideri, with a cutoff E-value of ≤1 × 10−5. To validate the HMM and BLAST search, these potential sequences were submitted to NCBI Batch CD-Search tools (CDD) [24], Pfam [25], and SMART (http://smart.embl-heidelberg.de/ accessed on 7 March 2022) in order to confirm the conserved PLAC8 domain. The predicted protein sequences lacking the PLAC8 domain or containing other structures were excluded. All reliable fw2.2-like sequences were uploaded to ExPASy [26] in order to calculate the number of amino acids, isoelectric points (pI), and molecular weights. The subcellular localization of the identified PLAC8 proteins was determined using Plant-mPLoc [27]. Multiple sequence alignment was performed using ClustalX (http://www.ebi.ac.uk accessed on 20 March 2022).
2.2. Analysis of fw2.2-like Gene Structure and Conserved Motifs
Sequences of pear genome-wide fw2.2-like gene family members were analyzed and structures were mapped using the online program GSDS (http://gsds.cbi.pku.edu.cn/ accessed on 22 March 2022). Conserved motifs in the pear PbFWLs protein were identified using the online software MEME 5.5.1 (http://meme-suite.org/ accessed on 23 March 2022) with the following optimized parameters: number of repeats, arbitrary; maximum number of motifs, 6; and optimal width of each motif, between 6 and 100 residues.
2.3. Analysis of Cis-Acting Elements in fw2.2-like Gene Promoters
The upstream sequence (1.5 kb) of the coding sequence of the fw2.2-like gene was retrieved from the Chinese white pear genomic data and then submitted to PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ accessed on 26 March 2022) for analysis of promoter sequences. The cis-acting element distribution was mapped using GSDS2.0 (http://gsds.cbi.pku.edu.cn accessed on 26 March 2022) [28].
2.4. Chromosomal Localization and Gene Duplication
The localization data of fw2.2-like pears were also obtained from genome annotation files (https://www.rosaceae.org/species/pyrus_betulifolia/genome_v1.0 accessed on 2 April 2022) [20]. Chromosome localization was mapped using MapChart 2.2 software [29]. In addition, gene duplication maps were obtained using MCScanX [30]. Gene duplications were confirmed using two criteria: (i) if shorter paired sequences covered more than 70% of longer sequences; (ii) if there was a similarity of paired sequences >70% [31,32]. Two genes located in the same chromosomal segment at less than 100 kb and separated by five or fewer genes were identified as tandem duplicates [33]. The differentiation time was calculated using the formula T = Ks/2r, where Ks is the synonymous mutation at each locus and r is the rate of differentiation of the plant nuclear genes. For dicotyledons, r was considered to be 1.5 × 10−8 synonymous mutations per locus per year [34].
2.5. Phylogenetic Analysis of fw2.2-like Proteins
PbFWLs were aligned with fw2.2/CNR proteins from other plants, such as those of Zea mays (12 sequences) [9], tomato (19 sequences and 1 sequence from wild tomato) [13,35], peach (23 sequences) [13], rice (8 sequences) [17], and physalis (1 sequence) [12], and multiple sequence comparisons were performed using ClustalW. Phylogenetic trees were constructed using comparisons from MEGA 7.0 [36] with the maximum likelihood method and a bootstrap set to 1000. The parameters were referenced in Guo et al. [9].
2.6. Plant Material
The three pear cultivars (Korla fragrant pear, ‘Zaomeixiang’ pear, and ‘Yali’ pear) and one wild pear (‘Duli’ pear) which were used in this study are listed in Supplementary Table S1. The 25-year-old trees were selected from the pear germplasm collection at the Research Center of Xinjiang Korla Fragrant Pear (Korla, China). These cultivated pear trees, grafted onto ‘Duli’ pear rootstocks, were spaced 2.5 × 4 m apart. They received annual routine horticultural care, were hand-pollinated with ‘Dangshan Su’ pollen at the anthesis on branches in the middle periphery of the canopy, and tagged.
2.7. Measuring the Characteristics of Fruit at Early Stages of Pear Fruit Development
Material at the anthesis (−4 and 0 days after full bloom, −4 and 0 DAFB) was collected randomly at the periphery of the tree in the middle of the canopy on untagged branches. A total of 10 flowers were collected at a time; all parts above the ovary (including stigma, stamens, and petals) and the stalk were removed to obtain the receptacle. Fruiting samples were collected starting with 10 DAFB, and fruit marked by pre-tagging was collected every 10 days until 50 DAFB (fruit cell division was expected to end around 50 DAFB), at a rate of 10 fruits at a time, with sepals and stalk parts removed. The longitudinal diameter of the receptacle (fruit) was measured using vernier calipers, the fresh weight of the receptacle (fruit) was weighed on a thousandth of a scale, and the receptacle (fruit) volume was calculated according to the longitudinal diameter method V = 4/3πd3 (V is the receptacle or fruit volume and d is the average of 1/2 receptacle or the fruit’s longitudinal diameter) [37]. This was repeated 3 times.
2.8. Measurements of Receptacle/Fruit Cell Size and Number
Pear fruits developed from receptacles, and a previous study showed that the receptacle develops into the pulp of pear, and the size difference between cultivars is mainly due to the number of mesocarp cells [38,39]. Thus, 10 tissues (receptacle equatorial axes and mesocarp tissue) per cultivar were preserved in an FAA solution (70% ethanol: acetic acid: formalin = 90:5:5) for paraffin sectioning. The tissues fixed in an FAA fixative solution were dehydrated, cleared, and embedded. Fruit sections were produced with a rotary microtome (ERM 3100, Heston, Jiangsu, China), and the sections were stained with the safranin and fast green counterstain method. The stained surfaces were observed under a Nikon Eclipse 80i microscope (Nikon, Tokyo, Japan) and photographed. The measurement of cell size and cell number in the receptacle/mesocarp was conducted according to the specifications of Tian et al. [40]. We used the following formula to calculate the cell volume: V = 4/3πr3 (V represents the cell volume, r represents the average value of the vertical and horizontal diameter of 1/2 cells); cell number = fruit volume/cell volume [37]. Relative cell proliferation rate = (cell number 2—cell number 1)/(time 2—time 1), −4 to 0 DAFB for the first period P1 (period 1), and so on, 0 to 10 DAFB, and 10 to 20 DAFB for P2 and P3, respectively.
2.9. RNA Isolation and qRT-PCR
The receptacle and mesocarp tissue RNA was extracted from Korla fragrant pear, ‘Zaomeixiang’ pear, ‘Yali’ pear, and ‘Duli’ pear on −4 DAFB, 0 DAFB, 10 DAFB, 20 DAFB, 30 DAFB, and 40 DAFB. The receptacle (fruit) was cut crosswise along the equatorial side of the receptacle (fruit) with a scalpel, and the ovary (mesocarp) tissue was taken in the middle of the crosswise section and placed in an ultra-low-temperature refrigerator at −80 °C to freeze and for preservation. Total RNA was extracted using a polysaccharide polyphenol plant total RNA extraction kit (TIANGEND, Beijing, China). The cDNA was synthesized by reverse transcription according to the instructions in the FastKing RT Kit (with gDNase) (TIANGEND, Beijing, China). Based on the obtained cDNA sequence of the pear fw2.2-like gene, qRT-PCR primers were designed and examined using Primer 5.0 software, using the pear TUB2 gene as an internal reference; all primer information is listed in Supplementary Table S2. The expression of each internal reference gene was detected using the BIO-RAD CFX Connect™ Fluorescent Quantitative PCR Assay System. The reagent used was Power SYBR® Green PCR Master Mix (Applied Biosystems Inc., MA, USA, Cat. No. 4367659), and the PCR programs used were 95 °C, 3 min pre-denaturation; 95 °C, 10 s denaturation; 55 °C, 20 s annealing; 72 °C, 20 s extension, with 40 cycles in total. Each sample was repeated 3 times. The relative expression of genes was calculated using 2−ΔΔCt.
