
Elucidating the Anatomical Features, Adaptive and Ecological Significance of Kopsia fruticosa Roxb. (Apocynaceae)

 ,
, 

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
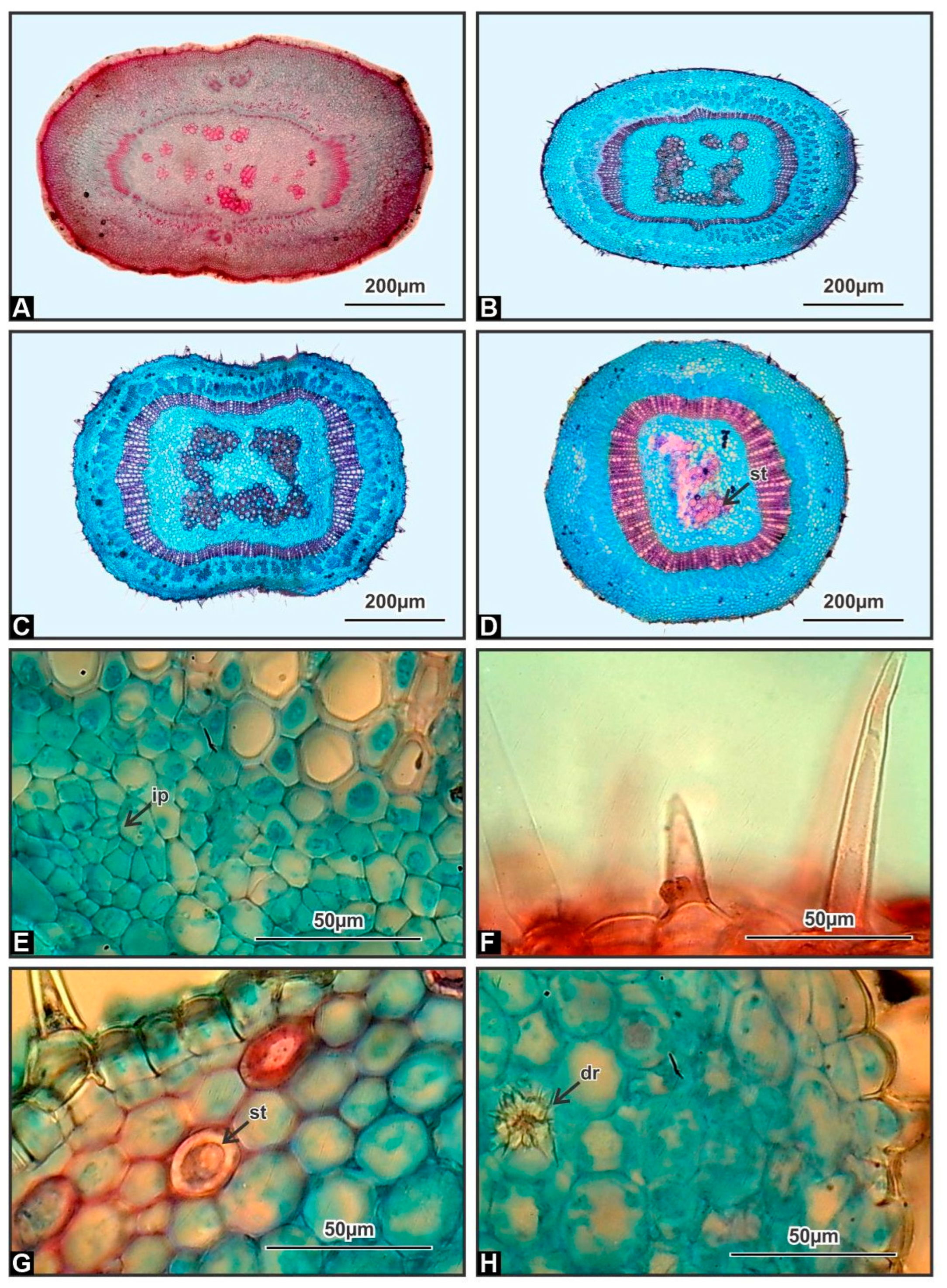
3.1. Stem
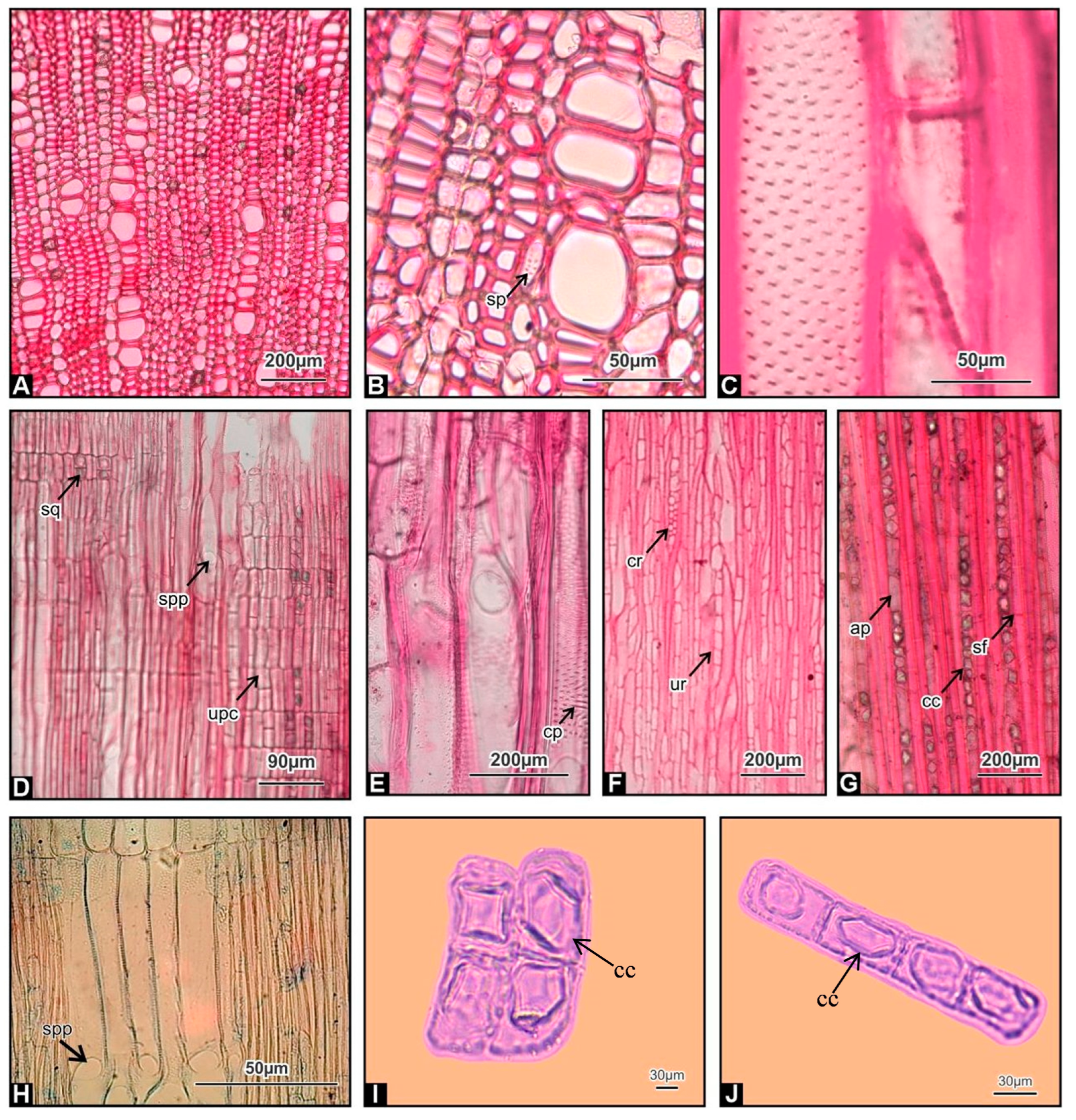
3.2. Stem Wood
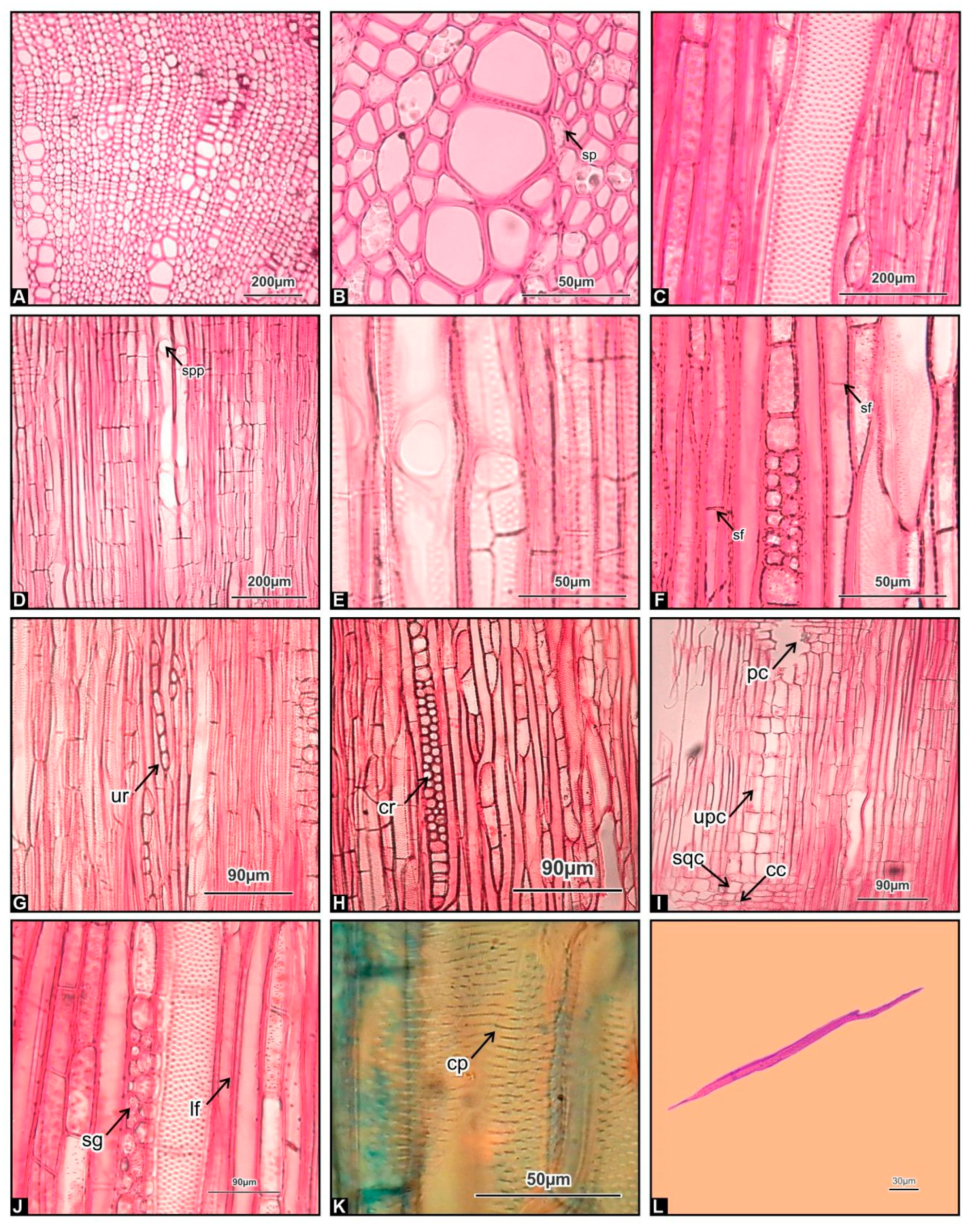
3.3. Root Wood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernández-Hernández, T.; Wiens, J.J. Why are there so many flowering plants? A multiscale analysis of plant diversification. Am. Nat. 2020, 195, 948–963. [Google Scholar] [CrossRef] [PubMed]
- Angiosperm Phylogeny Group IV. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ryan, P.R.; Delhaize, E.; Watt, M.; Richardson, A.E. Plant roots: Understanding structure and function in an ocean of complexity. Ann. Bot. 2016, 118, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Endress, M.E.; Liede-Schumann, S.; Meve, U. An updated classification for Apocynaceae. Phytotaxa 2014, 159, 175–194. [Google Scholar] [CrossRef]
- Wong, S.K.; Lim, Y.Y.; Chan, E.W. Botany, uses, phytochemistry and pharmacology of selected Apocynaceae species: A review. Phcog. Commn. 2013, 3, 2–11. [Google Scholar] [CrossRef]
- Wiart, C. Medicinal Plants of Asia and the Pacific, 1st ed.; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2006; p. 336. [Google Scholar]
- Mauger, A.; Jarret, M.; Kouklovsky, C.; Poupon, E.; Evanno, L.; Vincent, G. The chemistry of mavacurane alkaloids: A rich source of bis-indole alkaloids. Nat. Prod. Rep. 2021, 38, 1852–1886. [Google Scholar] [CrossRef]
- Kam, T.S.; Choo, Y.M. Venalstonine and dioxokopsan derivatives from Kopsiafruticosa. Phytochemistry 2004, 65, 2119–2122. [Google Scholar] [CrossRef]
- Kam, T.S.; Lim, K.H. Alkaloids of Kopsia. In The Alkaloids: Chemistry and Biology; Elsevier: Amsterdam, The Netherlands, 2008; Volume 66, pp. 1–111. [Google Scholar]
