
Studies on the Molecular Basis of Heterosis in Arabidopsis thaliana and Vegetable Crops
 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Heterosis Research Findings in A. thaliana
2.1. Genetic Analysis
2.2. Transcriptome
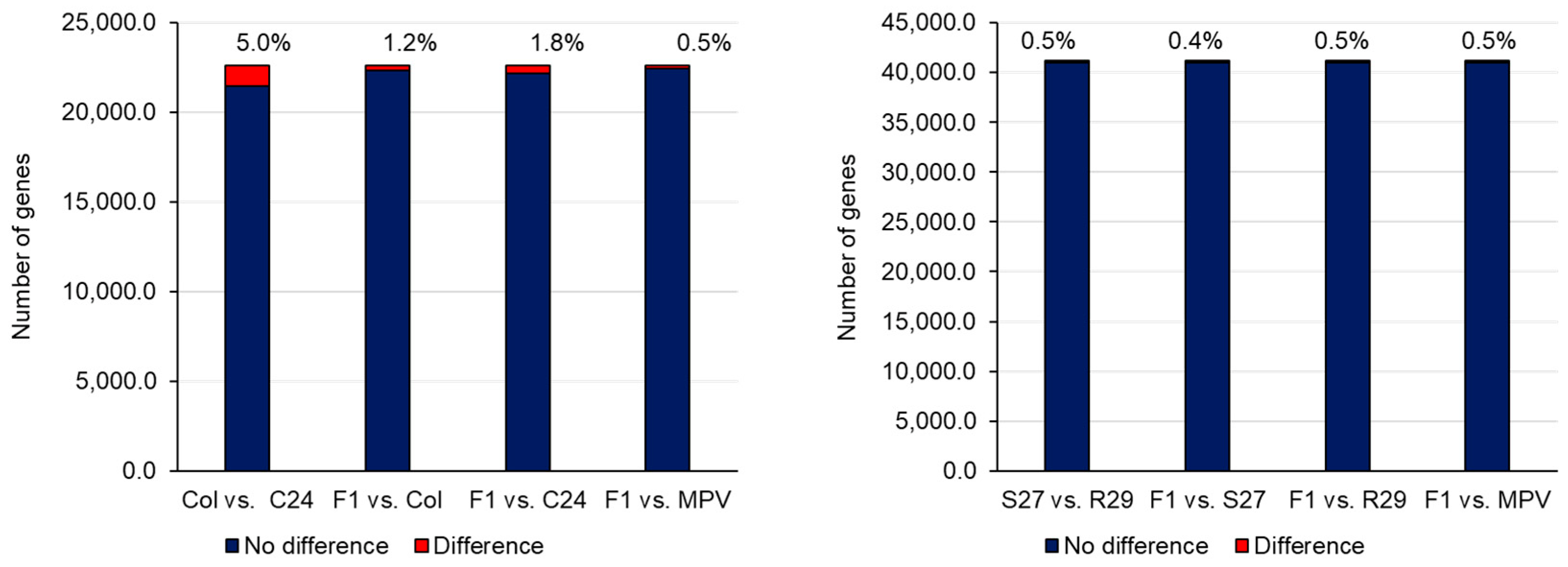
2.3. Epigenetics
2.4. Trade-Off between Growth and Disease Resistance
3. Heterosis in Leafy Vegetables
3.1. The Heterosis Phenotype in Vegetative Tissues
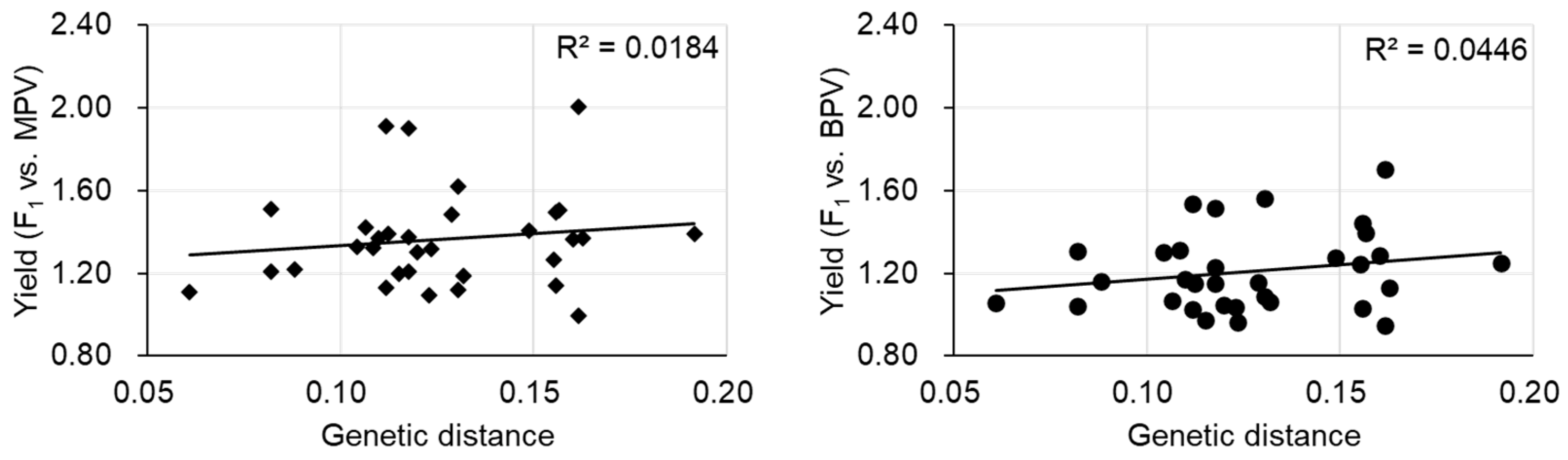
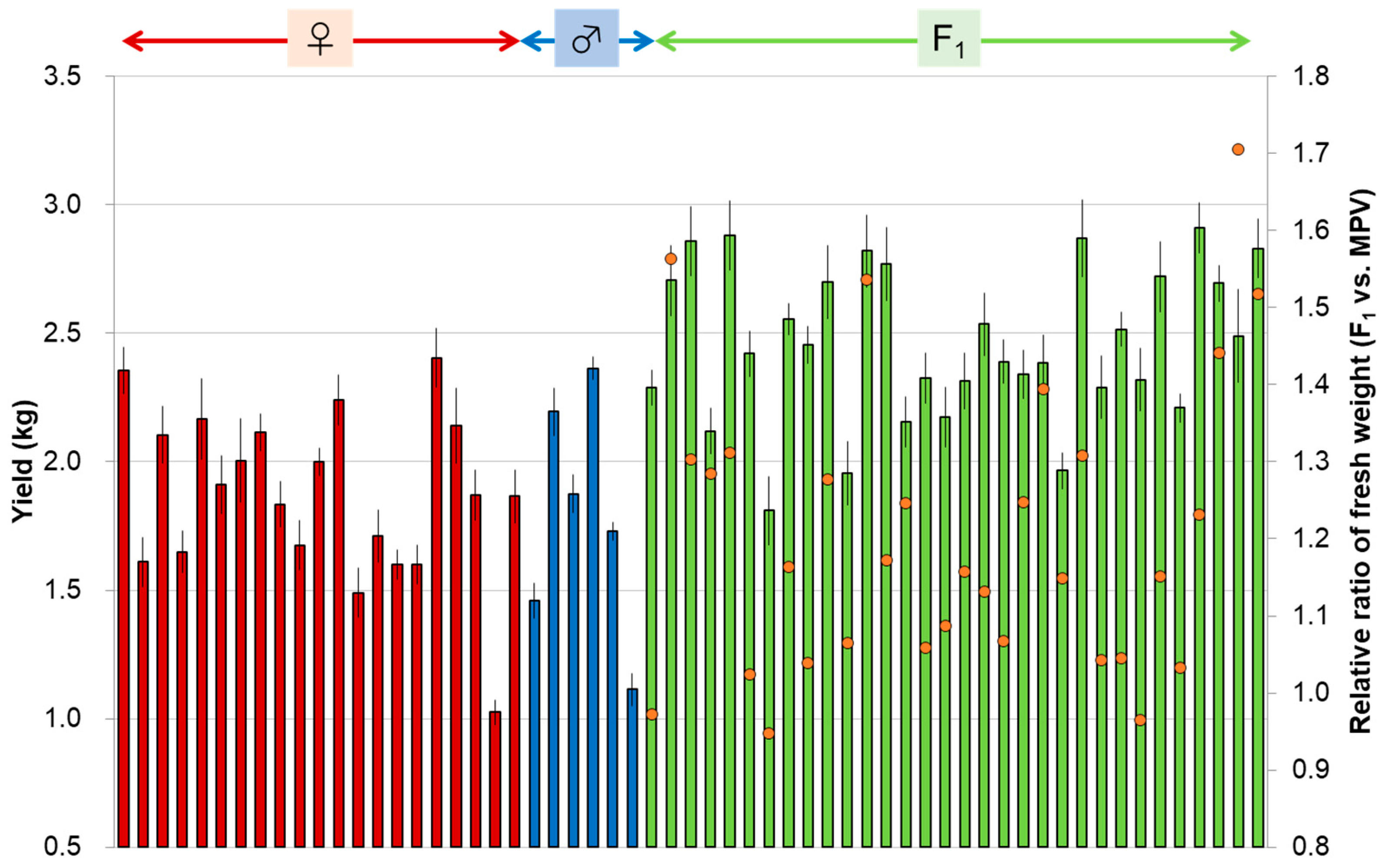
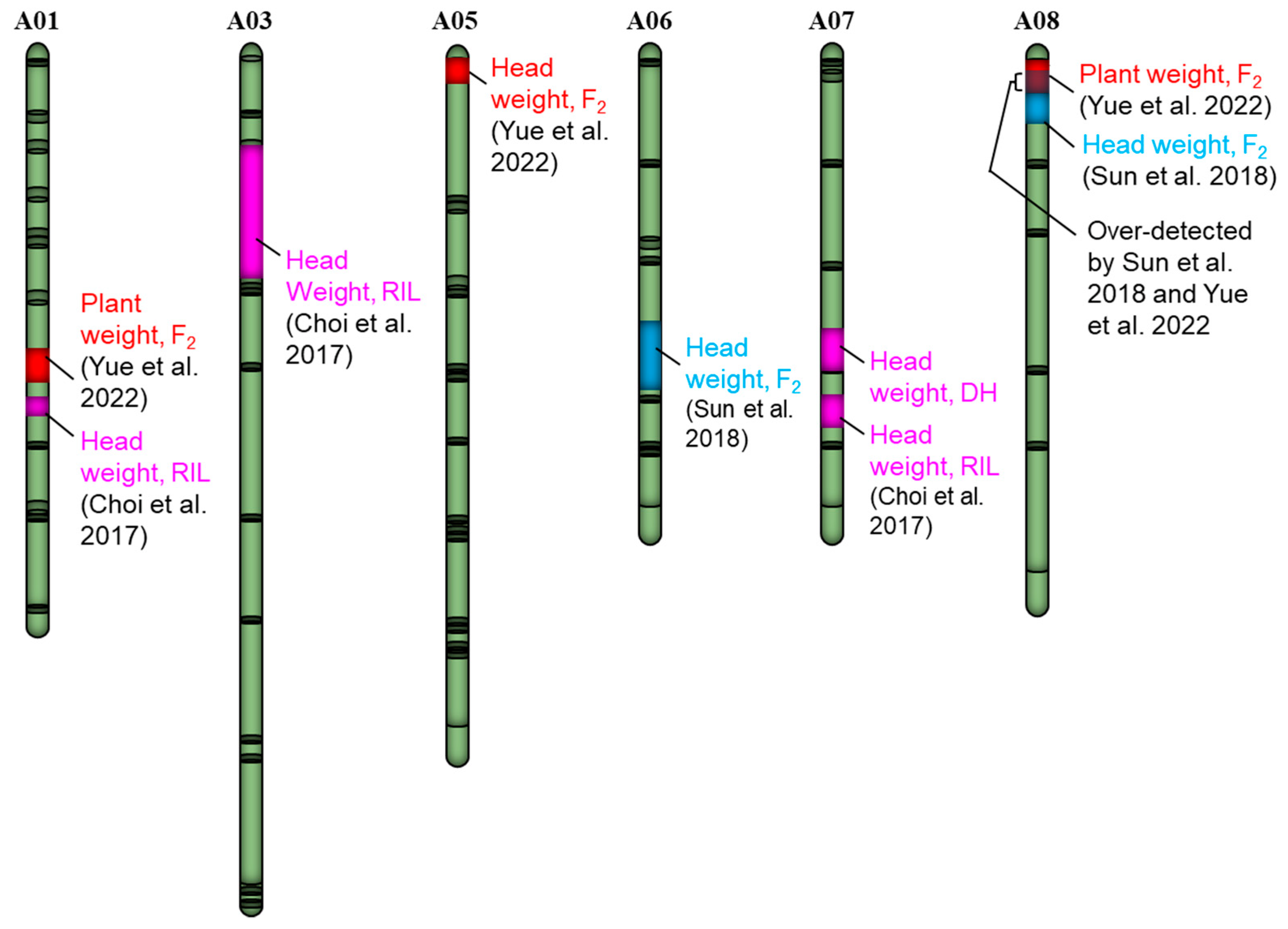
3.2. A Case Study in Brassica Vegetables
4. Heterosis in Fruit and Root Vegetables
5. Perspective
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Darwin, C. The Effects of Cross and Self Fertilisation in the Vegetable Kingdom, 1st ed.; John Murray: London, UK, 1876; pp. 1–482. [Google Scholar]
- Shull, G.H. What is “Heterosis”? Genetics 1948, 33, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Crow, J.F. 90 years ago: The beginning of hybrid maize. Genetics 1998, 148, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.H.; Yuan, L.P. Hybrid rice achievements, development and prospect in China. J. Integr. Agric. 2015, 14, 197–205. [Google Scholar] [CrossRef]
- Wakchaure, R.; Ganguly, S.; Praveen, P.K.; Sharma, S.; Kumar, A.; Mahajan, T.; Qadri, K. Importance of heterosis in animals: A review. Int. J. Adv. Eng. Technol. Innov. Sci. 2015, 1, 1–5. [Google Scholar]
- Bruce, A.B. The mendelian theory of heredity and the augmentation of vigor. Science 1910, 32, 627–628. [Google Scholar] [CrossRef]
- Jones, D.F. Dominance of linked factors as a means of accounting for heterosis. Proc. Natl. Acad. Sci. USA 1917, 3, 310–312. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, S.; Wake, T.; Nakamura, H.; Minamiyama, M.; Araki-Nakamura, S.; Ohmae-Shinohara, K.; Koketsu, E.; Okamura, S.; Miura, K.; Kawaguchi, H.; et al. The dominance model for heterosis explains culm length genetics in a hybrid sorghum variety. Sci. Rep. 2021, 11, 4532. [Google Scholar] [CrossRef]
- Busch, R.H.; Luchen, K.A.; Frohberg, R.C. F1 hybrids versus random F5 line performance and estimates of genetic effects in spring wheat. Crop Sci. 1971, 11, 357–361. [Google Scholar] [CrossRef]
- Wang, L.; Greaves, I.K.; Groszmann, M.; Wu, L.M.; Dennis, E.S.; Peacock, W.J. Hybrid mimics and hybrid vigor in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, E4959–E4967. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Zhang, Y.; Liao, Y.; Dennis, E.S.; Peacock, W.J.; Wu, X. Rice hybrid mimics have stable yields equivalent to those of the F1 hybrid and suggest a basis for hybrid vigour. Planta 2021, 254, 51. [Google Scholar] [CrossRef]
- Zhang, Y.; Ovenden, B.; He, Y.; Ye, W.; Wu, X.; Peacock, W.J.; Dennis, E.S. Hybrid vigour and hybrid mimics in Japonica rice. Agronomy 2022, 12, 1559. [Google Scholar] [CrossRef]
- Hull, F.H. Recurrent selection and overdominance. In Heterosis; Gowen, J.W., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 451–473. [Google Scholar]
- Krieger, U.; Lippman, Z.B.; Zamir, D. The flowering gene SINGLE FLOWER TRUSS drives heterosis for yield in tomato. Nat. Genet. 2010, 42, 459–463. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef]
- Yu, D.; Gu, X.; Zhang, S.; Dong, S.; Miao, H.; Gebretsadik, K.; Bo, K. Molecular basis of heterosis and related breeding strategies reveal its importance in vegetable breeding. Hortic. Res. 2021, 8, 120. [Google Scholar] [CrossRef]
- Guo, T.; Yang, N.; Tong, H.; Pan, Q.; Yang, X.; Tang, J.; Wang, J.; Li, J.; Yan, J. Genetic basis of grain yield heterosis in an “immortalized F2” maize population. Theor. Appl. Genet. 2014, 127, 2149–2158. [Google Scholar] [CrossRef]
- Huang, X.; Yang, S.; Gong, J.; Zhao, Q.; Feng, Q.; Zhan, Q.; Zhao, Y.; Li, W.; Cheng, B.; Xia, J.; et al. Genomic architecture of heterosis for yield traits in rice. Nature 2016, 537, 629–633. [Google Scholar] [CrossRef]
- Fujimoto, R.; Uezono, K.; Ishikura, S.; Osabe, K.; Peacock, W.J.; Dennis, E.S. Recent research on the mechanism of heterosis is important for crop and vegetable breeding systems. Breed. Sci. 2018, 68, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Fortuny, A.P.; Bueno, R.A.; Pereira da Costa, J.H.; Zanor, M.I.; Rodríguez, G.R. Tomato fruit quality traits and metabolite content are affected by reciprocal crosses and heterosis. J. Exp. Bot. 2021, 72, 5407–5425. [Google Scholar] [CrossRef]
- Kaushik, P.; Plazas, M.; Prohens, J.; Vilanova, S.; Gramazio, P. Diallel genetic analysis for multiple traits in eggplant and assessment of genetic distances for predicting hybrids performance. PLoS ONE 2018, 13, e0199943. [Google Scholar] [CrossRef] [Green Version]