3. Results
3.1. Identification and Sequence Analysis of fw2.2-likes in Pear
BLASTp and HMM searches were conducted to extensively identify pear fw2.2-likes using tomato fw2.2 protein sequences as queries. Thirty-six fw2.2-like genes were obtained in pear; then, the fw2.2-like protein sequences encoded by nonrepresentative transcripts were excluded. The remaining sequences were checked using CDD, Pfam, and SMART to determine if the remaining sequences had the full PLAC8 structural domain. In total, 14 sequences were confirmed as pear fw2.2-like proteins and were thus named PbFWL1 to PbFWL14. Gene names, gene IDs, chromosomal locations, gene location, amino acid numbers, molecular weights, isoelectric point (pI), instability index, and grand average of hydropathicity (GRAVY) are listed in Table 1. The predicted subcellular localization was the cell membrane and nucleus for PbFWL8 and PbFWL9 and the cell membrane for the other fw2.2-likes. The protein length of the pear fw2.2-like family gene members ranged from 151 (PbFWL1) to 415 amino acids (PbFWL9). The molecular weight of the pear fw2.2-like family genes ranged between 16.99 kDa (PbFWL1) and 47.51 kDa (PbFWL9). The fw2.2-like family genes of pear were distributed on nine chromosomes. The predicted pI values of PbFWL proteins ranged from 4.79 (PbFWL5) to 7.79 (PbFWL6). Their instability index was greater than 40. The hydrophilic index was between −0.484 and 0.239, and the cysteine residue content was between 15 and 20, which was higher than the average protein.
A comparison of the protein sequences of PbFWLs, ZmCNR1, OsFWL1, OsFWL3, OsFWL4, PfCNR1, and fw2.2 (Figure 1) showed that they all have PLAC8 structural domains and belong to the PLAC8 supergene family. In the PLAC8 domain, all homologs and some amino acids in between seem to be highly conserved. These amino acids are composed of two common protein motifs (CXXXXXCPC and QEYRELK), but the CXXXXXCPC motif was not observed in PbFWL8.
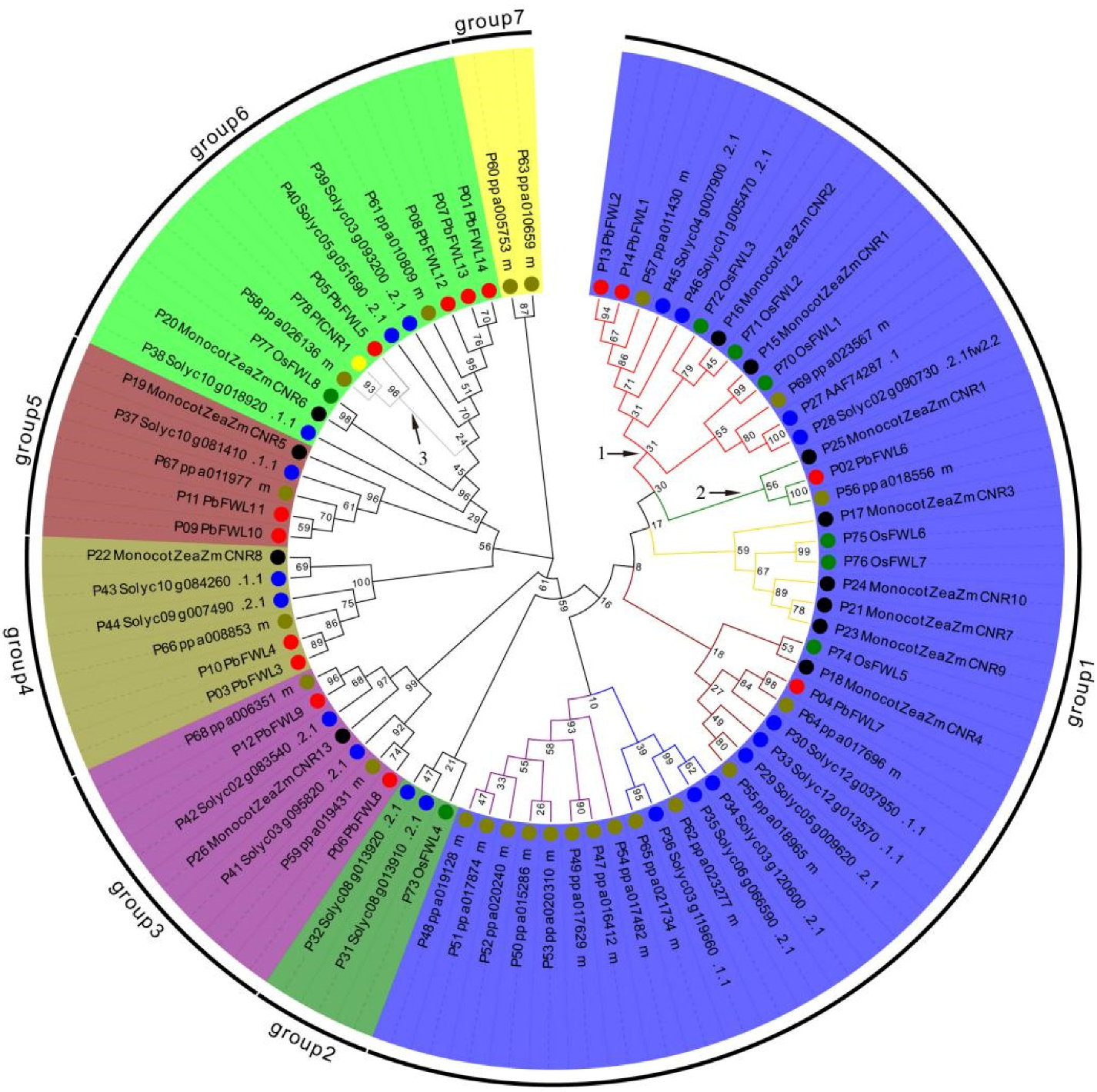
3.2. Phylogenetic Analyses of PbFWL Gene Family Members
To clarify the affinities and evolutionary distances between the pear fw2.2-like protein family and fw2.2-like proteins of other species, the full-length amino acid sequences of reported maize (12 members) [9], tomato (20 members) [13,35], peach (23 members) [13], rice (8 members) [8], physalis (1 member) [12] and pear fw2.2-like gene family members (14 members) were used in MEGA7.0 software to construct a phylogenetic tree (Figure 2). The protein sequences of fw2.2-like genes of tomato, Zea mays, peach, rice, and physalis were obtained from a previous study. Phylogenetic analysis showed that the 78 fw2.2/CNRs clustered into 7 distinct groups. In the clockwise direction, each group number is shown from 1~7. fw2.2/CNR members from the same species tended to cluster together in different phylogenetic tree groups. Group 1 contained 61.5% fw2.2/CNR proteins, labeled clade 1 in group 1 containing fw2.2, ZmCNR1, OsFWL1, and OsFWL3; this clade contained just two pear proteins, PbFWL1 and PbFWL2. PbFWL6 is a member of an adjacent subclade, labeled as 2 in group 1. A global pairwise amino acid comparison between fw2.2 and PbFWL1 showed 50.9% identity, and PbFWL2 showed 44.23% identity. fw2.2 was slightly more similar to PbFWL6 at 46.63%, with PbFWL2 and PbFWL6 showing 43.33% similar identity. However, the maximum likelihood test results indicate that PbFWL2 is closer to fw2.2 than PbFWL6. This closer proximity of PbFWL2 to fw2.2 was also revealed by three other family relationship algorithms, maximum parsimony, minimum evolution, and UPGMA cluster analysis, each completed using bootstrap tests. Group 6 contains 15.4% fw2.2/CNR proteins, labeled clade 3 in group 6, containing PfCNR1 and PbFWL5; a global pairwise amino acid comparison of PfCNR1 and PbFWL5 showed 85.1% identity.
3.3. Gene Structure and Motif Analysis
To understand the structural components of the PbFWL genes, we extracted the exon–intron information of 14 PbFWL genes from the pear database using in-house scripts. The examined genes between CDSs contained 3 to 7 exons and 2 to 6 introns (Figure 3B). PbFWL6 had 3 exons and 2 introns, and the examined genes had the smallest exon/intron number among all identified PbFWL genes. PbFWL9 had 7 exons and 6 introns, which represented the largest exon/intron number among the examined PbFWL genes. Exon/intron numbers varied among the homoeologous genes of the same family member but were essentially the same within the same subfamily (cluster I: exon: 4 to 5/intron: 3 to 4, cluster II: exon: 3/intron 2, cluster III: exon: 3/intron: 2, cluster IV: exon: 6 to 7/intron: 5 to 6, cluster V: exon: 3 to 4/intron: 2 to 3). Furthermore, all genes except PbFWL6 and PbFWL7 contained untranslated regions (UTR).
