


Effects of Auxin-Type Plant Growth Regulators and Cold Stress on the Endogenous Polyamines in Pea Plants

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Content of Oxidative Stress Biomarkers MDA and Hydrogen Peroxide (H2O2)
2.3. Determination of Proline Content
2.4. Determination of Total Phenolics Content
2.5. Analyses of Polyamine Content
2.6. Analyses of Chlorophyll and Carotenoids Contents
2.7. Statistical Analysis
3. Results
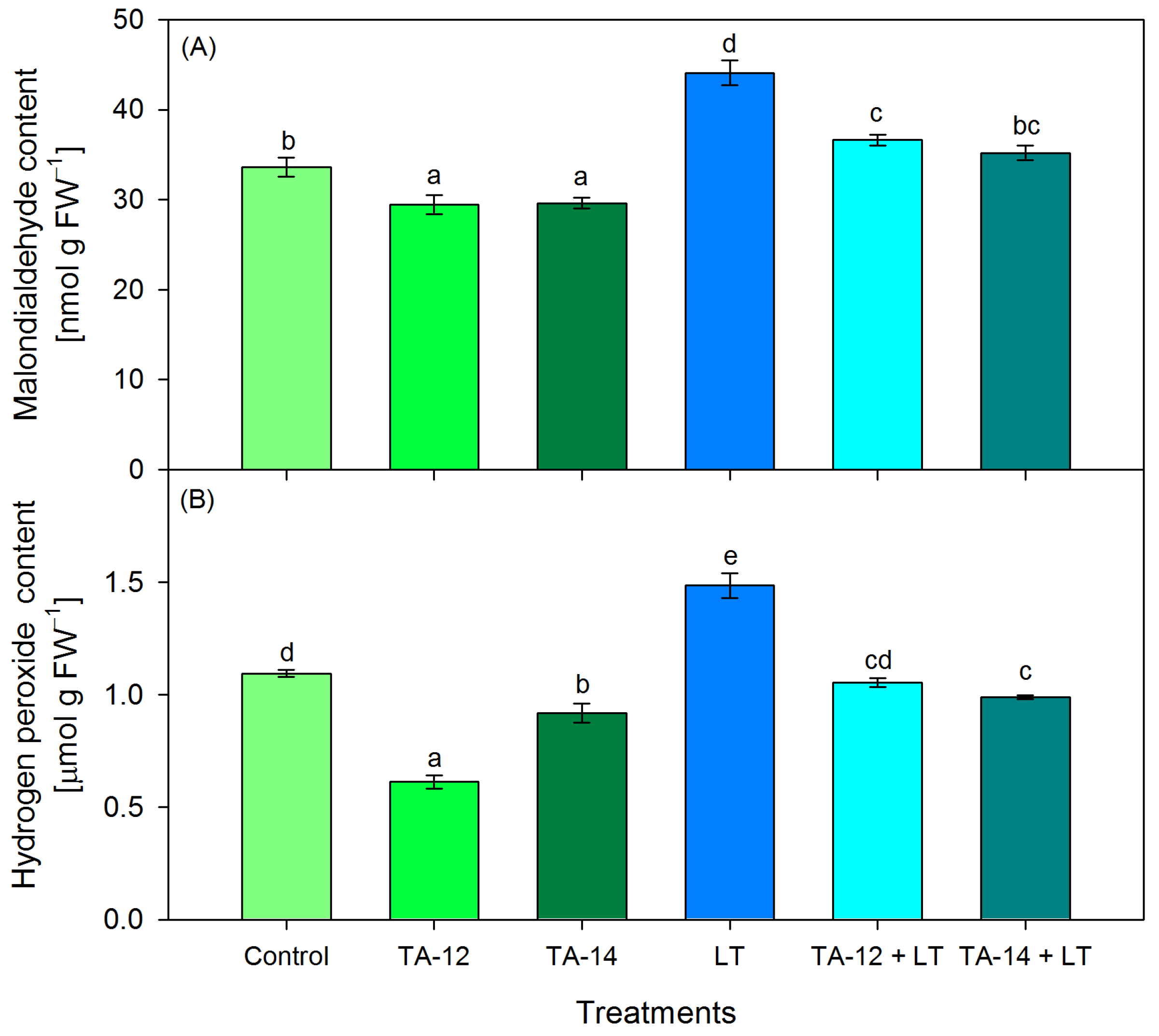
3.1. Content of Oxidative Stress Markers Malondialdehyde and Hydrogen Peroxide
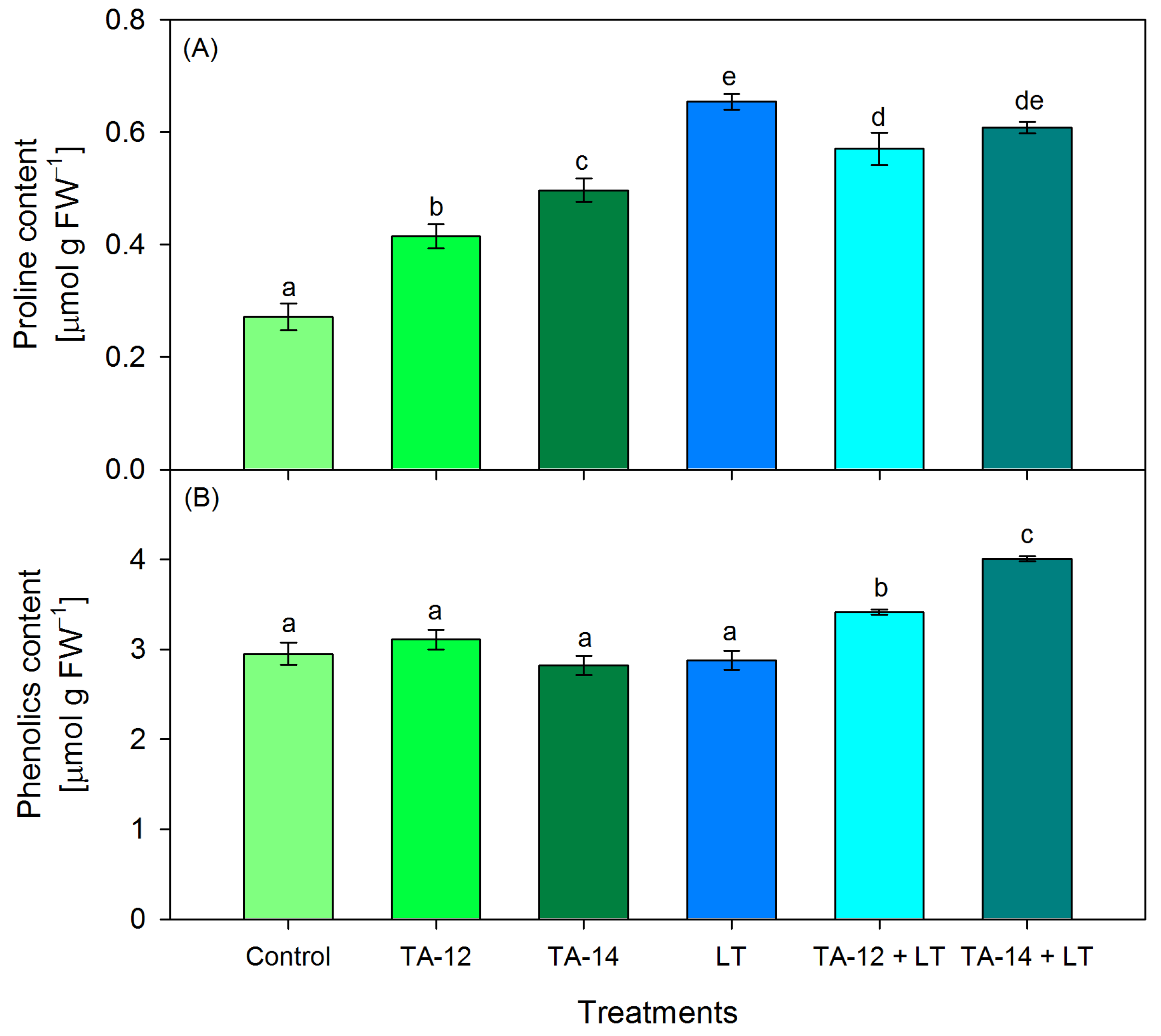
3.2. Content of Proline and Total Phenolics
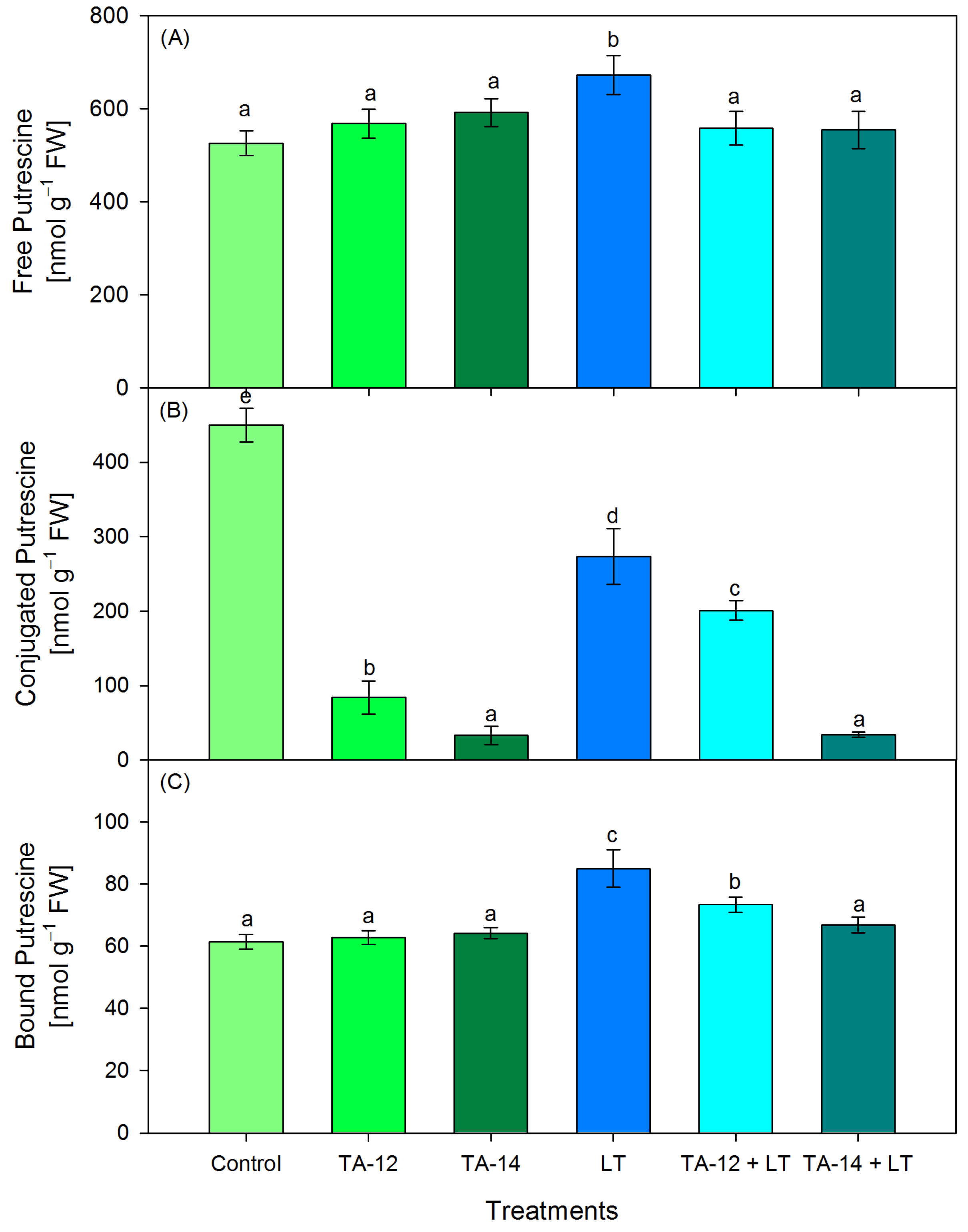
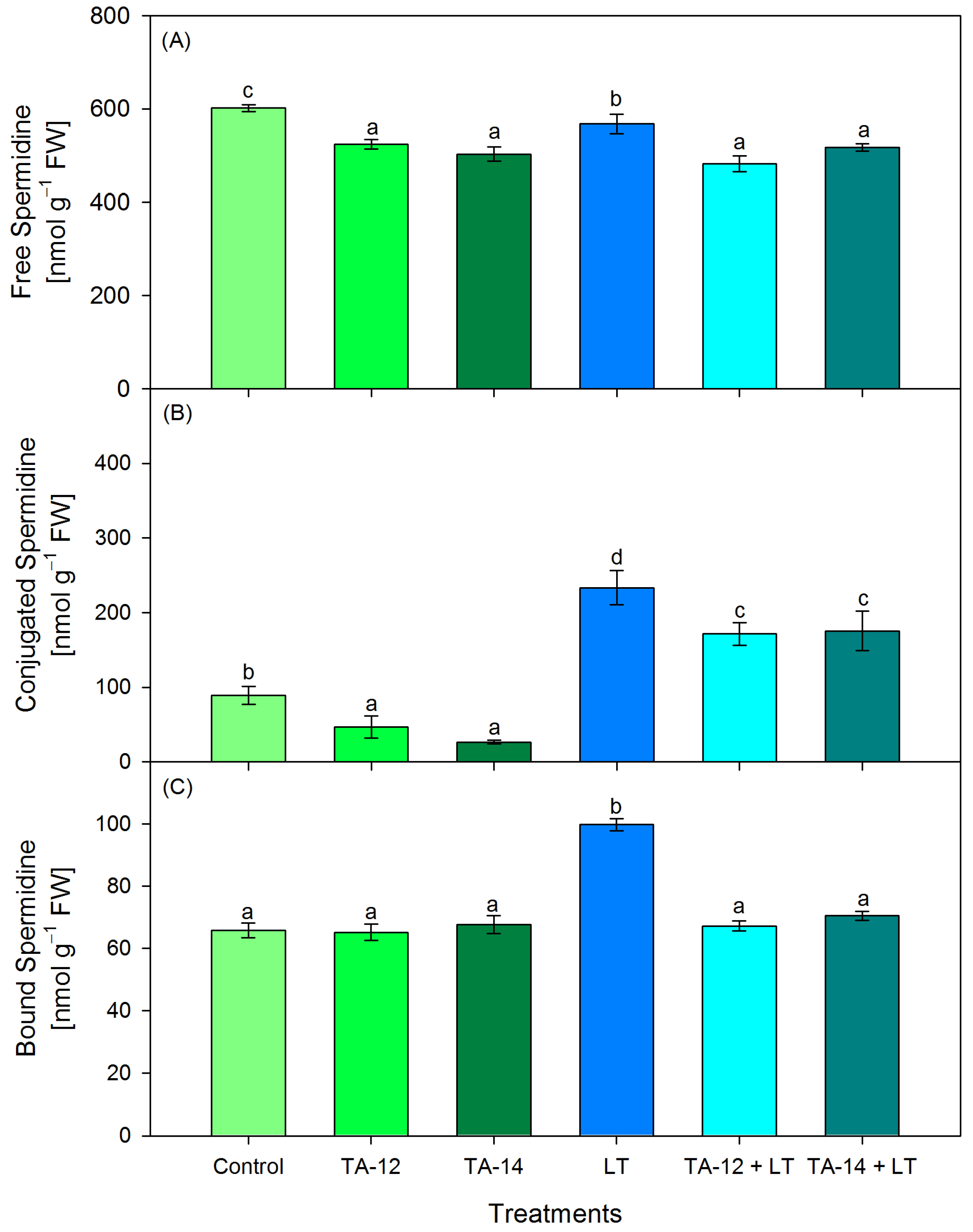
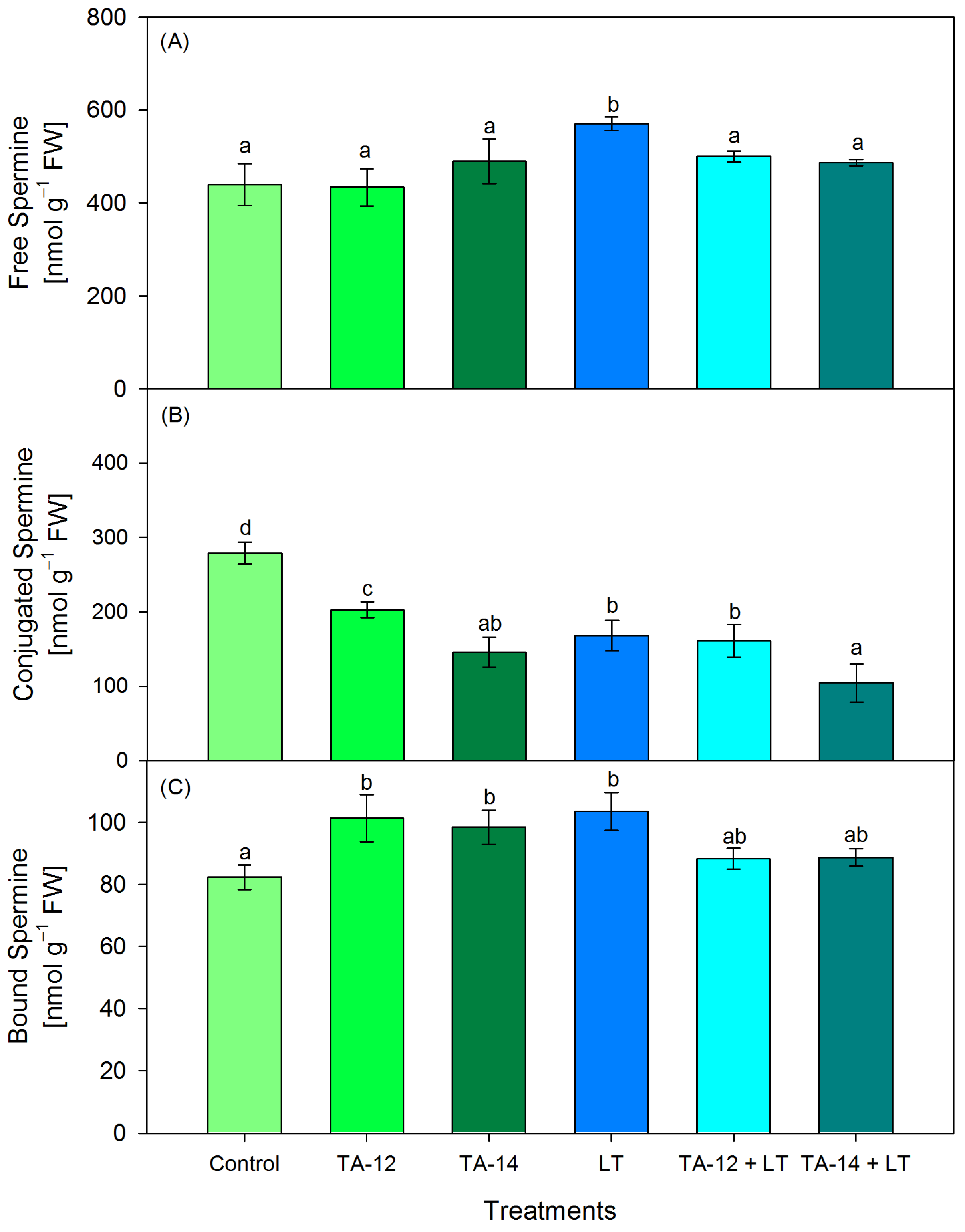
3.3. Content of Free, Conjugated and Bound PAs (Putrescine, Spermidine and Spermine)
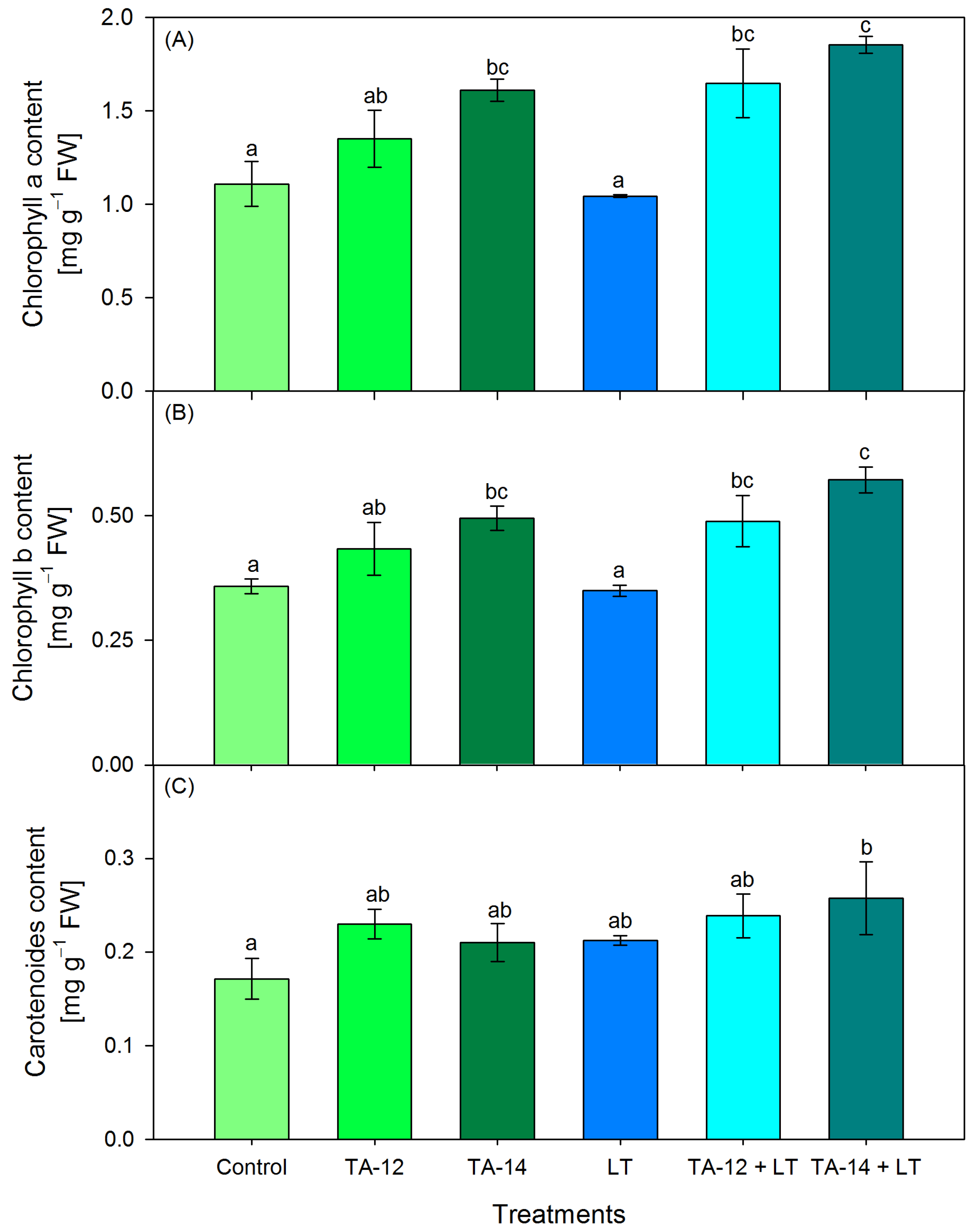
3.4. Content of Photosynthetic Pigments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dahl, W.J.; Foster, L.M.; Tyler, R.T. Review of the health benefits of peas (Pisum sativum L.). Brit. J. Nutr. 2012, 108, S3–S10. [Google Scholar] [CrossRef]
- Balliu, A.; Sallaku, G. The environment temperature affects post-germination growth and root system architecture of pea (Pisum sativum L.) plants. Sci. Horticult. 2021, 278, 109858. [Google Scholar] [CrossRef]
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.; Qin, Y. Plant Low-Temperature Stress: Signaling and Response. Agronomy 2022, 12, 702. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: The Physical Science Basis; Contribution of Working Group I to the Forth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S.D., Qin, D., Manning, M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Niu, S.; Luo, Y.; Li, D.; Cao, S.; Xia, J.; Li, J.; Smith, M.D. Plant growth and mortality under climatic extremes: An overview. Envir. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Younis, A.; Ramzan, F.; Ramzan, Y.; Zulfiqar, F.; Ahsan, M.; Lim, K.B. Molecular Markers Improve Abiotic Stress Tolerance in Crops: A Review. Plants 2020, 9, 1374. [Google Scholar] [CrossRef]
- Liu, R.; Fang, L.; Yang, T.; Zhang, X.; Hu, J.; Zhang, H.; Han, W.; Hua, Z.; Hao, J.; Zong, X. Marker-trait association analysis of frost tolerance of 672 worldwide pea (Pisum sativum L.) collections. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotech. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 2021, 105, 459–476. [Google Scholar] [CrossRef]
- Shi, Y.; Ding, Y.; Yang, S. Cold signal transduction and its interplay with phytohormones during cold acclimation. Plant Cell Physiol. 2015, 56, 7–15. [Google Scholar] [CrossRef]
- Jangra, A.; Chaturvedi, S.; Kumar, N.; Singh, H.; Sharma, V.; Thakur, M.; Tiwari, S.; Chhokar, V. Polyamines: The Gleam of Next-Generation Plant Growth Regulators for Growth, Development, Stress Mitigation, and Hormonal Crosstalk in Plants—A Systematic Review. J. Plant Growth Regul. 2022, 1–25. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Gondor, O.K.; Janda, T. Unfinished story of polyamines: Role of conjugation, transport and light-related regulation in the polyamine metabolism in plants. Plant Sci. 2021, 308, 110923. [Google Scholar] [CrossRef]
