
Insights into the Key Genes in Cucumis melo and Cucurbita moschata ToLCNDV Resistance


1
Institute for Research and Training in Agriculture and Fisheries, IFAPA Alameda del Obispo, Avda. Menéndez Pidal s/n, 14004 Córdoba, Spain
2
Institute for Research and Training in Agriculture and Fisheries, IFAPA La Mojonera, Camino San Nicolás, 1, 04745 Almeria, Spain
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(2), 231; https://doi.org/10.3390/horticulturae9020231
Submission received: 18 November 2022
/
Revised: 3 February 2023
/
Accepted: 5 February 2023
/
Published: 8 February 2023
(This article belongs to the Special Issue Plant Disease Management: Latest Advances and Prospects)
Abstract
:Tomato leaf curl New Delhi virus (ToLCNDV, family Geminiviridae, genus Begomovirus) is a whitefly-transmitted virus that causes widespread damage in Cucurbitaceae and Solaneceae crops worldwide. The Spanish strain, ToLCNDV-ES, affects mainly cucurbit crops and has spread through the Mediterranean basin since its first detection in 2013 in the south of Spain. The control of the virus has been based on the adoption of measures to control the vector, which have not been sufficient to reduce production losses. Therefore, the identification of key genes for ToLCNDV resistance is essential for the development of resistant plants. Regarding genetic control of resistance in cucurbit crops, one major locus on chromosome 11 and two additional regions in chromosomes 12 and 2 of C. melo linked to ToLCNDV resistance have been described recently. Concerning C. moschata, a major QTL was also identified on chromosome 8 that resulted in synteny with a QTL on chromosome 11 of C. melo. In this work, we investigated the molecular basis of ToLCNDV resistance in contrasting accessions of C. melo and C. moschata by transcriptional characterization of 10 different candidate genes controlling host factors related to proviral or antiviral mechanisms. Two proviral factor genes, ARP4 in C. melo and SYTA in C. moschata, showed clear differences in expression levels when the susceptible and resistant accessions were compared. The knowledge of proviral factors associated with resistance could be used to screen an active mutagenesis TILLING platform. This is the case of C. pepo, in which no ToLCNDV resistance has been described to date. The relationship between the regulation of the genes ARP4 and SYTA, as well as the genome position of the described loci related to ToLCNDV resistance, is also discussed.
1. Introduction
Viral plant diseases account for nearly half of all emerging plant diseases worldwide, causing damage to both commercial crops and natural vegetation [1]. Tomato leaf curl New Delhi virus (ToLCNDV), a member of the genus Begomovirus in the family Geminiviridae, is a limiting factor for cucurbit cultivation. Originally, the virus was described in India [2] as causing significant damage to tomato crops, but later the virus spread to other, nearby Asian countries, affecting other hosts, mainly species of the Cucurbitaceae and Solanaceae families. ToLCNDV was confined to the Asian continent until it emerged in 2013 in southeastern Spain [3]. The strain described in Spain, named ToLCNDV-ES, caused great damage mainly to zucchini, melon, and cucumber crops in the southeastern area of the country, but it is also capable of infecting other Solanaceae crops such as tomato [4,5,6].
The most effective way to control plant viruses is the development and use of resistant cultivars by means of genetic breeding. The identification of sources of resistance and molecular mechanisms underlying the trait is required for the development of improved material through molecular breeding. Regarding resistance sources, different accessions have been reported as ToLCNDV-resistant in Cucumis melo [7,8] and Cucurbita moschata [9]. Nevertheless, there has been no resistant material reported so far in Cucurbita pepo, although zucchini is severely affected by the virus [3,10]. In this sense, the variability found in the genus Cucurbita could assist ToLCNDV resistance introgression in the species.