The MEME website identified the conserved motifs of PbFWL proteins, revealing six in total. The length of these conserved motifs ranged from 15 to 50 amino acids (Supplementary Table S3). Using Pfam, CDD, and SMART analysis, motifs 1, 2, 3, 4, and 6 were found to correspond exactly to the regions of the PLAC8 domain (Figure 3C). We also identified motifs unique to each cluster. These included motifs 3, 5, and 6, which were only present in cluster I; motif 1 was missing in cluster III. The only exceptions were that motif 4 was missing in PbFWL8 (cluster IV). According to the analyses of conserved motifs and gene structures, although the number of conserved motifs and the length of exons and introns varied to some extent, the conserved motifs and gene structures of the same subfamily members were highly consistent.
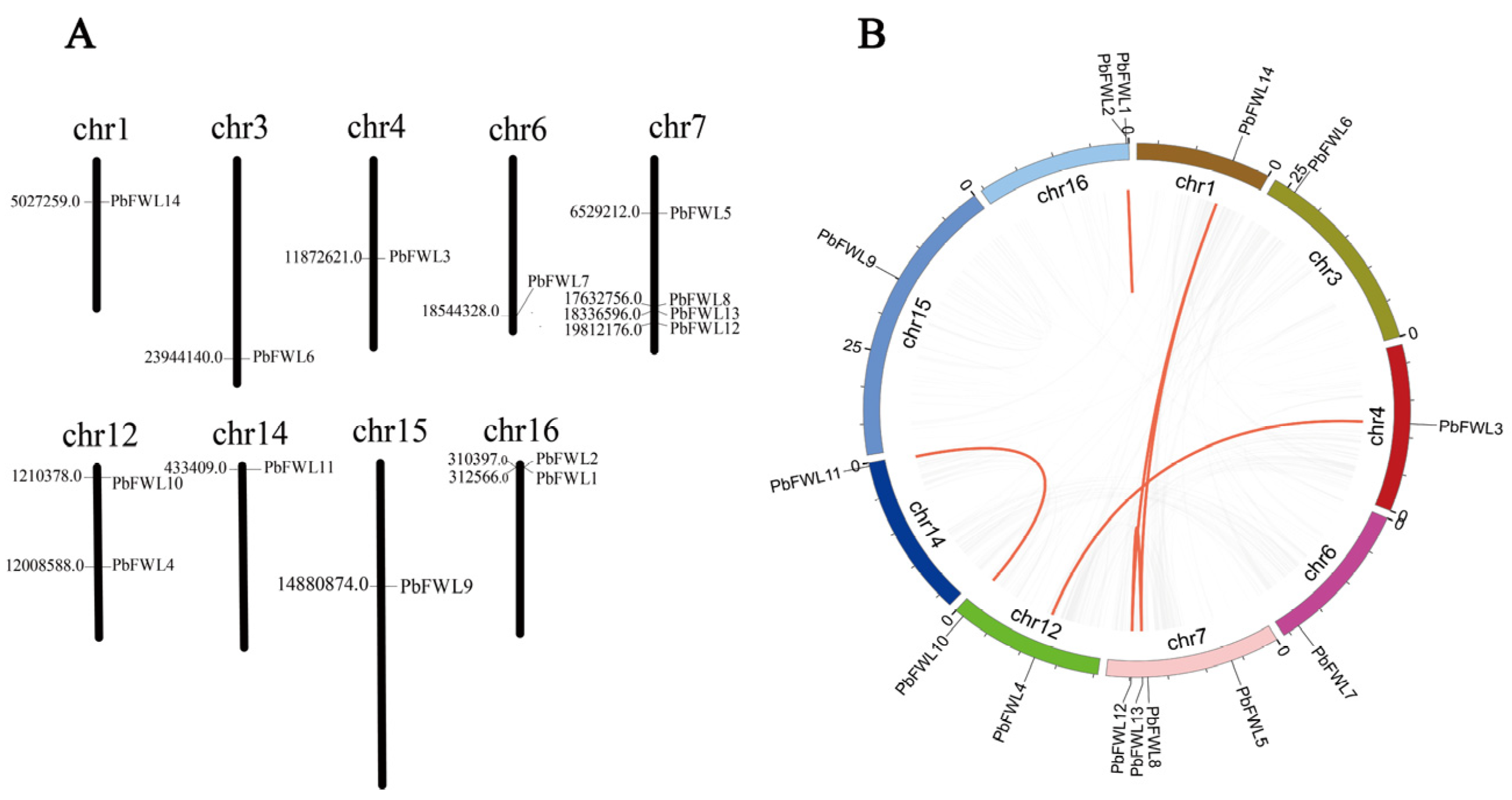
3.4. Chromosomal Distribution and Synteny Analysis of PbFWLs in Pyrus Bretschneideri
To further study the relationship between the genetic evolution within the fw2.2-like gene family in pear, we analyzed the chromosomal location and synteny of the PbFWL genes. The physical mapping of PbFWLs on the chromosomes of Pyrus bretschneideri showed that the distribution of PbFWLs was unequal; these genes were distributed on nine chromosomes, with one or more genes per chromosome (Figure 4A). One pair of genes (PbFWL1/PbFWL2) was repeated in tandem, and five pairs (PbFWL10/PbFWL11, PbFWL13/PbFWL14, PbFWL12/PbFWL14, PbFWL12/PbFWL13, PbFWL3/PbFWL4) were segmentally duplicated (Figure 4B). To better understand the drivers of pear fw2.2-like gene evolution, Ka/Ks values of gene duplication pairs were calculated. The results showed that the Ka/Ks values of all gene duplication pairs were <1, indicating that the pear fw2.2-like gene family may have undergone purifying selection during evolution. The Ks values ranged from 0.02 to 0.231, indicating that the earliest duplication occurred 7.7 million years ago and that the latest duplication occurred 0.667 million years ago (Supplementary Table S4).
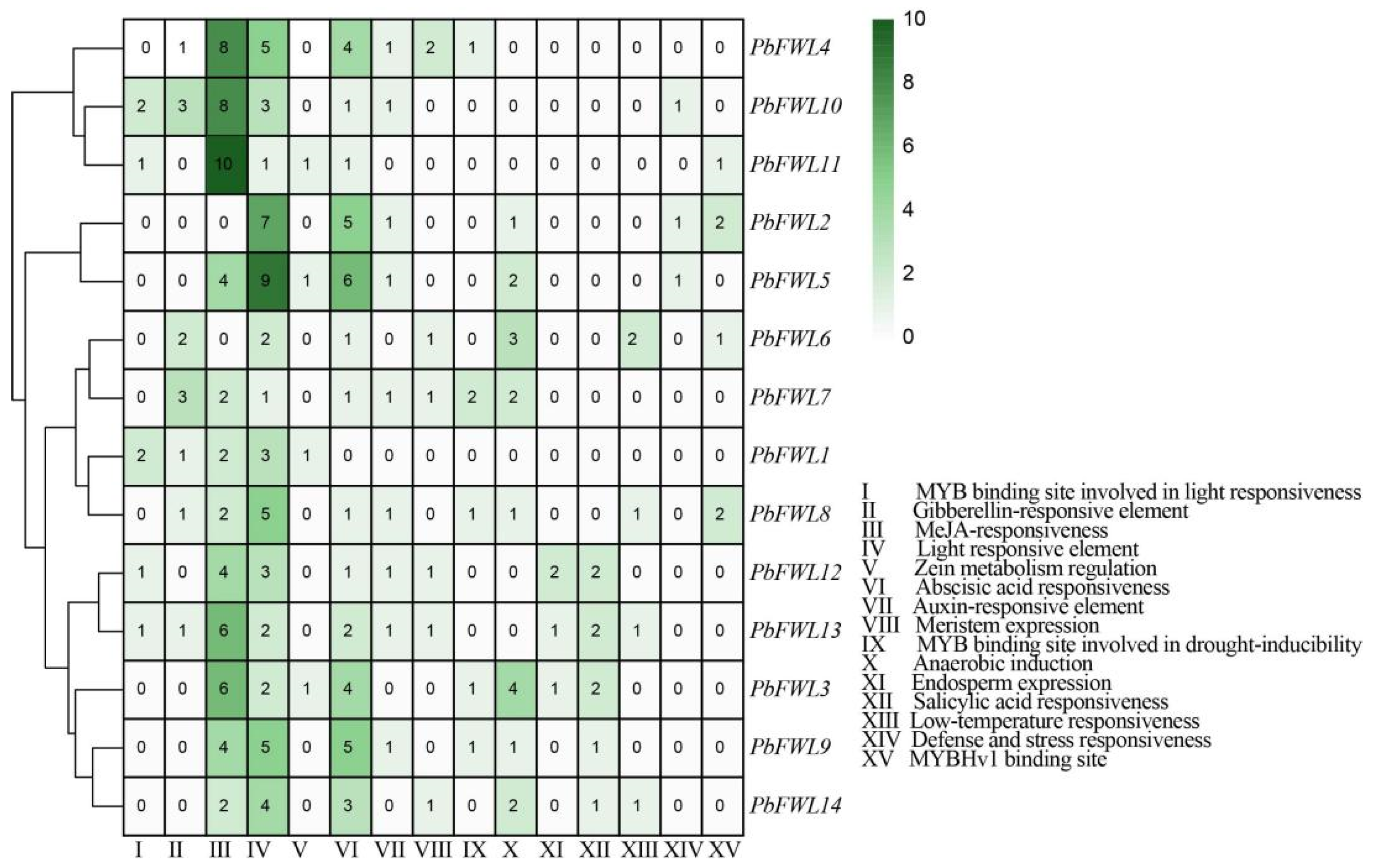
3.5. Analysis of the fw2.2-like Gene Family Promoter Elements in Pear
Cis-acting elements are important cues for predicting gene function. Thus, in order to understand the potential role of the fw2.2-like gene family in pear growth and development, we analyzed the promoter cis-acting elements of the pear fw2.2-like gene family. As a result, we found many cis-acting elements in the initiation of pear fw2.2-like gene family genes, and 15 of them were selected as interesting cis-acting elements for further analysis, specifically those related to stress, hormones, plant growth, and development (Figure 5). Additionally, we found that all PbFWL genes contain cis-acting elements related to hormone regulation, such as gibberellin (GA), methyl jasmonate (MeJA), abscisic acid (ABA), auxin, and salicylic-acid-responsive elements.