- Jankovska-Bortkevič, E.; Gavelienė, V.; Jurkonienė, S. Physiological roles and signaling of polyamines in plants under stressed conditions. In Emerging Plant Growth Regulators in Agriculture; Naeem, M., Aftab, T., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 303–316. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, S.; Zulfiqar, F.; Alam, M.; Fujita, M. Regulation of ROS metabolism in plants under environmental stress: A review of recent experimental evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Balestrini, R.; Chitarra, W.; Marsico, A.D.; Nerva, L. Getting ready with the priming: Innovative weapons against biotic and abiotic crop enemies in a global changing scenario. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Hossain, M.A., Liu, F., Burritt, D., Fujita, M., Huang, B., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 35–56. [Google Scholar]
- Llorens, E.; González-Hernández, A.I.; Scalschi, L.; Fernández-Crespo, E.; Camañes, G.; Vicedo, B.; García-Agustín, P. Priming mediated stress and cross-stress tolerance in plants: Concepts and opportunities. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Hossain, M.A., Liu, F., Burritt, D., Fujita, M., Huang, B., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–20. [Google Scholar]
- Guo, J.; Liu, S.; Li, X.; Liu, F. Crop exposure to cold stress: Responses in physiological, biochemical and molecular levels. In Sustainable Crop Productivity and Quality Under Climate Change; Liu, F., Li, X., Hogy, P., Jiang, D., Brestic, M., Liu, B., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 1–19. [Google Scholar]