- Yue, L.; Zhang, S.; Zhang, L.; Liu, Y.; Cheng, F.; Li, G.; Zhang, S.; Zhang, H.; Sun, R.; Li, F. Heterotic prediction of hybrid performance based on genome-wide SNP markers and the phenotype of parental inbred lines in heading Chinese cabbage (Brassica rapa L. ssp. pekinensis). Sci. Hortic. 2022, 296, 110907. [Google Scholar] [CrossRef]
- Kawamura, K.; Kawanabe, T.; Shimizu, M.; Nagano, A.J.; Saeki, N.; Okazaki, K.; Kaji, M.; Dennis, E.S.; Osabe, K.; Fujimoto, R. Genetic distance of inbred lines of Chinese cabbage and its relationship to heterosis. Plant Gene 2016, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.J. Genomic and epigenetic insights into the molecular bases of heterosis. Nat. Rev. Genet. 2013, 14, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Lippman, Z.B.; Zamir, D. Heterosis: Revisiting the magic. Trends Genet. 2007, 23, 60–66. [Google Scholar] [CrossRef]
- Groszmann, M.; Greaves, I.K.; Fujimoto, R.; Peacock, W.J.; Dennis, E.S. The role of epigenetics in hybrid vigour. Trends Genet. 2013, 29, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, P.; Signore, A. How has the consistency of the common catalogue of varieties of vegetable species changed in the last ten years? Sci. Hortic. 2021, 277, 109805. [Google Scholar] [CrossRef]
- Yamagishi, H.; Bhat, S.R. Cytoplasmic male sterility in Brassicaceae crops. Breed. Sci. 2014, 64, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, R.; Nishio, T. Self-Incompatibility. Adv. Bot. Res. 2007, 45, 139–154. [Google Scholar]
- Chen, L.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef]
- Singh, S.; Dey, S.S.; Bhatia, R.; Kumar, R.; Behera, T.K. Current understanding of male sterility systems in vegetable Brassicas and their exploitation in hybrid breeding. Plant Reprod. 2019, 32, 231–256. [Google Scholar] [CrossRef]
- Seymour, D.K.; Chae, E.; Grimm, D.G.; Pizarro, C.M.; Habring-Müller, A.; Vasseur, F.; Rakitsch, B.; Borgwardt, K.M.; Koenig, D.; Weigel, D. Genetic architecture of nonadditive inheritance in Arabidopsis thaliana hybrids. Proc. Natl. Acad. Sci. USA 2016, 113, E7317–E7326. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Wang, X.; Ren, D.; Huang, H.; Xu, M.; He, G.; Deng, X.W. Genomic architecture of biomass heterosis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 8101–8106. [Google Scholar] [CrossRef] [Green Version]
- Kusterer, B.; Piepho, H.P.; Utz, H.F.; Schön, C.C.; Muminovic, J.; Meyer, R.C.; Altmann, T.; Melchinger, A.E. Heterosis for biomass-related traits in Arabidopsis investigated by quantitative trait loci analysis of the triple testcross design with recombinant inbred lines. Genetics 2007, 177, 1839–1850. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.C.; Kusterer, B.; Lisec, J.; Steinfath, M.; Becher, M.; Scharr, H.; Melchinger, A.E.; Selbig, J.; Schurr, U.; Willmitzer, L.; et al. QTL analysis of early stage heterosis for biomass in Arabidopsis. Theor. Appl. Genet. 2010, 120, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Lisec, J.; Steinfath, M.; Meyer, R.C.; Selbig, J.; Melchinger, A.E.; Willmitzer, L.; Altmann, T. Identification of heterotic metabolite QTL in Arabidopsis thaliana RIL and IL populations. Plant J. 2009, 59, 777–788. [Google Scholar] [CrossRef]
- Andorf, S.; Meyer, R.C.; Selbig, J.; Altmann, T.; Repsilber, D. Integration of a systems biological network analysis and QTL results for biomass heterosis in Arabidopsis thaliana. PLoS ONE 2012, 7, e49951. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, R.; Taylor, J.M.; Shirasawa, S.; Peacock, W.J.; Dennis, E.D. Heterosis of Arabidopsis hybrids between C24 and Col is associated with increased photosynthesis capacity. Proc. Natl. Acad. Sci. USA 2012, 109, 7109–7114. [Google Scholar] [CrossRef] [Green Version]
- Saeki, N.; Kawanabe, T.; Ying, H.; Shimizu, M.; Kojima, M.; Abe, H.; Okazaki, K.; Kaji, M.; Taylor, J.M.; Sakakibara, H.; et al. Molecular and cellular characteristics of hybrid vigour in a commercial hybrid of Chinese cabbage. BMC Plant Biol. 2016, 16, 45. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Kim, E.D.; Ha, M.; Lackey, E.; Liu, J.; Zhang, Y.; Sun, Q.; Chen, Z.J. Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nature 2009, 457, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; He, H.; Li, J.; Chen, W.; Wang, X.; Guo, L.; Peng, Z.; He, G.; Zhong, S.; Qi, Y.; et al. Genome-wide analysis of DNA methylation and gene expression changes in two Arabidopsis ecotypes and their reciprocal hybrids. Plant Cell 2012, 24, 875–892. [Google Scholar] [CrossRef] [Green Version]