3.6. Differential Expression of the PbFWL Genes during Early Fruit Development among Pear Varieties with Different Fruit Sizes
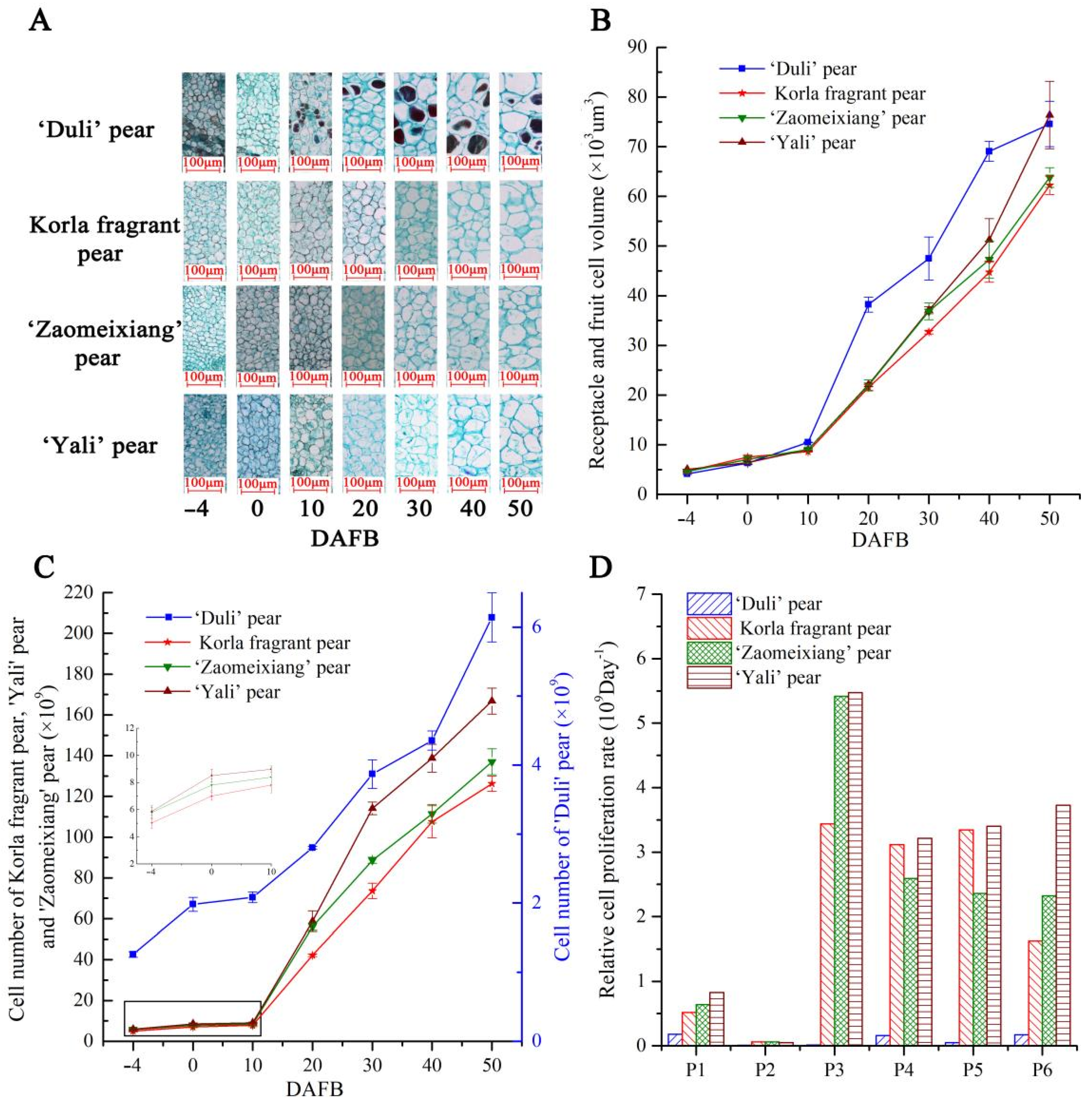
To determine whether PbFWLs are involved in the regulation of receptacle and fruit development early in fruit development, we analyzed the correlation of PbFWL gene expression with cell division related to the receptacle and flesh using large-fruited (‘Yali’ pear), medium-fruited (Korla fragrant pear, ‘Zaomeixiang’ pear), and small-fruited wild cultivars (‘Duli’ pear) during early fruit development stages. There were apparent differences in the receptacle and fruit size between the ‘Duli’ pear and the other three pear cultivars (Supplementary Figure S1). The observation of cells in the receptacle and early fruit showed that cells in the preliminary blooming stage (−4 and 0 DAFB) were of similar size, and that the cell size of the small-fruited wild cultivar was greater than that of the other cultivars at 10 to 40 DAFB (Figure 6A,B). In general, cell numbers in the same periods were higher for the ‘Yali’ pear than the ‘Zaomeixiang’ pear, followed by the Korla fragrant pear and the ‘Duli’ pear (Figure 6C). Analysis of the relative cell proliferation rates during the adjacent time periods revealed that the receptacle cells had two cell proliferation phases: a first phase before 0 DAFB (P1), when the cell proliferation occurs slowly, and a second phase from 10 DAFB to 50 DAFB (P3 to P5), when cells proliferated rapidly (Figure 6D).