- Anisimovienė, N.; Jankauskienė, J.; Novickienė, L. Actualities in plant cold acclimation. Scientific works of the Institute of Horticulture. Lithuanian Research Centre for Agriculture and Forestry and Lithuanian University of Agriculture. Sodinink. Ir Daržinink. 2008, 27, 99–109. [Google Scholar]
- Gavelienė, V.; Novickienė, L.; Pakalniškytė, L. Effect of auxin physiological analogues on rapeseed (Brassica napus) cold hardening, seed yield and quality. J. Plant Res. 2013, 126, 283–292. [Google Scholar] [CrossRef]
- Jankauskienė, J.; Mockevičiūtė, R.; Gavelienė, V.; Jurkonienė, S.; Anisimovienė, N. The Application of Auxin-like Compounds Promotes Cold Acclimation in the Oilseed Rape Plant. Life 2022, 12, 1283. [Google Scholar] [CrossRef] [PubMed]
- Sergiev, I.; Todorova, D.; Shopova, E.; Jankauskienė, J.; Jankovska-Bortkevič, E.; Jurkonienė, S. Effects of auxin analogues and heat stress on garden pea. Zemdirb.-Agricult. 2018, 105, 243–248. [Google Scholar] [CrossRef]
- Sergiev, I.; Todorova, D.; Shopova, E.; Brankova, L.; Jankauskienė, J.; Jurkonienė, S.; Gavelienė, V.; Mockevičiūtė, R. Assessment of synthetic auxin-type compounds as potential modulators of herbicide action in Pisum sativum L. Biologia 2020, 75, 1845–1853. [Google Scholar] [CrossRef]
- Todorova, D.; Sergiev, I.; Shopova, E.; Brankova, L.; Jankauskienė, J.; Jurkonienė, S.; Gavelienė, V.; Mockevičiūtė, R. Physiological responses of Pea plants to treatment with synthetic auxins and auxin-type herbicide. Botanica 2021, 27, 125–133. [Google Scholar] [CrossRef]
- Sergiev, I.; Todorova, D.; Shopova, E.; Jankauskienė, J.; Jankovska-Bortkevič, E.; Jurkonienė, S. Exogenous auxin type compounds amend PEG-induced physiological responses of pea plants. Sci. Horticult. 2019, 248, 200–205. [Google Scholar] [CrossRef]