- Groszmann, M.; Gonzalez-Bayon, R.; Lyons, R.L.; Greaves, I.K.; Kazan, K.; Peacock, W.J.; Dennis, E.S. Hormone-regulated defense and stress response networks contribute to heterosis in Arabidopsis F1 hybrids. Proc. Natl. Acad. Sci. USA 2015, 112, E6397–E6406. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Peral, M.M.; Trigueros, M.; Sherman, B.; Ying, H.; Taylor, J.M.; Peacock, W.J.; Dennis, E.S. Patterns of gene expression in developing embryos of Arabidopsis hybrids. Plant J. 2017, 89, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, L.M.; Greaves, I.K.; Zhu, A.; Dennis, E.S.; Peacock, W.J. PIF4-controlled auxin pathway contributes to hybrid vigor in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, E3555–E3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Bayon, R.; Shen, Y.; Groszmann, M.; Zhu, A.; Wang, A.; Allu, A.D.; Dennis, E.S.; Peacock, W.J.; Greaves, I.K. Senescence and defense pathways contribute to heterosis. Plant Physiol. 2019, 180, 240–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.; Greaves, I.K.; Liu, P.C.; Wu, L.; Dennis, E.S.; Peacock, W.J. Early changes of gene activity in developing seedlings of Arabidopsis hybrids relative to parents may contribute to hybrid vigour. Plant J. 2016, 88, 597–607. [Google Scholar] [CrossRef]
- Meyer, R.C.; Witucka-Wall, H.; Becher, M.; Blacha, A.; Boudichevskaia, A.; Dörmann, P.; Fiehn, O.; Friedel, S.; von Korff, M.; Lisec, J.; et al. Heterosis manifestation during early Arabidopsis seedling development is characterized by intermediate gene expression and enhanced metabolic activity in the hybrids. Plant J. 2012, 71, 669–683. [Google Scholar] [CrossRef]
- Liu, W.; He, G.; Deng, X.W. Biological pathway expression complementation contributes to biomass heterosis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2023278118. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirasch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Yue, H.; Nie, X.; Yan, Z.; Weining, S. N6-methyladenosine regulatory machinery in plants: Composition, function and evolution. Plant Biotechnol. J. 2019, 17, 1194–1208. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, X.; Bao, M.; Song, X.; Zhang, Y.; Wang, H.; Xie, H.; Mao, F.; Wang, S.; Jin, H.; et al. Transcriptome-wide analysis of RNA m6A methylation and gene expression changes among two Arabidopsis ecotypes and their reciprocal hybrids. Front. Plant Sci. 2021, 12, 685189. [Google Scholar] [CrossRef]
- Fujimoto, R.; Sasaki, T.; Ishikawa, R.; Osabe, K.; Kawanabe, T.; Dennis, E.S. Molecular mechanisms of epigenetic variation in plants. Int. J. Mol. Sci. 2012, 13, 9900–9922. [Google Scholar] [CrossRef] [Green Version]
- Kawakatsu, T.; Ecker, J.R. Diversity and dynamics of DNA methylation: Epigenomic resources and tools for crop breeding. Breed. Sci. 2019, 69, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Greaves, I.K.; Groszmann, M.; Ying, H.; Taylor, J.M.; Peacock, W.J.; Dennis, E.S. Trans chromosomal methylation in Arabidopsis hybrids. Proc. Natl. Acad. Sci. USA 2012, 109, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Greaves, I.K.; Dennis, E.S.; Peacock, W.J. Genome-wide analyses of four major histone modifications in Arabidopsis hybrids at the germinating seed stage. BMC Genom. 2017, 18, 137. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, D.; Lang, Z.; He, L.; Yang, L.; Zeng, L.; Li, Y.; Zhao, C.; Huang, H.; Zhang, H.; et al. Methylation interactions in Arabidopsis hybrids require RNA-directed DNA methylation and are influenced by genetic variation. Proc. Natl. Acad. Sci. USA 2016, 113, E4248–E4256. [Google Scholar] [CrossRef] [Green Version]
- Rigal, M.; Becker, C.; Pélissier, T.; Pogorelcnik, R.; Devos, J.; Ikeda, Y.; Weigel, D.; Mathieu, O. Epigenome confrontation triggers immediate reprogramming of DNA methylation and transposon silencing in Arabidopsis thaliana F1 epihybrids. Proc. Natl. Acad. Sci. USA 2016, 113, E2083–E2092. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, T.; Ishikura, S.; Miyaji, N.; Sasaki, T.; Wu, L.M.; Itabashi, E.; Takada, S.; Shimizu, M.; Takasaki-Yasuda, T.; Osabe, K.; et al. Role of DNA methylation in hybrid vigor in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, E6704–E6711. [Google Scholar] [CrossRef] [Green Version]
- Dapp, M.; Reinders, J.; Bédiée, A.; Balsera, C.; Bucher, E.; Theiler, G.; Granier, C.; Paszkowski, J. Heterosis and inbreeding depression of epigenetic Arabidopsis hybrids. Nat. Plants 2015, 1, 15092. [Google Scholar] [CrossRef]
- Lauss, K.; Wardenaar, R.; Oka, R.; van Hulten, M.H.A.; Guryev, V.; Keurentjes, J.J.B.; Stam, M.; Johannes, F. Parental DNA methylation states are associated with heterosis in epigenetic hybrids. Plant Physiol. 2018, 176, 1627–1645. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, Y.; Xu, T.; Srivastava, A.K.; Wang, D.; Zeng, L.; Yang, L.; He, L.; Zhang, H.; Zheng, Z.; et al. The chromatin remodeler DDM1 promotes hybrid vigor by regulating salicylic acid metabolism. Cell Discov. 2016, 2, 16027. [Google Scholar] [CrossRef] [Green Version]