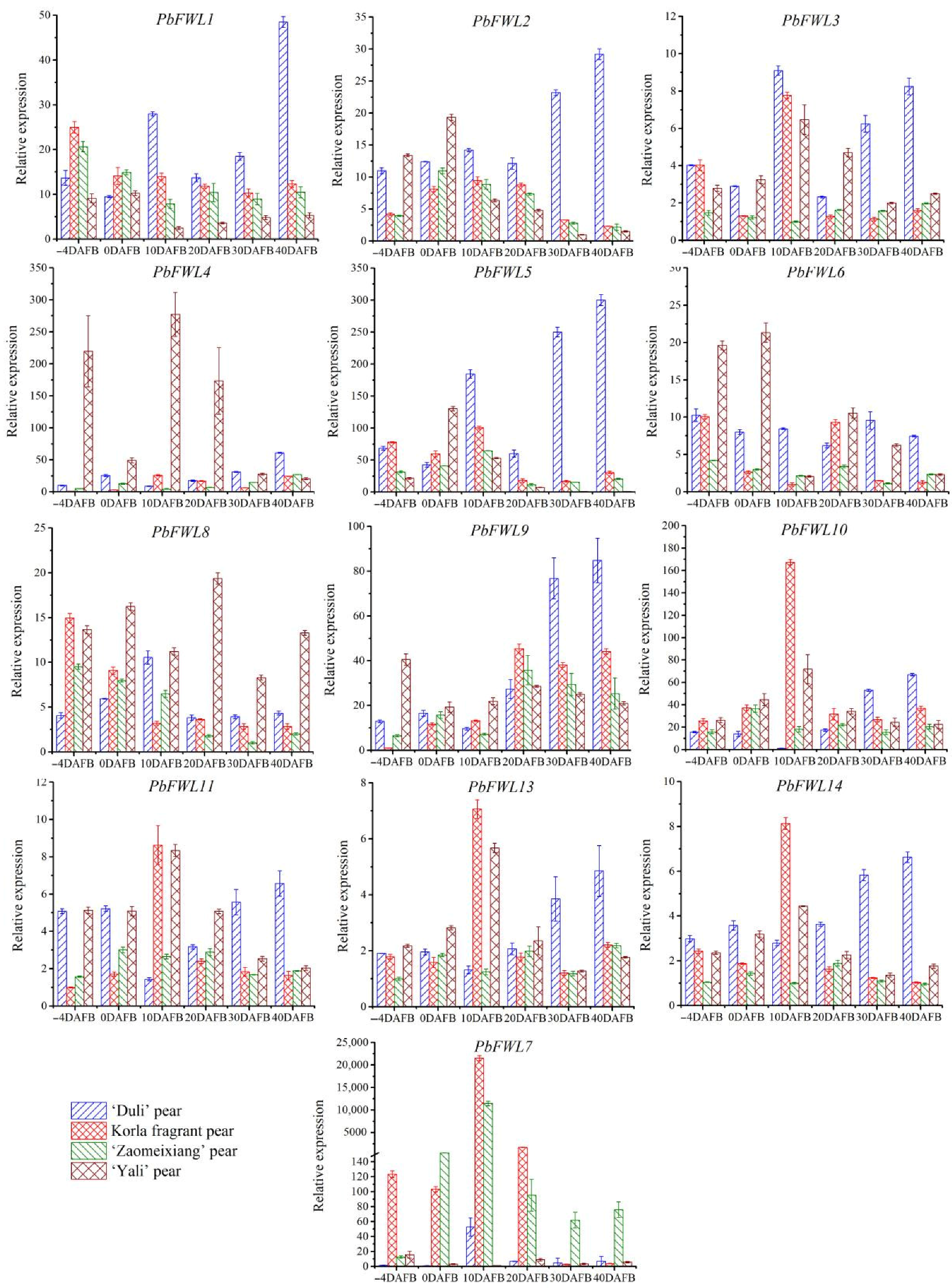
The fw2.2 gene negatively regulates tomato fruit size and weight by affecting carpel cell division. Based on the results of pear receptacle (fruit) tissue slices in this experiment (Figure 6), we speculated that the accumulation level of PbFWLs transcripts in small-fruited wild cultivar (‘Duli’ pear) is larger than that in medium-fruited cultivars (Korla fragrant pear and ‘Zaomeixiang’ pear) followed by large-fruited cultivar (‘Yali’ pear). To verify this conclusion, we assessed the gene expression levels of 14 selected PbFWL genes using qRT-PCR of the receptacle or pulp tissues at different developmental stages (−4, 0, 10, 20, 30, and 40 DAFB) in the ‘Duli’ pear, Korla fragrant pear, ‘Zaomeixiang’ pear, and ‘Yali’ pear. Since the sequences of PbFWL12 and PbFWL13 are highly similar, it is impossible to design primers to separate the two sequences. Finally, the expression levels of 13 PbFWL genes were monitored (Figure 7). Gene expression data showed that 13 PbFWL genes did not exhibit regular expression patterns in the receptacle. Except for PbFWL1/2/5, the remaining PbFWL genes also did not exhibit regular expression patterns in the pulp. Lower PbFWL1/2/5 transcription levels were found in the fruits at the fruit cell division phase (10 to 40 DADB) in the large-fruited cultivar (‘Yali’ pear) compared to those levels in the same phase in medium-fruited cultivars (Korla fragrant pear and ‘Zaomeixiang’ pear), and medium-fruited cultivars levels were lower than small-fruited wild cultivar levels (‘Duli’ pear). The transcript levels of PbFWL1/2/5 were the highest at the 40 DAFB stage of small-fruited wild cultivars and the 10 DAFB stage of medium and large fruits. The above description of correlation was an artificial judgment and lacks statistical support, and so we quantified the division of pulp cells with gene expressions falling between 10 to 40 DAFB for correlation analysis (Table 2). The results showed that between 10 to 40 DAFB, the expression of PbFWL1/2/5 correlated with the division of pulp cells during this period, that of PbFWL1 was significantly negatively correlated, while that of PbFWL2 and PbFWL5 were highly significantly negatively correlated.
4. Discussion
The fw2.2-like gene family and the broader family of proteins containing the PLAC8 structural domain are key components in the regulation of cell division and organ growth [41]. fw2.2-like gene family members have been isolated and identified in multiple species, but the number of fw2.2-like gene family members differs across different species [42]. In this study, we described the identification of 14 homologous proteins to fw2.2 in pear, which were named the PbFWL genes. The number of fw2.2-like family gene members in maize was similar (12) [9] and less than that in tomato (19) [43] and peach (23) [13]. Although the number of fw2.2-like gene members in pear is the same as that in maize, the genome size of pear is 527 Mb [9], which is much smaller than that of maize at 2300 Mb [44]. Thus, the number of fw2.2-like genes in the genome does not depend entirely on the genome size. In plants, the fw2.2-like/CNR gene family members display short gene sequences. The length of amino acids in pear PbFWL genes ranged from 151 aa to 415 aa, which is very similar to that of maize (157 aa to 429 aa) [9]. An analysis of the isolated sequences revealed that the Cys-rich amino acids constitute the characteristic PLAC8 domain (PF04749) [45]. In the PLAC8 domain, there are two highly conserved motifs, CCXXXXCPC and QEYRELK. The function of QEYRELK is unknown [43,46].
In tomato, fw2.2 gene family members are located on the cell membrane [43]. However, in this paper, the pear fw2.2-like gene family members are not entirely located on the cell membrane. This result is consistent with that of rice [17]. PbFWL8 and PbFWL9 are predicted to be both membrane proteins and nuclear proteins, and this double-localization of proteins is common [47]. However, software predictions can be inaccurate in studies of the exact location of family proteins in cells. Further experiments, such as transient gene expression assays, are needed. The PbFWL proteins’ instability index was greater than 40, and the hydrophilicity index was between −0.484 and 0.134, indicating that putative pear PbFWLs are unstable hydraulic proteins. This result is consistent with that of FW in pomegranate [18]. The evolutionary tree shows that the members of tomato fw2.2 gene families are mainly located in group 1, which reflects that the evolution direction of tomato fw2.2 family genes may have a clear bias—that is, they evolve in the order of group 1. The pear fw2.2-like gene family members have four members in group 1 and group 6, indicating that the members of the pear fw2.2-like family are mainly evolved in the direction of group 1 and group 6. Members in groups 1 and 6 may play a more important role in the pear. Guo et al. studied the maize fw2.2-like gene family to investigate how the fw2.2-like gene affects maize plant or organ size, and phylogenetic and protein analyses revealed that ZmCNR1 is closest to fw2.2 and that they are structurally similar; for these reasons, ZmCNR1 was considered to be the best possible candidate maize direct line for the tomato fw2.2 homologs. Therefore, ZmCNR1 was selected for functional validation of the transgene [9]. In our study, the fw2.2 and ZmCNR1 specific subclades to which they belong, labeled clade 1 in Figure 3, contain just two pear proteins, PbFWL1 and PbFWL2. In addition, PfCNR1 and PbFWL5 specific subclades were labeled as clade 3. We speculate that PbFWL1, PbFWL2, and PbFWL5 may be involved in regulating cell division in organs such as pear fruits.
In PbFWLs, we found 6 tandem repeat genes, accounting for 42.8% of the total family members. Among them, three tandem repeat genes were distributed in group 6, which indicated that tandem repeats played essential roles in amplifying members in this group. Paralogous pairs with a Ka/Ks ratio <1 in pear, PbFWL genes indicate mutation restriction with purifying selection, similar to what occurs in pomegranate FWs [18]. Additionally, cis-elements have an important influence on the expression of downstream genes. In our study, the promoter regions of multiple family members had cis-elements that respond to ABA, IAA, and MeJA signals. The vital role of ABA in response to drought and other abiotic stresses has been confirmed [48]. MeJA is an essential signaling molecule that regulates plant resistance to biotic stress [49]. Therefore, it is speculated that PbFWL genes may be involved in the stress response of pears to biotic or abiotic stresses.