- Todorova, D.; Katerova, Z.; Shopova, E.; Brankova, L.; Sergiev, I.; Jankauskienė, J.; Jurkonienė, S. The Physiological Responses of Wheat and Maize Seedlings Grown under Water Deficit Are Modulated by Pre-Application of Auxin-Type Plant Growth Regulators. Plants 2022, 11, 3251. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.; Norman, H.; Krizek, D.; Mirecki, R. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Swain, T.; Goldstein, L. Methods in Polyphenol Chemistry; Pridham, J.B., Ed.; Pergamon Press: Oxford, UK, 1964; pp. 131–146. [Google Scholar]
- Smith, T.; Best, G. Polyamines in barley seedlings. Phytochem. 1977, 16, 841–843. [Google Scholar] [CrossRef]
- Torrigiani, P.; Altamura, M.; Copitani, F.; Serafini-Fracasini, D.; Bagni, N. De novo root formation in thin cell layers of tobacco: Changes in free and bound polyamines. Physiol. Plant. 1989, 77, 294–301. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophyls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef]
- Generozova, I.P.; Butsanets, P.A.; Shugaev, A.G. Mitochondrial respiration after combined action of dehydration and low temperature in pea seedlings. Biol. Plant. 2019, 63, 11–19. [Google Scholar] [CrossRef]
- Tang, X.; An, B.; Cao, D.; Xu, R.; Wang, S.; Zhang, Z.; Liu, X.; Sun, X. Improving photosynthetic capacity, alleviating photosynthetic inhibition and oxidative stress under low temperature stress with exogenous hydrogen sulfide in blueberry seedlings. Front. Plant Sci. 2020, 11, 108. [Google Scholar] [CrossRef]