To unravel the molecular mechanisms underlying plant virus resistance, the basis of the active dialogue between both partners should be determined. This dialogue is not always hostile, since some plant factors can help the virus (with a limited capacity) complete its vital cycle. As a result, depending on whether they are proviral or antiviral, host factors can either help or hinder the pathogen. Proviral factors determine plant virus susceptibility by participating in virus life cycle processes [11], whereas antiviral factors are activated in response to virus infection [12]. Different host genes involved in plant virus resistance, comprising both categories, have been reported. Considering proviral factors, actin filaments facilitate the transport of viral movement proteins in different plant virus families [13], and an actin-related protein, ARP4, has been recently proposed as being involved in ToLCNDV melon resistance [14]. Moreover, there is evidence associating microtubule disruption with plant defense responses against pathogens [15]. Microtubules act as a dynamic scaffold for the development of viral replicate complexes in the plant cell [13]. In this context, the role of the TORTIFOLIA protein in the microtubule dynamics related to plant virus spread has also been described [16]. Another gene related to virus movement is the synaptotagmin SYTA, reported as a regulator of cell-to-cell movement of diverse plant viruses [17]. In the case of the replication and translation of plant viruses, proviral factors have also been described. Although the most reported factors essential for RNA virus infection have been eIF4 and eIF4G, which are involved in the initiation phase of translation [18], in the case of DNA viruses, factor PELOTA seems to be needed in the recycling of the ribosome phase [19,20]. In this sense, this gene has been shown to be critical in the resistance of tomato against the begomovirus TYLCV in the last phase of protein biosynthesis of the translation machinery [19]. According to antiviral factors, genomic regions related to gene silencing or viral protein recognition have also been reported as relevant in plant virus resistance. RNA silencing is a nucleotide sequence-specific process that induces mRNA degradation or translation inhibition at the post-transcriptional level [21]. The role of RDR (RNA-dependent RNA polymerase), AGO (Argonaute) and SGS (Suppressor of gene silencing) genes has been studied in tomato against DNA begomoviruses and specifically ToLCNDV [22,23,24]. Regarding viral protein recognition, the nuclear shuttle protein (NSP) of ToLCNDV induces a hypersensitive response in tobacco and tomato plants [25]. The strategy of plant antiviral defense involving this type of viral protein is based on the suppression of the host global translation, which is mediated by the transmembrane immune receptor NIK1 (nuclear shuttle protein NSP-Interacting Kinase) [26].
Recent studies have located the genetic regions controlling ToLCNDV resistance in cucurbits on genetic maps. One major quantitative trait locus (QTL) in chromosome 11 and two additional regions in chromosomes 12 and 2 have been reported in C. melo [8]. According to C. moschata, a major QTL was identified in chromosome 8 that resulted in synteny with QTL in chromosome 11 of C. melo and chromosome 17 of C. pepo [27]. The aim of this study was to gain insight into the molecular basis of ToLCNDV resistance by determining the expression profile of 10 selected genes among contrasting accessions of C. melo and C. moschata of 10 different candidate genes controlling host factors related to proviral (virus movement and replication) or antiviral (gene silencing and viral protein recognition) mechanisms. The genes with the most relevant responses as well as their regulation-associated factors were discussed in relation to the QTLs already reported. In the case of melon, this study will widen the resistance sources analyzed in a previous work [14] with the analysis of new potential candidate genes. In the case of pumpkin, the identified genetic variants involved in incompatible interactions could be used to screen an available and active mutagenesis TILLING platform of C. pepo [28] species with no ToLCNDV resistance described to date.
2. Materials and Methods
2.1. Plant Material, Whitefly and Virus Inoculation
Three accessions of C. melo and C. moschata, contrasting for ToLCNDV resistance, were used to evaluate the mRNA response to the virus stress. The melon accessions used in this project were C. melo subsp. agrestis group Momordica PI 414723 (Mom-PI414Ind) and PI 124112 (Mom-PI 124Ind), kindly supplied by USDA-NPGS and described as resistant to ToLCNDV [7], and the susceptible C. melo subsp. melo group Inodorus cv. Piel de Sapo. In the case of C. moschata, the selected accessions were PI 604506 and PI 381814, previously reported as symptomless or with slight symptoms after whitefly and sap inoculation with ToLCNDV [9]. PI 604506 is the improved pumpkin cultivar Large Cheese from the United States, and PI 381814 is an Indian landrace. The susceptible C. moschata genotype studied was the accession PI 419083.