- Miyaji, N.; Fujimoto, R. Hybrid vigor: Importance of epigenetic processes and consequences for breeding. Adv. Bot. Res. 2018, 88, 247–275. [Google Scholar]
- Miller, M.; Song, Q.; Shi, X.; Juenger, E.T.; Chen, Z.J. Natural variation in timing of stress-responsive gene expression predicts heterosis in intraspecific hybrids of Arabidopsis. Nat. Commun. 2015, 6, 7453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Li, B.; Zheng, X.Y.; Li, J.; Yang, M.; Dong, X.; He, G.; An, C.; Deng, X.W. Salicylic acid biosynthesis is enhanced and contributes to increased biotrophic pathogen resistance in Arabidopsis hybrids. Nat. Commun. 2015, 6, 7309. [Google Scholar]
- Calvo-Baltanás, V.; Wang, J.; Chae, E. Hybrid incompatibility of the plant immune system: An opposite force to heterosis equilibrating hybrid performances. Front. Plant Sci. 2021, 11, 576796. [Google Scholar] [CrossRef]
- Yang, L.; Liu, P.; Wang, X.; Jia, A.; Ren, D.; Tang, Y.; Tang, Y.; Deng, X.W.; He, G. A central circadian oscillator confers defense heterosis in hybrids without growth vigor costs. Nat. Commun. 2021, 12, 2317. [Google Scholar] [CrossRef]
- Shull, G.H. The composition of a field of maize. J. Hered. 1908, 4, 296–301. [Google Scholar] [CrossRef] [Green Version]
- East, E.M.; Jones, D.F. Inbreeding and Outbreeding: Their Genetic and Sociological Significance; Lippincott: Philadelphia, PA, USA, 1919; p. 285. [Google Scholar]
- Hochholdinger, F.; Hoecker, N. Towards the molecular basis of heterosis. Trends Plant Sci. 2007, 12, 427–432. [Google Scholar] [CrossRef]
- Barth, S.; Busimi, A.K.; Friedrich Utz, H.; Melchinger, A.E. Heterosis for biomass yield and related traits in five hybrids of Arabidopsis thaliana L. Heynh. Heredity 2003, 91, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.C.; Törjék, O.; Becher, M.; Altmann, T. Heterosis of biomass production in Arabidopsis. Establishment during early development. Plant Physiol. 2004, 134, 1813–1823. [Google Scholar] [CrossRef] [Green Version]
- Syed, N.H.; Chen, Z.J. Molecular marker genotypes, heterozygosity and genetic interactions explain heterosis in Arabidopsis thaliana. Heredity 2005, 94, 295–304. [Google Scholar] [CrossRef]
- Groszmann, M.; Gonzalez-Bayon, R.; Greaves, I.K.; Wang, L.; Huen, A.K.; Peacock, W.J.; Dennis, E.S. Intraspecific Arabidopsis hybrids show different patterns of heterosis despite the close relatedness of the parental genomes. Plant Physiol. 2014, 166, 265–280. [Google Scholar] [CrossRef] [Green Version]
- van Hulten, M.H.A.; Paulo, M.J.; Kruijer, W.; Vries, H.B.D.; Kemperman, B.; Becker, F.F.M.; Yang, J.; Lauss, K.; Stam, M.E.; van Eeuwijk, F.A.; et al. Assessment of heterosis in two Arabidopsis thaliana common-reference mapping populations. PLoS ONE 2018, 13, e0205564. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wu, L.M.; Greaves, I.K.; Dennis, E.S.; Peacock, W.J. In Arabidopsis hybrids and Hybrid Mimics, up-regulation of cell wall biogenesis is associated with the increased plant size. Plant Direct 2019, 3, e00174. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Su, T.; Zhang, D.; Wang, W.; Xin, X.; Yu, Y.; Zhao, X.; Yu, S.; Zhang, F. Genome-wide analysis of changes in miRNA and target gene expression reveals key roles in heterosis for Chinese cabbage biomass. Hortic. Res. 2021, 8, 39. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Ahmed, N.U.; Jung, H.J.; Kim, H.T.; Park, J.I.; Nou, I.S. Discovery of candidate genes for heterosis breeding in Brassica oleracea L. Acta Physiol. Plant. 2017, 39, 180. [Google Scholar] [CrossRef]
- Verma, V.K.; Kalia, P. Combining ability analysis and its relationship with gene action and heterosis in early maturity cauliflower. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 877–884. [Google Scholar] [CrossRef]
- Basunanda, P.; Radoev, M.; Ecke, W.; Friedt, W.; Becker, H.C.; Snowdon, R.J. Comparative mapping of quantitative trait loci involved in heterosis for seedling and yield traits in oilseed rape (Brassica napus L.). Theor. Appl. Genet. 2010, 120, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Wolko, J.; Dobrzycka, A.; Bocianowski, J.; Bartkowiak-Broda, I. Estimation of heterosis for yield-related traits for single cross and three-way cross hybrids of oilseed rape (Brassica napus L.). Euphytica 2019, 215, 156. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Wang, A.; Zhang, Y.; Dennis, E.S.; Peacock, W.J.; Greaves, A. Early establishment of photosynthesis and auxin biosynthesis plays a key role in early biomass heterosis in Brassica napus (Canola) hybrids. Plant Cell Physiol. 2020, 61, 1134–1143. [Google Scholar] [CrossRef]