- Subramaniam, G.; Hiraku, O.; Hayashi, M.; Koyano, T.; Komiyama, K.; Kam, T.S. Biologically active aspidofractinine alkaloids from Kopsiasingapurensis. J. Nat. Prod. 2008, 71, 53–57. [Google Scholar] [CrossRef]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons: Leaves, Stem, and Wood in Relation to Taxonomy with Notes on Economic Uses; Clarendon: Oxford, UK, 1950; p. 1500. [Google Scholar]
- El-Taher, A.M.; Gendy, A.E.; Alkahtani, J.; Elshamy, A.I.; Abd-El Gawad, A.M. Taxonomic implication of integrated chemical, morphological, and anatomical attributes of leaves of eight Apocynaceae taxa. Diversity 2020, 12, 334. [Google Scholar] [CrossRef]
- Beckers, V.; Rapini, A.; Smets, E.; Lens, F. Comparative wood anatomy and origin of woodiness in subfamilies Secamonoideae and Asclepiadoideae (Apocynaceae). Taxon 2022, 71, 1230–1250. [Google Scholar] [CrossRef]
- Baas, P.; Lens, F.; Wheeler, E.A. Wood anatomy. In Flora Malesiana. Apocynaceae (Subfamilies Rauvolfioideae and Apocynoideae); Nooteboom, H.P., Ed.; Series 1, Seed plants; National Herbarium of the Netherlands: Leiden, The Netherlands, 2007; Volume 18, pp. 16–18. [Google Scholar]
- Lens, F.; Endress, M.E.; Baas, P.; Jansen, S.; Smets, E. Wood anatomy of Rauvolfioideae (Apocynaceae): A search for meaningful non-DNA characters at the tribal level. Am. J. Bot. 2008, 95, 1199–1215. [Google Scholar] [CrossRef] [Green Version]
- Johansen, D.A. Plant Microtechnique, 1st ed.; McGraw-Hill Book Company, Inc.: London, UK, 1940; p. 530. [Google Scholar]
- IAWA. List of microscopic features of hardwood identification. IAWA Bull. 1989, 10, 219–332. [Google Scholar]
- Carlquist, S. Ecological factors in wood evolution: A floristic approach. Am. J. Bot. 1977, 64, 887–896. [Google Scholar] [CrossRef]
- Kramer, J.P.; Koslowski, T. Fisiologia das Árvores; Fundação Calouste Gulbenkian: Lisboa, Portugal, 1960; p. 745. [Google Scholar]