- Xu, C.; Yang, Z.Q.; Yang, S.Q.; Wang, L.; Wang, M.T. High humidity alleviates photosynthetic inhibition and oxidative damage of tomato seedlings under heat stress. Photosynthetica 2020, 58, 146–155. [Google Scholar] [CrossRef]
- Bhatt, U.; Sharma, S.; Soni, V. Differential photosynthetic responses in Riccia gangetica under heat, cold, salinity, submergence, and UV-B stresses. J. Photochem. Photobiol. 2022, 12, 100146. [Google Scholar] [CrossRef]
- Nasibi, F.; Kalantari, K.M.; Tavakoli, Z.M. Effects of Hydrogen Sulfide on Cold-Induced Oxidative Damage in Cucumis sativus L. Int. J. Horticult. Sci. Technol. 2020, 7, 199–211. [Google Scholar]
- Kishor, P.B.K.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homoeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.; Browse, J. Cold comfort farm: The acclimation of plants to freezing temperatures. Plant Cell Environ. 2000, 23, 893–902. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Król, A.; Amarowicz, R.; Weidner, S. The effects of cold stress on the phenolic compounds and antioxidant capacity of grapevine (Vitis vinifera L.) leaves. J. Plant Physiol. 2015, 189, 97–104. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Kasukabe, Y.; He, L.; Nada, K.; Misawa, S.; Ihara, I.; Tachibana, S. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stressregulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 712–722. [Google Scholar] [CrossRef]
- Shen, W.; Nada, K.; Tachibana, S. Involvement of polyamines in the chilling tolerance of cucumber cultivars. Plant Physiol. 2000, 124, 431–439. [Google Scholar] [CrossRef]