2.2. Virus Inoculation Experiment
2.2.1. Virus Inoculation
Groups of 20 ToLCNDV-infected whiteflies were transferred to 16 resistant and 16 susceptible plants per species (C. melo and C. moschata) at the second true-leaf stage for a 24 h inoculation access period (IAP). The plants were sprayed with imidacloprid at a concentration of 20 mg/L the next day to kill the insects and then kept in insect cages. The same number of resistant and susceptible plants were maintained in separate insect cages as uninfected controls. Leaf samples from four plants of each genotype, inoculated or not, were collected 0 days post infection (dpi), 3 dpi, 6 dpi, 10 dpi, and 15 dpi. The samples were stored at −80 °C until they could be used, as described below.
2.2.2. Quantitative Virus Detection
The relative concentrations of ToLCNDV in C. melo and C. moschata accessions were assessed by qPCR. Total DNA and RNA were extracted using AllPrep DNA/RNA Mini Kit (Quiagen, Madrid, Spain). The extraction was diluted into a final concentration of 50 ng/μL. Three biological replicates were considered. Each replicate was an independent extraction from pools of four plants as described below. qPCR reactions were carry out in 96-well reaction plates using TaqMan PCR Master Mix (Applied Biosystems, Carlsbad, CA, USA). The TaqMan PCR conditions, primers, probes to detect ToLCNDV and the internal control base on the 18S ribosomal RNA gene were as described in [14]. Three technical replicates were performed per sample, and the tests were run on the ABI Prism 7000 DNA sequence detection system (Applied Biosystems, Carlsbad, CA, USA). The relative accumulation of the virus to a calibrator sample was calculated using the formula 2−ΔΔCt, where ΔΔCt is the difference between the ΔCt of each sample and the ΔCt of the calibrator sample [9,30].
2.3. Expression Analysis
2.3.1. RNA Isolation and cDNA Synthesis
Leaf samples from four different plants were collected from each treatment, mixed separately by species and genotype, replicated three times, and immediately immersed in liquid nitrogen before being stored at −80 °C for RNA isolation. Each replicate was an independent RNA extraction from a pool of four plants to minimize the variation in gene expression between individual plants. Total RNA was extracted using TRIsure (Bioline, London, UK) as a reagent, according to the manufacturer’s protocol. The concentrations and purity of extracted RNA were quantified with an ND-2000c Spectrophotometer (Eppendorf, Hamburg, Germany). For cDNA synthesis, only RNA samples with 260/280 ratios between 1.9 and 2.1 and 260/230 ratios greater than 2.0 were used. Reverse transcription of 1 μg of total RNA to cDNA was synthetized using the QuantiTec Reverse Transcription Kit (Qiagen, Hilden, Germany) with a blend of oligo-dT and random primers following the manufacturer’s instructions. Each reaction contained a negative control to detect potential genomic DNA contamination.
2.3.2. Gene Expression Analysis
A set of 10 candidate genes were selected for their previous implications in virus resistance: ARP4 [14,31,32], TORT [16], SYTA [17,33,34], PELOTA [19,20], SGS1 and SGS2 [24], AGO1 [22,35], RDR1 [36,37], RDR6 [23], and NIK1 [38].