- Patil, V.S.; Rao, K.S.; Rajput, K.S. Development of intraxylary phloem and internal cambium in Ipomoea hederifolia (Convolvulaceae). J. Torrey Bot. Soc. 2009, 136, 423–432. [Google Scholar] [CrossRef]
- Patil, V.S.; Koyani, R.D.; Sanghvi, G.V.; Rajput, K.S. Structure and development of internal phloem in Solanum pseudocapsicum (Solanaceae). IAWA J. 2014, 35, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zózimo, E.; Tamaio, N.; Vieira, R.C. Development of intraxylary phloem in the stem of Combretum rotundifolium (Combretaceae). IAWA J. 2011, 32, 14–24. [Google Scholar] [CrossRef]
- Rajput, K.S.; Kapadane, K.K.; Ramoliya, D.G.; Thacker, K.D.; Gondaliya, A.D. Inter- and Intraxylary Phloem in Vascular Plants: A Review of Subtypes, Occurrences, and Development. Forests 2022, 13, 2174. [Google Scholar] [CrossRef]
- Rao, A.R.; Malaviya, M. The distribution, structure and ontogeny of sclereids in Dendropthœ falcata (Lf) ettings. Proc. Indian Acad. Sci. 1962, 55, 239–243. [Google Scholar] [CrossRef]
- Nakata, P.A. Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar] [CrossRef]
- Sidiyasa, K.; Baas, P. Ecological and systematic wood anatomy of Alstonia (Apocynaceae). IAWA J. 1998, 19, 207–229. [Google Scholar] [CrossRef]
- Yong, S.; Furukawa, I.; Jiang, Z. Anatomical variations in the woody plants of and areas. In Desert Plants Biology and Biotechnology; Ramavat, K.G., Ed.; Springer: New York, NY, USA, 2010; pp. 136–155. [Google Scholar]
- Carlquist, S. Comparative Wood Anatomy: Systematic, Ecological, and Evolutionary Aspects of Dicotyledon Wood, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2001; p. 448. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, 2nd ed.; Clarendon Press: Oxford, UK, 1983; p. 1500. [Google Scholar]