- Ha, H.C.; Sirisoma, N.S.; Kuppusamy, P.; Zweiler, J.L.; Woster, P.M.; Casero, R.A., Jr. The natural polyamine spermine functions directly as a free scavenger. Proc. Nat. Acad. Sci. USA 1998, 95, 11140–11145. [Google Scholar] [CrossRef]
- Garbero, M.; Andrade, A.; Reinoso, H.; Fernández, B.; Cuesta, C.; Granda, V.; Escudero, C.; Abdala, G.; Pedranzani, H. Differential effect of short-term cold stress on growth, anatomy, and hormone levels in cold-sensitive versus-resistant cultivars of Digitaria eriantha. Acta Physiol. plant. 2012, 34, 2079–2091. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Knirsch, V.; Jarosova, J.; Gaudinova, A.; Zupkova, B.; Prášil, I.T.; Janda, T.; Brzobohatý, B.; Skalák, J.; et al. Light Quality and Intensity Modulate Cold Acclimation in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 2736. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Tang, G.; Zhu, X.; Gakiere, B. Lysine catabolism: A stress and development super-regulated metabolic pathway. Curr. Opin. Plant Biol. 2001, 4, 261–266. [Google Scholar] [CrossRef]
- Mir, A.R.; Siddiqui, H.; Alam, P.; Hayat, S. Foliar spray of Auxin/IAA modulates photosynthesis, elemental composition, ROS localization and antioxidant machinery to promote growth of Brassica juncea. Physiol. Mol. Biol. Plants 2020, 26, 2503–2520. [Google Scholar] [CrossRef]
- Ahmad, A.; Hayat, S.; Fariduddin, Q.; Ahmad, I. Photosynthetic efficiency of plants of Brassica juncea, treated with chlorosubstituted auxins. Photosynthetica 2001, 39, 565–568. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, A.; Rathinasabapathi, B. Oxidative stress tolerance in plants: Novel interplay between auxin and reactive oxygen species signaling. Plant Signal. Behavior. 2013, 8, e25761. [Google Scholar] [CrossRef]