Tomato fw2.2 was the first cloned quantitative trait gene for of control fruit size in plants, accounting for 30% of the fruit weight variation. The transcript levels of small-fruited wild tomatoes are higher than those of large-fruited cultivated tomatoes, regulating the number of carpel cells and affecting fruit size [50,51]. The function of FWL in controlling cell proliferation is universal in plants [42]. For example, the expression of the avocado PaFWL in small-fruit pulp tissue is higher than that in normal pulp tissue [14]; PfCNR1 affects the cell cycle and thus controls the size of berries and seeds in physalis [12]. Previous studies have shown that, during the cell division of pear fruit, PbFWL1 and PbFWL2 are more highly expressed in small fruits than in large fruits, with expression being negatively correlated with cell division in pear fruit [40]. This study found that the expression levels of 13 member genes of the pear fw2.2-like gene family do not correlate with the division of torus cells. Only the expression levels of PbFWL1, PbFWL2, and PbFWL5 from 10 to 40 DAFB were higher in small fruit than in medium fruit, followed by large fruit, with expression negatively correlated with pear fruit cell division. The expression levels of PbFWL1, PbFWL2, and PbFWL5 in the pulp of the large and medium fruits reached their peaks during the 10 DAFB period. In summary, we speculate that the functions of PbFWL1/2/5 genes are similar to those of fw2.2 and PfCNR1; the difference is that the pear fw2.2-like gene regulates the division of pulp cells in the young fruit stage of pear, affecting the size of pear fruits. PbFWL1/2/5, as membrane proteins, may interact with other genes to affect pear fruit size directly or indirectly. In addition, Arabidopsis AtPCR1 and rice OsFWL4 endow plants with cadmium resistance [17,52]. Therefore, PbFWLs may have other functions, which need to be studied in the future.
5. Conclusions
Fw2.2 is a primary quantitative trait locus that controls fruit size. The various homologs of fw2.2 (fw2.2-like) have been identified in many plants and constitute a large family found in their plant inner organisms. In this study, we identified 14 PbFWLs from the genome of Pyrus bretschneideri, and the biological information about fw2.2-like family members was analyzed. The RT-qPCR results showed that PbFW1/2/5 regulated fruit size at the young fruit stage and was the transcription factor regulating pear fruit size variation, while the expression of other PbFWLs was poorly correlated with fruit size. This study provides a good foundation for our next study on the molecular mechanism of how the pear fw2.2-like gene regulates fruit size.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9040429/s1, Figure S1: Fruit size variation in pears; Table S1: Plant materials used in this study and their characteristic fruit size and maturation period; Table S2: Sequence of primers used in the experiment; Table S3: List of the putative motifs of PbFWL proteins; Table S4: Ka/Ks value of homologous PbFWL gene pairs.
Author Contributions
X.P.: analysis of data, conducting the experiment, writing—original draft, writing—review and editing. J.T.: funding acquisition, conceptualization, project administration; supervision, writing—review and editing. J.L.: writing—review and editing; Y.W.: methodology, project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31760561) and the Xinjiang Uygur Autonomous Region Horticulture Key Discipline Fund (2016-10758-3).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data generated or analyzed during this study are included in this published article.
Acknowledgments
Thanks are given to the University of Xinjiang Agricultural for supporting this project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gillaspy, G.; Gruissem, B.D. Fruits: A develomental perspective. Plant Cell. 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monforte, A.J.; Díaz, A.; Caño-Delgado, A.; van der Knaap, E. The genetic basis of fruit morphology in horticultural crops: Lessons from tomato and melon. J. Exp. Bot. 2013, 65, 4625–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frary, A.; Nesbitt, T.C.; Frary, A.; Grandillo, S.; van der Knaap, E.; Cong, B.; Liu, J.; Meller, J.; Elber, R.; Alpert, K.B.; et al. fw2.2: A quantitative trait locus key to the evolution of tomato fruit size. Science 2000, 289, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, N.; Beemster, G.T.; Inzé, D. David and goliath: What can the tiny weed arabidopsis teach us to improve biomass production in crops? Curr. Opin. Plant Biol. 2009, 12, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Q.; Shi, J.; Scheben, A.; Zhan, J.; Wang, X.; Liu, G.; Yan, G.; King, G.J.; Edwards, D.; Wang, H. Genetic and signalling pathways of dry fruit size: Targets for genome editing-based crop improvement. Plant Biotechnol. J. 2019, 18, 1124–1140. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, Y.; McGregor, C.; Liu, S.; Luan, F.; Gao, M.; Weng, Y. Genetic architecture of fruit size and shape variation in cucurbits: A comparative perspective. Theor. Appl. Genet. 2019, 133, 1–21. [Google Scholar] [CrossRef]
- Tanksley, S.D. The genetic, developmental, and molecular bases of fruit size and shape variation in tomato. Plant Cell. 2004, 16, S181–S189. [Google Scholar] [CrossRef]
- Alpert, K.B.; Tanksley, S.D. High-resolution mapping and isolation of a yeast artificial chromosome contig containing fw2.2: A major fruit weight quantitative trait locus in tomato. Proc. Natl. Acad. Sci. USA 1996, 93, 15503–15507. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Rupe, M.A.; Dieter, J.A.; Zou, J.; Spielbauer, D.; Duncan, K.E.; Howard, R.J.; Hou, Z.; Simmons, C.R. Cell number regulator1 affects plant and organ size in maize: Implications for crop yield enhancement and heterosis. Plant Cell. 2010, 22, 1057–1073. [Google Scholar] [CrossRef] [Green Version]
- Thibivilliers, S.; Farmer, A.; Libault, M. Biological and cellular functions of the microdomain-associated FWL/CNR protein family in plants. Plants 2020, 9, 377. [Google Scholar] [CrossRef] [Green Version]
- Arthur, B.; Frédéric, G.; Nathalie, G.; Christian, C. In search of the still unknown function of FW2.2/CELL NUMBER REGU-LATOR, a major regulator of fruit size in tomato. J. Exp. Bot. 2021, 72, 5300–5311. [Google Scholar] [CrossRef]
- Li, Z.; He, C. Physalis floridana Cell Number Regulator1 encodes a cell membrane-anchored modulator of cell cycle and neg-atively controls fruit size. J. Exp. Bot. 2014, 66, 257–270. [Google Scholar] [CrossRef] [Green Version]
- De Franceschi, P.; Stegmeir, T.; Cabrera, A.; van der Knaap, E.; Rosyara, U.R.; Sebolt, A.M.; Dondini, L.; Dirlewanger, E.; Quero-Garcia, J.; Campoy, J.A.; et al. Cell number regulator genes in Prunus provide candidate genes for the control of fruit size in sweet and sour cherry. Mol. Breed. 2013, 32, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Dahan, Y.; Rosenfeld, R.; Zadiranov, V.; Irihimovitch, V. A proposed conserved role for an avocado fw2.2-like gene as a negative regulator of fruit cell division. Planta 2010, 232, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Zhang, L.; Jiang, Y.; Huang, T.; Chen, X.; Liu, Y.; Wu, J.; Yang, X.; Lin, S. EjFWLs are repressors of cell division during early fruit morphogenesis of loquat. Sci. Hortic. 2021, 287, 110261. [Google Scholar] [CrossRef]
- Kuang, C.; Li, J.; Liu, H.; Sun, X.; Zhu, X.; Hua, W. Genome-wide identification and evolutionary analysis of the fruit-weight 2.2-like gene family in polyploid oilseed rape (Brassica napus L.). DNA Cell Biol. 2020, 39, 766–782. [Google Scholar] [CrossRef]
- Xu, J.; Xiong, W.; Cao, B.; Yan, T.; Luo, T.; Fan, T.; Luo, M. Molecular characterization and functional analysis of “fruit-weight 2.2-like” gene family in rice. Planta 2013, 238, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, T.; Yuan, Z. Pomegranate PLAC8 family. Acta Hortic. 2019, 1245, 35–40. [Google Scholar] [CrossRef]