- Aakanksha; Yadava, S.K.; Yadav, B.G.; Gupta, V.; Mukhopadhyay, A.; Pental, D.; Pradhan, A.K. Genetic analysis of heterosis for yield influencing traits in Brassica juncea using a doubled haploid population and its backcross progenies. Front. Plant Sci. 2021, 12, 721631. [Google Scholar] [CrossRef]
- Wang, Q.; Yan, T.; Long, Z.; Huang, L.Y.; Zhu, Y.; Xu, Y.; Chen, X.; Pak, H.; Li, J.; Wu, D.; et al. Prediction of heterosis in the recent rapeseed (Brassica napus) polyploid by pairing parental nucleotide sequences. PLoS Genet. 2021, 17, e1009879. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xiong, J.; Shalby, N.; Zhuo, C.; Jia, Y.; Yang, Q.Y.; Tu, J. Comparison of dynamic 3D chromatin architecture uncovers heterosis for leaf size in Brassica napus. J. Adv. Res. 2022, 42, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Tamta, S.; Singh, J. Heterosis in tomato for growth and yield traits. Int. J. Veg. Sci. 2018, 24, 169–179. [Google Scholar] [CrossRef]
- Chandel, R.; Sadashiva, A.T.; Ravishankar, K.V.; Das, A.; Rout, B.M.; Singh, S. Genetic combining, heterosis analysis for horticultural traits in tomato (Solanum lycopersicum L.) using ToLCV-resistant lines and molecular validation of Ty genes. Plant Genet. Resour. 2021, 19, 512–521. [Google Scholar] [CrossRef]
- Rajendran, S.; Bae, J.H.; Park, M.W.; Oh, J.H.; Jeong, H.W.; Lee, Y.K.; Park, S.J. Tomato yield effects of reciprocal hybridization of Solanum lycopersicum cultivars M82 and Micro-Tom. Plant Breed. Biotechnol. 2022, 10, 37–48. [Google Scholar] [CrossRef]
- Kakizaki, Y. Hybrid vigor in egg-plants and its practical utilization. Genetics 1931, 16, 1–25. [Google Scholar] [CrossRef]
- Kaushik, P.; Prohens, J.; Vilanova, S.; Gramazio, P.; Plazas, M. Phenotyping of eggplant wild relatives and interspecific hybrids with conventional and phenomics descriptors provides insight for their potential utilization in breeding. Front. Plant Sci. 2016, 7, 677. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, P. Line × Tester analysis for morphological and fruit biochemical traits in eggplant (Solanum melongena L.) using wild relatives as testers. Agronomy 2019, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Sharma, V.; Jain, B.T.; Kaushik, P. Heterosis breeding in eggplant (Solanum melongena L.): Gains and provocations. Plants 2020, 9, 403. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Bradbury, P.J.; Brown, P.J.; Hung, H.; Sun, Q.; Flint-Garcia, S.; Rocheford, T.R.; McMullen, M.D.; Holland, J.B.; Buckler, E.S. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat. Genet. 2011, 43, 159–162. [Google Scholar] [CrossRef]
- Ko, D.K.; Rohozinski, D.; Song, Q.; Taylor, S.H.; Juenger, T.E.; Harmon, F.G.; Chen, Z.J. Temporal shift of circadian-mediated gene expression and carbon fixation contributes to biomass heterosis in maize hybrids. PLoS Genet. 2016, 12, e1006197. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Zhang, Z.; Tan, X.; Jiang, Y.; Gao, J.; Lin, L.; Wang, Z.; Ren, J.; Wang, X.; Qin, L.; et al. Association of the molecular regulation of ear leaf senescence/stress response and photosynthesis/metabolism with heterosis at the reproductive stage in maize. Sci. Rep. 2016, 6, 29843. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Mezmouk, S.; Baumgarten, A.; Buckler, E.S.; Guill, K.E.; McMullen, M.D.; Mumm, R.H.; Ross-Ibarra, J. Incomplete dominance of deleterious alleles contributes substantially to trait variation and heterosis in maize. PLoS Genet. 2017, 13, e1007019. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Q.; Chen, M.; Ding, Y.; Yang, X.; Liu, J.; Li, X.; Zhou, C.; Tian, Q.; Lu, Y.; et al. Genome-wide identification and analysis of heterotic loci in three maize hybrids. Plant Biotechnol. J. 2020, 18, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Birdseye, D.; de Boer, L.A.; Bai, H.; Zhou, P.; Shen, Z.; Schmelz, E.A.; Springer, N.M.; Briggs, S.P. Plant height heterosis is quantitatively associated with expression levels of plastid ribosomal proteins. Proc. Natl. Acad. Sci. USA 2021, 118, e2109332118. [Google Scholar] [CrossRef]
- Song, G.S.; Zhai, H.L.; Peng, Y.G.; Zhang, L.; Wei, G.; Chen, X.Y.; Xiao, Y.G.; Wang, L.; Chen, Y.J.; Wu, B.; et al. Comparative transcriptional profiling and preliminary study on heterosis mechanism of super-hybrid rice. Mol. Plant 2010, 3, 1012–1025. [Google Scholar] [CrossRef]