- Zhai, S.; Xia, X.; He, Z. Carotenoids in staple cereals: Metabolism, regulation, and genetic manipulation. Front. Plant Sci. 2016, 7, 227. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Total Put, nmol g–1 FW | Total Spd, nmol g–1 FW | Total Spm, nmol g–1 FW |
|---|---|---|---|
| Control | 1037 (100 *) | 756 (100) | 800 (100) |
| TA-12 | 750 (72) | 636 (84) | 738 (92) |
| TA-14 | 689 (66) | 597 (79) | 734 (92) |
| LT | 1230 (119) | 930 (123) | 842 (105) |
| TA-12 + LT | 853 (82) | 721 (95) | 750 (94) |
| TA-14 + LT | 655 (63) | 763 (101) | 680 (85) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankovska-Bortkevič, E.; Katerova, Z.; Todorova, D.; Jankauskienė, J.; Mockevičiūtė, R.; Sergiev, I.; Jurkonienė, S. Effects of Auxin-Type Plant Growth Regulators and Cold Stress on the Endogenous Polyamines in Pea Plants. Horticulturae 2023, 9, 244. https://doi.org/10.3390/horticulturae9020244
Jankovska-Bortkevič E, Katerova Z, Todorova D, Jankauskienė J, Mockevičiūtė R, Sergiev I, Jurkonienė S. Effects of Auxin-Type Plant Growth Regulators and Cold Stress on the Endogenous Polyamines in Pea Plants. Horticulturae. 2023; 9(2):244. https://doi.org/10.3390/horticulturae9020244
Chicago/Turabian StyleJankovska-Bortkevič, Elžbieta, Zornitsa Katerova, Dessislava Todorova, Jurga Jankauskienė, Rima Mockevičiūtė, Iskren Sergiev, and Sigita Jurkonienė. 2023. "Effects of Auxin-Type Plant Growth Regulators and Cold Stress on the Endogenous Polyamines in Pea Plants" Horticulturae 9, no. 2: 244. https://doi.org/10.3390/horticulturae9020244