- Li, J.; Zhang, M.; Li, X.; Khan, A.; Kumar, S.; Allan, A.C.; Lin-Wang, K.; Espley, R.V.; Wang, C.; Wang, R.; et al. Pear genetics: Recent advances, new prospects, and a roadmap for the future. Hortic. Res. 2021, 9, uhab040. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Jiang, Z.Q.; Li, S.Q.; He, T.M. The development factors of Pyrus sinkiangensis Korla fragrant pear industry. Chin. Agric. Sci. Bull. 2021, 37, 159–164. (In Chinese) [Google Scholar]
- Zhang, R.-P.; Wu, J.; Li, X.-G.; Khan, M.A.; Chen, H.; Korban, S.S.; Zhang, S.-L. An AFLP, SRAP, and SSR genetic linkage map and identification of QTLs for fruit traits in Pear (Pyrus L.). Plant Mol. Biol. Rep. 2012, 31, 678–687. [Google Scholar] [CrossRef]
- Yamamoto, T.; Terakami, S.; Takada, N.; Nishio, S.; Onoue, N.; Nishitani, C.; Kunihisa, M.; Inoue, E.; Iwata, H.; Hayashi, T.; et al. Identification of QTLs controlling harvest time and fruit skin color in Japanese pear (Pyrus pyrifolia Nakai). Breed. Sci. 2014, 64, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular lo-calization. PLoS ONE 2010, 28, e11335. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Cavalcanti, A.; Chen, F.-C.; Bouman, P.; Li, W.-H. Extent of gene duplication in the genomes of drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, X.; Yue, J.-X.; Tian, D.; Chen, J.-Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol. Genet. Genom. 2008, 280, 187–198. [Google Scholar] [CrossRef]
- Wang, L.; Guo, K.; Li, Y.; Tu, Y.; Hu, H.; Wang, B.; Cui, X.; Peng, L. Expression profiling and integrative analysis of the CE-SA/CSL superfamily in rice. BMC Plant Biol. 2010, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Duan, W.; Song, X.; Tang, J.; Peng, W.; Bei, Z.; Hou, X. Retention, molecular evolution, and expression divergence of the auxin/indole acetic acid and auxin response factor gene families inbrassica rapashed light on their evolution patterns in plants. Genome Biol. Evol. 2015, 8, 302–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesbitt, T.C.; Tanksley, S.D. Comparative sequencing in the genus lycopersicon: Implications for the evolution of fruit size in the domestication of cultivated tomatoes. Genetics 2002, 162, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 337, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.Z.; Ma, X.T. Comparison of several common methods to measure the volume of fruit. China Fruits 1995, 2, 44–45. (In Chinese) [Google Scholar]
- Zhang, C.; Tanabe, K.; Wang, S.; Tamura, F.; Yoshida, A.; Matsumoto, K. The impact of cell division and cell enlargement on the evolution of fruit size in Pyrus pyrifolia. Ann. Bot. 2006, 98, 537–543. [Google Scholar] [CrossRef]
- Cong, B.; Tanksley, S.D. FW2.2 and cell cycle control in developing tomato fruit: A possible example of gene co-option in the evolution of a novel organ. Plant Mol. Biol. 2006, 62, 867–880. [Google Scholar] [CrossRef]
- Tian, J.; Zeng, B.; Luo, S.-P.; Li, X.-G.; Wu, B.; Li, J. Cloning, localization and expression analysis of two fw2.2-like genes in small- and large-fruited pear species. J. Integr. Agric. 2016, 15, 282–294. [Google Scholar] [CrossRef]
- Libault, M.; Stacey, G. Evolution of FW2.2-like (FWL) and PLAC8 genes in eukaryotes. Plant Signal. Behav. 2010, 5, 1226–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Wang, L.; Yan, L.; Li, Q.; Yong, B.; Zhu, W. Evolutionary developmental mechanisms underlying the origin and diversification of the fruits. Sci. Sin. Vitae 2019, 49, 301–319. [Google Scholar] [CrossRef]
- Azzi, L. Etude du rôle de FW2.2 dans le développement du fruit de tomate. In Pour Obtenir le Grade de Docteur; Université Sciences et Technologies-Bordeaux I: Bordeaux, France, 2013; Available online: https://theses.hal.science/tel-01089718 (accessed on 16 December 2013).
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabreira-Cagliari, C.; Dias, N.D.C.; Bohn, B.; Fagundes, D.G.D.S.; Margis-Pinheiro, M.; Bodanese-Zanettini, M.H.; Cagliari, A. Revising the PLAC8 gene family: From a central role in differentiation, proliferation, and apoptosis in mammals to a mul-tifunctional role in plants. Genome 2018, 61, 857–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.Y.; Choi, Y.I.; Shim, D.; Kim, D.Y.; Noh, E.W.; Martinoia, E.; Lee, Y. Transgenic poplar for phytoremediation. In Biotechnology and Sustainable Agriculture 2006 and Beyond; Springer: Berlin, Germany, 2007; pp. 265–271. [Google Scholar] [CrossRef]
- Arnoys, E.J.; Wang, J.L. Dual localization: Proteins in extracellular and intracellular compartments. Acta Histochem. 2007, 109, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in arabidopsis. Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef] [Green Version]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just JASMONATE-SALICYLATE antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Cong, B.; Liu, J.; Tanksley, S.D. Natural alleles at a tomato fruit size quantitative trait locus differ by heterochronic regulatory mutations. Proc. Natl. Acad. Sci. USA 2002, 99, 13606–13611. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Cong, B.; Tanksley, S.D.; Mergaert, P.; Nikovics, K.; Kelemen, Z.; Maunoury, N.; Vaubert, D.; Kondorosi, A.; Kondorosi, E. Generation and analysis of an artificial gene dosage series in tomato to study the mechanisms by which the cloned quantitative trait locus fw2.2 controls fruit size. Plant Physiol. 2003, 132, 292–299. [Google Scholar] [CrossRef] [Green Version]
- Song, W.Y.; Martinoia, E.; Lee, J.; Kim, D.; Kim, D.Y.; Vogt, E.; Shim, D.; Choi, K.S.; Hwang, I.; Lee, Y. A novel family of cys-rich membrane proteins mediates cadmium resistance in Arabidopsis. Plant Physiol. 2004, 135, 1027–1039. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Multiple sequence alignment of PbFWLs, ZmCNR1, OsFWL1, OsFWL3, OsFWL4, OsFWL5, PfCNR1, and fw2.2. Identical and similar amino acids are shown in light blue and blue, respectively. The PLAC8 structure domain is indicated by the red box. The conserved motifs CXXXXXCPC and QEYRELK are indicated by black five-pointed stars and filled circles.
Figure 1.
Multiple sequence alignment of PbFWLs, ZmCNR1, OsFWL1, OsFWL3, OsFWL4, OsFWL5, PfCNR1, and fw2.2. Identical and similar amino acids are shown in light blue and blue, respectively. The PLAC8 structure domain is indicated by the red box. The conserved motifs CXXXXXCPC and QEYRELK are indicated by black five-pointed stars and filled circles.

Figure 2.
Phylogenetic tree analysis of fw2.2/CNR proteins in pear and other plants. The solid red circles represent pears, the solid black circles represent corn, the solid blue circles represent tomatoes, the solid olive circles represent peaches, and the solid green circles represent rice.
Figure 2.
Phylogenetic tree analysis of fw2.2/CNR proteins in pear and other plants. The solid red circles represent pears, the solid black circles represent corn, the solid blue circles represent tomatoes, the solid olive circles represent peaches, and the solid green circles represent rice.

Figure 3.
Gene structures of PbFWL genes, phylogenetic relationships, and conserved motifs of PbFWL proteins. (A) Phylogenetic tree of 14 PbFWL proteins in pear. The full-length amino acid sequences of 14 PbFWL proteins were imported into MEGA 7.0 software to construct an unrooted neighbor-joining phylogenetic tree. (B) Gene structure analysis of pear fw2.2-like members. The yellow boxes represent exons. The black lines represent introns. The green boxes represent UTR. The numbers 0, 1, and 2 represent the intron phases. (C) The conserved motifs in PbFWL proteins. The six predicted conserved structural domains are indicated by differently colored boxes. The motif information is shown in Supplementary Table S3.
Figure 3.
Gene structures of PbFWL genes, phylogenetic relationships, and conserved motifs of PbFWL proteins. (A) Phylogenetic tree of 14 PbFWL proteins in pear. The full-length amino acid sequences of 14 PbFWL proteins were imported into MEGA 7.0 software to construct an unrooted neighbor-joining phylogenetic tree. (B) Gene structure analysis of pear fw2.2-like members. The yellow boxes represent exons. The black lines represent introns. The green boxes represent UTR. The numbers 0, 1, and 2 represent the intron phases. (C) The conserved motifs in PbFWL proteins. The six predicted conserved structural domains are indicated by differently colored boxes. The motif information is shown in Supplementary Table S3.