- Dan, Z.; Liu, P.; Huang, W.; Zhou, W.; Yao, G.; Hu, J.; Zhu, R.; Lu, B.; Zhu, Y. Balance between a higher degree of heterosis and increased reproductive isolation: A strategic design for breeding inter-subspecific hybrid rice. PLoS ONE 2014, 9, e93122. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.; Hu, W.; Zhang, B.; Xing, Y. The regulatory network mediated by circadian clock genes is related to heterosis in rice. J. Integr. Plant Biol. 2015, 57, 300–312. [Google Scholar] [CrossRef]
- Zhu, D.; Zhou, G.; Xu, C.; Zhang, Q. Genetic components of heterosis for seedling traits in an elite rice hybrid analyzed using an immortalized F2 population. J. Genet. Genom. 2016, 43, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Ni, Z.; Yao, Y.; Zhang, Y.; Sun, Q. Identification of differentially expressed proteins between hybrid and parents in wheat (Triticum aestivum L.) seedling leaves. Theor. Appl. Genet. 2009, 118, 213–225. [Google Scholar] [CrossRef]
- Yi, H.; Lee, J.; Song, H.; Dong, X.; Hur, Y. Genome-wide analysis of heterosis-related genes in non-heading Chinese cabbage. J. Plant Biotechnol. 2017, 44, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Duan, W.; Chen, Z.; Yuan, J.; Xiao, D.; Hou, X.; Li, Y. Enhanced photosynthetic activity in pak choi hybrids is associated with increased grana thylakoids in chloroplasts. Plant J. 2020, 103, 2211–2224. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jayasinghege, C.P.A.; Guo, J.; Zhang, E.; Wang, X.; Xu, Z. Comparative transcriptomic analysis of gene expression inheritance patterns associated with cabbage head heterosis. Plants 2021, 10, 275. [Google Scholar]
- Li, X.; Lv, H.; Zhang, B.; Fang, Z.; Yang, L.; Zhuang, M.; Liu, Y.; Li, Z.; Wang, Y.; Zhang, Y. Dissection of two QTL clusters underlying yield-related heterosis in the cabbage founder parent 01-20. Hortic. Plant J. 2023, 9, 77–88. [Google Scholar] [CrossRef]
- Li, H.; Yuan, J.; Wu, M.; Han, Z.; Li, L.; Jiang, H.; Jia, Y.; Han, X.; Liu, M.; Sun, D.; et al. Transcriptome and DNA methylome reveal insights into yield heterosis in the curds of broccoli (Brassica oleracea L. var. italic). BMC Plant Biol. 2018, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Chen, L.; Wei, T.; Zhou, H.; Bai, C.; Yan, X.; Miao, Z.; Xie, J.; Zhang, L. Transcriptome analysis of biological pathways associated with heterosis in Chinese cabbage. Genomics 2020, 112, 4732–4741. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, S.; Hua, S.; Shen, E.; Ye, C.Y.; Cai, D.; Timko, M.P.; Zhu, Q.H.; Fan, L. Analysis of transcriptional and epigenetic changes in hybrid vigor of allopolyploid Brassica napus uncovers key roles for small RNAs. Plant J. 2017, 91, 874–893. [Google Scholar] [CrossRef] [Green Version]
- Birchler, J.A.; Yao, H.; Chudalayandi, S.; Vaiman, D.; Veitia, R.A. Heterosis. Plant Cell 2010, 22, 2105–2112. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Liu, Y.; Zhang, Y.; Gu, R. Advances in research on the mechanism of heterosis in plants. Front. Plant Sci. 2021, 12, 745726. [Google Scholar] [CrossRef]
- Yue, L.; Sun, R.; Li, G.; Cheng, F.; Gao, L.; Wang, Q.; Zhang, S.; Zhang, H.; Zhang, S.; Li, F. Genetic dissection of heterotic loci associated with plant weight by Graded pool-seq in heading Chinese cabbage (Brassica rapa). Planta 2022, 255, 126. [Google Scholar] [CrossRef]
- Choi, S.R.; Yu, X.; Dhandapani, V.; Li, X.; Wang, Z.; Lee, S.Y.; Oh, S.H.; Pang, W.; Ramchiary, N.; Hong, C.P.; et al. Integrated analysis of leaf morphological and color traits in different populations of Chinese cabbage (Brassica rapa ssp. pekinensis). Theor. Appl. Genet. 2017, 130, 1617–1634. [Google Scholar] [CrossRef]
- Sun, X.; Luo, S.; Luo, L.; Wang, X.; Chen, X.; Lu, Y.; Shen, S.; Zhao, J.; Bonnema, G. Genetic analysis of Chinese cabbage reveals correlation between rosette leaf and leafy head variation. Front. Plant Sci. 2018, 9, 1455. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jiang, J.; Ren, A.; Xu, X.; Zhang, H.; Zhao, T.; Jiang, X.; Sun, Y.; Li, J.; Yang, H. Heterosis and combining ability analysis of fruit yield, early maturity, and quality in tomato. Agronomy 2021, 11, 807. [Google Scholar] [CrossRef]
- Geleta, L.F.; Labuschagne, M.T.; Viljoen, C.D. Relationship between heterosis and genetic distance based on morphological traits and AFLP markers in pepper. Plant Breed. 2004, 123, 467–473. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, Z.; Chen, W.; Li, X.; Zhou, S.; Liang, C.; Li, X.; Yang, B.; Zou, X.; Liu, F.; et al. Integration of mRNA and miRNA profiling reveals the heterosis of three hybrid combinations of Capsicum annuum varieties. GM Crops Food 2021, 12, 224–241. [Google Scholar] [CrossRef]