- Jaquish, L.L.; Ewers, F.W. Seasonal conductivity and embolism in the roots and stems of two clonal ring-porous trees, Sassafras albidum (Lauraceae) and Rhus typhina (Anacardiaceae). Am. J. Bot. 2001, 88, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Longui, E.L.; Galão, A.T.D.; Rajput, K.S.; Melo, A.C.G. Anatomical investigation of root, stem and branch wood in 10-year-old Inga laurina in the context of anatomical adaptation to hydraulic and mechanical stresses. An. Biol. 2018, 40, 31–39. [Google Scholar] [CrossRef]
- Carlquist, S. Wood anatomy of Compositae: A summary, with comments on factors controlling wood evolution. Aliso 1966, 6, 25–44. [Google Scholar] [CrossRef] [Green Version]
- Mowbray, T.B.; Oosting, H.J. Vegetation gradients in relation to environment and phenology in a southern Blue Ridge gorge. Ecol. Monogr. 1968, 38, 309–344. [Google Scholar] [CrossRef]
- Tomlinson, P.B.; Craighead, F.C. Growth-ring studies on the native trees of sub-tropical Florida. In Research Trends in Plant Anatomy; Ghouse, A.K.M., Yunus, M., Eds.; Tata McGraw-Hill Publishing Company: New Delhi, India, 1972; pp. 39–51. [Google Scholar]
- Jansen, S.; Smets, E.; Baas, P. Vestures in woody plants: A review. IAWA J. 1998, 19, 347–382. [Google Scholar] [CrossRef]
- Dickison, W.C. Integrative Plant Anatomy; Harcourt Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Jansen, S.; Baas, P.; Gasson, P.; Smets, E. Vestured pits: Do they promote safer water transport? Int. J. Plant Sci. 2003, 164, 405–413. [Google Scholar] [CrossRef]
- Jansen, S.; Baas, P.; Gasson, P.; Lens, F.; Smets, E. Variation in xylem structure from tropics to tundra: Evidence from vestured pits. Proc. Natl. Acad. Sci. USA 2004, 101, 8833–8837. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Lens Objective | |
|---|---|
| (1 Ocular Unit) | (µm) |
| 4× | 25 |
| 10× | 10 |
| 40× | 2.5 |
| 100× | 1 |