Figure 4.
Schematic diagram of chromosome distribution and interchromosomal relationships of PbFWLs in pears. (A) The distribution pattern of PbFWLs in 9 pear chromosomes, where the chromosome number is indicated at the top of each chromosome. (B) The distribution pattern synteny analysis of fw2.2-like gene family of pear; the gray lines indicate all synteny blocks in the pear genome, and the red lines indicate duplicated PbFWL gene pairs.
Figure 4.
Schematic diagram of chromosome distribution and interchromosomal relationships of PbFWLs in pears. (A) The distribution pattern of PbFWLs in 9 pear chromosomes, where the chromosome number is indicated at the top of each chromosome. (B) The distribution pattern synteny analysis of fw2.2-like gene family of pear; the gray lines indicate all synteny blocks in the pear genome, and the red lines indicate duplicated PbFWL gene pairs.

Figure 5.
Predicted cis-elements in the promoter of the PbFWL genes. The color scale at the top right indicates the number of cis-acting elements. Green color indicates the number of cis-acting elements on fw2.2-like member. There were a total of 15 cis-acting elements, including (I) MYB binding site involved in light responsiveness; (II) Gibberellin-responsive element; (III) MeJA responsiveness; (IV) light-responsive element; (V) Zein metabolism regulation; (VI) abscisic acid responsiveness; (VII) auxin-responsive element; (VIII) Meristem expression; (IX) MYB binding site involved in drought inducibility; (X) anaerobic induction; (XI) endosperm expression; (XII) salicylic acid responsiveness; (XIII) low-temperature responsiveness; (XIV) defense and stress responsiveness; and (XV) MYBHv1 binding site.
Figure 5.
Predicted cis-elements in the promoter of the PbFWL genes. The color scale at the top right indicates the number of cis-acting elements. Green color indicates the number of cis-acting elements on fw2.2-like member. There were a total of 15 cis-acting elements, including (I) MYB binding site involved in light responsiveness; (II) Gibberellin-responsive element; (III) MeJA responsiveness; (IV) light-responsive element; (V) Zein metabolism regulation; (VI) abscisic acid responsiveness; (VII) auxin-responsive element; (VIII) Meristem expression; (IX) MYB binding site involved in drought inducibility; (X) anaerobic induction; (XI) endosperm expression; (XII) salicylic acid responsiveness; (XIII) low-temperature responsiveness; (XIV) defense and stress responsiveness; and (XV) MYBHv1 binding site.

Figure 6.
Cell proliferation and cell expansion during early pear fruit morphogenesis with different fruit sizes. (A) Longitudinal sections of material tissues in different stages are shown; the bar stands for 100 µm. (B) Cell size. (C) Cell number. (D) The relative cell proliferation rate. The vertical bars represent the standard error of triplicate experiments.
Figure 6.
Cell proliferation and cell expansion during early pear fruit morphogenesis with different fruit sizes. (A) Longitudinal sections of material tissues in different stages are shown; the bar stands for 100 µm. (B) Cell size. (C) Cell number. (D) The relative cell proliferation rate. The vertical bars represent the standard error of triplicate experiments.

Figure 7.
Relative expression levels of PbFWL genes in ‘Duli’ pear, Korla fragrant pear, ‘Zaomeixiang’ pear, and ‘Yali’ pear flower bud and fruit development. The vertical bars represent the standard error of triplicate experiments.
Figure 7.
Relative expression levels of PbFWL genes in ‘Duli’ pear, Korla fragrant pear, ‘Zaomeixiang’ pear, and ‘Yali’ pear flower bud and fruit development. The vertical bars represent the standard error of triplicate experiments.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sequence characteristics of 14 genes identified in pear.
| Gene Name | Gene ID | Chr | Gene Location (bp) | AA | MW (kDa) | pI | Instability Index(Ⅱ) | GRAVY | cys | S.L. |
|---|---|---|---|---|---|---|---|---|---|---|
| PbFWL1 | rna5458 | 16 | 312,566(-)314,118 | 151 | 16.99 | 7.41 | 54.00 | −0.124 | 16 | Cell membrane |
| PbFWL2 | rna5456 | 16 | 310,397(-)314,118 | 208 | 23.08 | 5.67 | 54.46 | −0.333 | 15 | Cell membrane |
| PbFWL3 | rna12156 | 4 | 11,872,621(-)11,875,857 | 246 | 26.72 | 5.29 | 56.18 | −0.363 | 16 | Cell membrane |
| PbFWL4 | rna4422 | 12 | 12,008,588(-)12,011,010 | 251 | 27.19 | 4.93 | 56.13 | −0.445 | 15 | Cell membrane |
| PbFWL5 | rna27717 | 7 | 6,529,212(-)6,531,603 | 242 | 26.91 | 4.79 | 56.34 | −0.314 | 17 | Cell membrane |
| PbFWL6 | rna1124 | 3 | 23,944,140(-)23,945,131 | 178 | 19.54 | 7.79 | 53.62 | 0.022 | 20 | Cell membrane |
| PbFWL7 | rna4877 | 6 | 18,544,328(-)18,545,220 | 179 | 19.33 | 6.04 | 50.31 | −0.232 | 16 | Cell membrane |
| PbFWL8 | rna23574 | 7 | 17,632,755(-)17,634,097 | 242 | 27.16 | 5.81 | 45.29 | −0.135 | 19 | Cell membrane Nucleus |
| PbFWL9 | rna17541 | 15 | 14,880,874(-)14,886,180 | 415 | 47.51 | 6.58 | 49.00 | −0.484 | 18 | Cell membrane Nucleus |
| PbFWL10 | rna19452 | 12 | 1,210,378(-)1,213,360 | 180 | 19.42 | 4.87 | 60.41 | 0.239 | 17 | Cell membrane |
| PbFWL11 | rna6937 | 14 | 433,409(-)436,373 | 188 | 20.36 | 4.9 | 59.62 | 0.134 | 17 | Cell membrane |
| PbFWL12 | rna24884 | 7 | 19,812,175(-)19,824,369 | 239 | 26.55 | 5.28 | 47.81 | −0.365 | 17 | Cell membrane |
| PbFWL13 | rna27387 | 7 | 18,336,597(-)18,354,923 | 239 | 26.53 | 5.27 | 45.69 | −0.365 | 17 | Cell membrane |
| PbFWL14 | rna41755 | 1 | 5,027,259(-)5,029,618 | 239 | 26.47 | 5.28 | 41.77 | −0.318 | 17 | Cell membrane |
Table 2.
Correlation between PbFWL1/2/5 expression and pulp cell devision in 10 to 40 DAFB.
| Cell Division (C) | Expression | |||
|---|---|---|---|---|
| PbFWL1 | PbFWL2 | PbFWL3 | ||
| ‘Duli’ pear | 0.036 | 27.179 | 19.683 | 198.781 |
| Korla fragrant pear | 0.126 | 12.127 | 5.962 | 41.572 |
| ‘Zaomeixiang’ pear | 0.124 | 9.432 | 5.305 | 28.070 |
| ‘Yali’ pear | 0.132 | 4.067 | 3.410 | 15.589 |
| Correlation | r = −0.958 | r = −0.996 | r = −0.996 | |
| P = 0.042 | P = 0.004 | P = 0.004 | ||
Not: C indicates average cell cycle of pulp cells in 10 to 40 DAFB, C = log2 [NUM (40 DAFB)/NUM (10 DAFB)]; Expression (E) indicates average expression of gene in 10 to 40 DAFB, Expression = [E (10 DAFB) + E (20 DAFB) + E (30 DAFB) + E (40 DAFB)]/4.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pu, X.; Tian, J.; Li, J.; Wen, Y. Genome-Wide Identification and Expression Analysis of the fw2.2-like Gene Family in Pear. Horticulturae 2023, 9, 429. https://doi.org/10.3390/horticulturae9040429
AMA Style
Pu X, Tian J, Li J, Wen Y. Genome-Wide Identification and Expression Analysis of the fw2.2-like Gene Family in Pear. Horticulturae. 2023; 9(4):429. https://doi.org/10.3390/horticulturae9040429
Chicago/Turabian StylePu, Xiaoqiu, Jia Tian, Jiang Li, and Yue Wen. 2023. "Genome-Wide Identification and Expression Analysis of the fw2.2-like Gene Family in Pear" Horticulturae 9, no. 4: 429. https://doi.org/10.3390/horticulturae9040429
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.