- Naves, E.R.; Scossa, F.; Araújo, W.L.; Nunes-Nesi, A.; Fernie, A.R.; Zsögön, A. Heterosis for capsacinoids accumulation in chili pepper hybrids is dependent on parent-of-origin effect. Sci. Rep. 2022, 12, 14450. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; Prohens, J.; Nuez, F. Performance of hybrids between local varieties of eggplant (Solanum melongena) and its relation to the mean of parents and to morphological and genetic distances among parents. Eur. J. Hortic. Sci. 2008, 73, 76. [Google Scholar]
- Liu, C.; Liu, X.; Han, Y.; Meng, H.; Cheng, Z. Heterosis prediction system based on non-additive genomic prediction models in cucumber (Cucumis sativus L.). Sci. Hortic. 2022, 293, 110677. [Google Scholar] [CrossRef]
- Wu, L. Relationship between SRAP marker based on genetic distance, combining ability and heterosis in pepper. Chin. J. Trop. Crops 2020, 41, 661–668. [Google Scholar]
- Luan, F.; Sheng, Y.; Wang, Y.; Staub, J.E. Performance of melon hybrids derived from parents of diverse geographic origins. Euphytica 2010, 173, 1–16. [Google Scholar] [CrossRef]
- Dafna, A.; Halperin, I.; Oren, E.; Isaacson, T.; Tzuri, G.; Meir, A.; Schaffer, A.A.; Burger, J.; Tadmor, Y.; Buckler, E.S.; et al. Underground heterosis for yield improvement in melon. J. Exp. Bot. 2021, 72, 6205–6218. [Google Scholar] [CrossRef] [PubMed]
- Onofri, A.; Terzaroli, N.; Russi, L. Linear models for diallel crosses: A review with R functions. Theor. Appl. Genet. 2021, 134, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Shajari, M.; Soltani, F.; Bihamta, M.R.; Alabboud, M. Genetic analysis and inheritance of floral and fruit traits in melon (Cucumis melo) in the full diallel cross. Plant Breed. 2021, 140, 486–496. [Google Scholar] [CrossRef]
- Aiswarya, C.S.; Vijeth, S.; Sreelathakumary, I.; Kaushik, P. Diallel analysis of chilli pepper (Capsicum annuum L.) genotypes for morphological and fruit biochemical traits. Plants 2020, 9, 1. [Google Scholar]
- Das, I.; Hazra, P.; Longjam, M.; Bhattacharjee, T.; Maurya, P.K.; Banerjee, S.; Chattopadhyay, A. Genetic control of reproductive and fruit quality traits in crosses involving cultivars and induced mutants of tomato (Solanum lycopersicum L.). J. Genet. 2020, 99, 56. [Google Scholar] [CrossRef]
- Datta, D.R.; Rafii, M.Y.; Misran, A.; Jusoh, M.; Yusuff, O.; Haque, M.A.; Jatto, M.I. Half diallel analysis for biochemical and morphological traits in cultivated eggplants (Solanum melongena L.). Agronomy 2021, 11, 1769. [Google Scholar] [CrossRef]
- Kaur, S.; Sharma, S.P.; Sarao, N.K.; Deol, J.K.; Gill, R.; Abd-Elsalam, K.A.; Alghuthaymi, M.A.; Hassan, M.M.; Chawla, N. Heterosis and combining ability for fruit yield, sweetness, β-Carotene, ascorbic acid, firmness and Fusarium wilt resistance in muskmelon (Cucumis melo L.) involving genetic male sterile lines. Horticulturae 2022, 8, 82. [Google Scholar] [CrossRef]
- Pavan, M.P.; Gangaprasad, S. Studies on mode of gene action for fruit quality characteristics governing shelf life in tomato (Solanum lycopersicum L.). Sci. Hortic. 2022, 293, 110687. [Google Scholar] [CrossRef]
- Semel, Y.; Nissenbaum, J.; Menda, N.; Zinder, M.; Krieger, U.; Issman, N.; Pleban, T.; Lippman, Z.; Gur, A.; Zamir, D. Overdominant quantitative trait loci for yield and fitness in tomato. Proc. Natl. Acad. Sci. USA 2006, 103, 12981–12986. [Google Scholar] [CrossRef] [Green Version]
- Gur, A.; Zamir, D. Unused natural variation can lift yield barriers in plant breeding. PLoS Biol. 2004, 2, e245. [Google Scholar] [CrossRef] [Green Version]
- Gur, A.; Zamir, D. Mendelizing all components of a pyramid of three yield QTL in tomato. Front. Plant Sci. 2015, 6, 1096. [Google Scholar] [CrossRef] [Green Version]
- Yeager, A.F. Determinate growth in the tomato. J. Hered. 1927, 18, 263–265. [Google Scholar] [CrossRef]
- Thouet, J.; Quinet, M.; Ormenese, S.; Kinet, J.M.; Périlleux, C. Revisiting the involvement of SELF-PRUNING in the sympodial growth of tomato. Plant Physiol. 2008, 148, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Liberatore, K.L.; Park, S.J.; Alvarez, J.P.; Lippman, Z.B. Tomato yield heterosis is triggered by a dosage sensitivity of the florigen pathway that fine-tunes shoot architecture. PLoS Genet. 2013, 9, e1004043. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Jiang, K.; Tal, L.; Yichie, Y.; Gar, O.; Zamir, D.; Eshed, Y.; Lippman, Z.B. Optimization of crop productivity in tomato using induced mutations in the florigen pathway. Nat. Genet. 2014, 46, 1337–1342. [Google Scholar] [CrossRef]
- Soyk, S.; Müller, N.A.; Park, S.J.; Schmalenbach, I.; Jiang, K.; Hayama, R.; Zhang, L.; Van Eck, J.; Jiménez-Gómez, J.M.; Lippman, Z.B. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat. Genet. 2017, 49, 162–168. [Google Scholar] [CrossRef]