| Fibre Lengths | Vessel Size | ||||
|---|---|---|---|---|---|
| Class | Subclass | Lengths (µm) | Class | Subclass | Tangential Diameter (µm) |
| Short | Extremely short | Less than 500 | Small | Extremely small | Less than 25 |
| Very short | 500–700 | Very small | 25–50 | ||
| Moderately short | 700–900 | Moderately small | 50–100 | ||
| Medium | Medium | 900–1600 | Medium | Medium | 100–200 |
| Long | Moderately long | 1600–2200 | Large | Moderately large | 200–300 |
| Very long | 2200–3000 | Very large | 300–400 | ||
| Extremely long | Over 3000 | Extremely large | More than 400 | ||
| Wood Elements | Stem Wood | Root Wood |
|---|---|---|
| Vessel length (µm) | 382 ± 3.95 | 302 ± 1.48 |
| Vessel diameter (µm) | 43 ± 0.30 | 39 ± 0.27 |
| Vessel Frequency (Number/mm2) | 47 ± 0.06 | 31 ± 0.05 |
| Fibre tracheid length (µm) | 196 ± 2.19 | 195 ± 2.15 |
| Libriform fibre length (µm) | 752 ± 6.44 | 611 ± 5.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, S.N.; Sahney, M.; Tripathi, A.; Pandey, P.; Jatav, H.S.; Minkina, T.; Rajput, V.D. Elucidating the Anatomical Features, Adaptive and Ecological Significance of Kopsia fruticosa Roxb. (Apocynaceae). Horticulturae 2023, 9, 387. https://doi.org/10.3390/horticulturae9030387
Tripathi SN, Sahney M, Tripathi A, Pandey P, Jatav HS, Minkina T, Rajput VD. Elucidating the Anatomical Features, Adaptive and Ecological Significance of Kopsia fruticosa Roxb. (Apocynaceae). Horticulturae. 2023; 9(3):387. https://doi.org/10.3390/horticulturae9030387
Chicago/Turabian StyleTripathi, Shakti Nath, Manju Sahney, Arpita Tripathi, Praveen Pandey, Hanuman Singh Jatav, Tatiana Minkina, and Vishnu D. Rajput. 2023. "Elucidating the Anatomical Features, Adaptive and Ecological Significance of Kopsia fruticosa Roxb. (Apocynaceae)" Horticulturae 9, no. 3: 387. https://doi.org/10.3390/horticulturae9030387