- Kang, M.S.; Kim, Y.J.; Heo, J.; Rajendran, S.; Wang, X.; Bae, J.H.; Lippman, Z.; Park, S.J. Newly discovered alleles of the tomato antiflorigen gene SELF PRUNING provide a range of plant compactness and yield. Int. J. Mol. Sci. 2022, 23, 7149. [Google Scholar] [CrossRef]
- Díaz, A.; Martín-Hernández, A.M.; Dolcet-Sanjuan, R.; Garcés-Claver, A.; Álvarez, J.M.; Garcia-Mas, J.; Picó, B.; Monforte, A.J. Quantitative trait loci analysis of melon (Cucumis melo L.) domestication-related traits. Theor. Appl. Genet. 2017, 130, 1837–1856. [Google Scholar] [CrossRef]
- Zhao, G.; Lian, Q.; Zhang, Z.; Fu, Q.; He, Y.; Ma, S.; Ruggieri, V.; Monforte, A.J.; Wang, P.; Julca, I.; et al. A comprehensive genome variation map of melon identifies multiple domestication events and loci influencing agronomic traits. Nat. Genet. 2019, 51, 1607–1615. [Google Scholar] [CrossRef]
- Lian, Q.; Fu, Q.; Xu, Y.; Hu, Z.; Zheng, J.; Zhang, A.; He, Y.; Wang, C.; Xu, C.; Chen, B.; et al. QTLs and candidate genes analyses for fruit size under domestication and differentiation in melon (Cucumis melo L.) based on high resolution maps. BMC Plant Biol. 2021, 21, 126. [Google Scholar] [CrossRef]
- Fernández-Silva, I.; Moreno, E.; Eduardo, I.; Arús, P.; Alvarez, J.M.; Monforte, A.J. On the genetic control of heterosis for fruit shape in melon (Cucumis melo L.). J. Hered. 2009, 100, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, H.Y.; Zhou, H.; Mu, H.L.; Wu, S.H.; Jiang, Y.L.; Yang, Z.; Hao, Y.Y.; Zhu, J.; Bao, W.L.; Cheng, S.H.; et al. Integrated analysis of mRNA and non-coding RNA transcriptome in pepper (Capsicum chinense) hybrid at seedling and flowering stages. Front. Genet. 2021, 12, 685788. [Google Scholar] [CrossRef] [PubMed]
- Tonosaki, K.; Fujimoto, R.; Dennis, E.S.; Raboy, V.; Osabe, K. Will epigenetics be a key player in crop breeding? Front. Plant Sci. 2022, 13, 958350. [Google Scholar] [CrossRef] [PubMed]
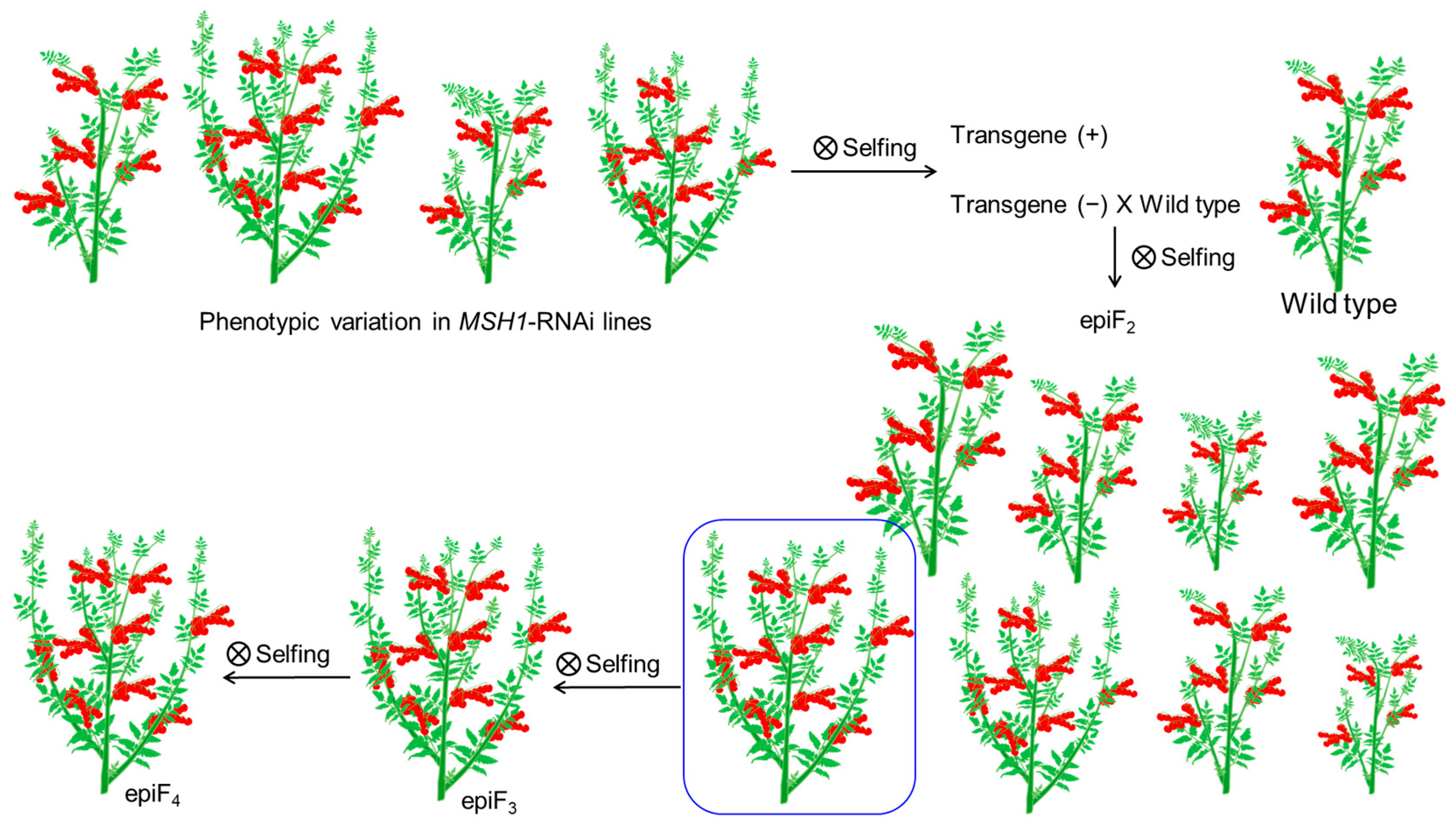
- Yang, X.; Kundariya, H.; Xu, Y.Z.; Sandhu, A.; Yu, J.; Hutton, S.F.; Zhang, M.; Mackenzie, S.A. MutS HOMOLOG1-derived epigenetic breeding potential in tomato. Plant Physiol. 2015, 168, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagosz, B. The relationship between heterosis and genetic distances based on RAPD and AFLP markers in carrot. Plant Breed. 2011, 130, 574–579. [Google Scholar] [CrossRef]
- Turner, S.D.; Maurizio, P.L.; Valdar, W.; Yandell, B.S.; Simon, P.W. Dissecting the genetic architecture of shoot growth in carrot (Daucus carota L.) using a diallel mating design. G3 2018, 8, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Yi, X.; Wang, Q.; Wang, J.; Wang, S.; Liu, L.; Xu, L.; Wang, Y. Classification and identification of S haplotypes in radish based on SRK kinase domain sequence analysis. Plants 2022, 11, 2304. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, Z.; Tang, D.; Zhu, Y.; Wang, P.; Li, D.; Zhu, G.; Xiong, X.; Shang, Y.; Li, C.; et al. Genome design of hybrid potato. Cell 2021, 184, 3873–3883. [Google Scholar] [CrossRef]
- Li, D.; Lu, X.; Zhu, Y.; Pan, J.; Zhou, S.; Zhang, X.; Zhu, G.; Shang, Y.; Huang, S.; Zhang, C. The multi-omics basis of potato heterosis. J. Integr. Plant Biol. 2022, 64, 671–687. [Google Scholar] [CrossRef]
- Fujimoto, R.; Taylor, J.M.; Sasaki, T.; Kawanabe, T.; Dennis, E.S. Genome wide gene expression in artificially synthesized amphidiploids of Arabidopsis. Plant Mol. Biol. 2011, 77, 419–431. [Google Scholar] [CrossRef]
- Tonosaki, K.; Osabe, K.; Kawanabe, T.; Fujimoto, R. The importance of reproductive barriers and the effect of allopolyploidization on crop breeding. Breed. Sci. 2016, 66, 333–349. [Google Scholar] [CrossRef] [Green Version]
- Mehraj, H.; Akter, A.; Miyaji, N.; Miyazaki, J.; Shea, D.J.; Fujimoto, R.; Doullah, M.A. Genetics of clubroot and Fusarium wilt disease resistance in Brassica vegetables: The application of marker assisted breeding for disease resistance. Plants 2020, 9, 726. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiraki, S.; Fujiwara, K.; Kamiya, Y.; Akter, M.A.; Dennis, E.S.; Fujimoto, R.; Mehraj, H. Studies on the Molecular Basis of Heterosis in Arabidopsis thaliana and Vegetable Crops. Horticulturae 2023, 9, 366. https://doi.org/10.3390/horticulturae9030366
Shiraki S, Fujiwara K, Kamiya Y, Akter MA, Dennis ES, Fujimoto R, Mehraj H. Studies on the Molecular Basis of Heterosis in Arabidopsis thaliana and Vegetable Crops. Horticulturae. 2023; 9(3):366. https://doi.org/10.3390/horticulturae9030366
Chicago/Turabian StyleShiraki, Saaya, Kazumasa Fujiwara, Yoshiki Kamiya, Mst. Arjina Akter, Elizabeth S. Dennis, Ryo Fujimoto, and Hasan Mehraj. 2023. "Studies on the Molecular Basis of Heterosis in Arabidopsis thaliana and Vegetable Crops" Horticulturae 9, no. 3: 366. https://doi.org/10.3390/horticulturae9030366