EST sequences from the DHL92 C. melo and Rifu C. moschata transcriptome databases at the Cucurbit Genomics Database (www.icugi.org) were used to design specific primer pairs. Primer design considering homologous sequences was carried out using Primer3web (version 4.1.0) online software (http://primer3.ut.ee). The pair of primers designed for each candidate gene amplified a fragment out 100 bp-length fragments at the optimal melting temperature (Tm) of 60 °C with GC contents between 35 and 65% (Table 1). A CFX Connect real-time PCR system (Bio-Rad Laboratories, USA) was used for quantitative PCR (qPCR) using SYBR Green technology. In each qPCR reaction, 300 nM of each gene-specific primer, 1.5 μL of cDNA sample (≈10 ng of input RNA), and 2× iTaq Fast SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) were reacted as follows: 3 min at 95 °C, 40 cycles of 30 s at 95 °C and 30 s at 60 °C. Eventually, a dissociation analysis of the PCR products was carried out by running a gradient from 65 to 95 °C to confirm the presence of a single PCR product. Two technical replicates were performed for each sample and a negative control using water as a template.
2.3.3. Data Analysis
Using LinReg software and the criteria of including three–five fluorescent data points with R2 ≥ 0.998 to define a linear regression line [39], the amplification efficiency of each primer pair was estimated from the exponential phase data of each individual amplification plot and the equation (1 + E) = 10slope. Expression levels of the target genes were calculated using the advanced relative quantification model with efficiency correction, multiple reference gene normalization, and error propagation rules [40]. ADP (ADP ribosylation factor 1) and RAN (GTP-binding nuclear protein) genes were included as housekeeping genes for C. melo and C. moschata [41]. The standard error was calculated using three biological replicates. The values of the susceptible and resistant samples considered in each time point (three replicates) were used in a Student’s t test to calculate probabilities of different response of the gene in the susceptible sample against each of the two resistant accessions per species.
2.4. cDNA Sequencing
The most relevant genes in terms of expression were sequenced. ARP4 in the case of C. melo and SYTA in the case of C. moschata were sequenced in the cDNAs of each of the three accessions per species. Fragments were PCR amplified in a Bio-Rad MyCycler Thermal Cycler PCR using a set of primers designed for each species (three primer pairs for each accession and species) with Primer3web (version 4.1.0) online software (http://primer3.ut.ee). cDNA amplification was carried out in triplicate in total volumes of 25 μL containing 40 ng of cDNA, 0.5 μM of each primer, 2.5 μL of 10× NH4 Reaction Buffer, 0.75 μL of 50 mM MgCl2 solution, 0.2 mM of the dNTP mixture, and 1.25 unit of Taq DNA polymerase (Bioline). Cycling conditions included initial denaturation for 8 min at 94 °C, followed by 35 cycles of 35 s at 94 °C, 35 s at 56 °C and 1 min at 72 °C, with a final extension step of 7 min at 72 °C. Amplification of each sample was confirmed by loading 5 μL of PCR reaction onto 2% agarose gel stained with Safe-Red™ dye and visualized under UV light. PCR products were purified with DNA Clean & Concentrator™ (Zymo Research) according to the manufacturer’s instructions, quantified as described above for RNA concentration measurement, and used for direct DNA sequencing (Sanger sequencing) at the STABVIDA sequencing facilities (Caparica, Portugal) using respective forward and reverse primers. Each cDNA sample was sequenced in duplicate. Using Geneious 7.0.6 software, forward and reverse sequences from each sample were aligned to generate consensus sequences [42].
3. Results
3.1. Virus Inoculation Experiment
Melon and pumpkin plants inoculated with non-viruliferous whiteflies were ToLCNDV negative when they were tested by TaqMan PCR. However, those inoculated with viruliferous whiteflies showed viral amounts at 6 dpi, 10 dpi and 15 dpi that were increasing with time. The viral loads detected in resistant melon accessions PI414723 and PI124112 were consistently lower than those found in the susceptible Piel de Sapo genotype, especially at 10 dpi and 15 dpi. In addition, pumpkin plants from accessions PI 604506 and PI 381814 had uniformly lower viral loads from 10 dpi than the susceptible C. moschata accession PI 419083 (Figure S1).
3.2. Expression Analysis of Candidate Genes in C. melo and C. moschata
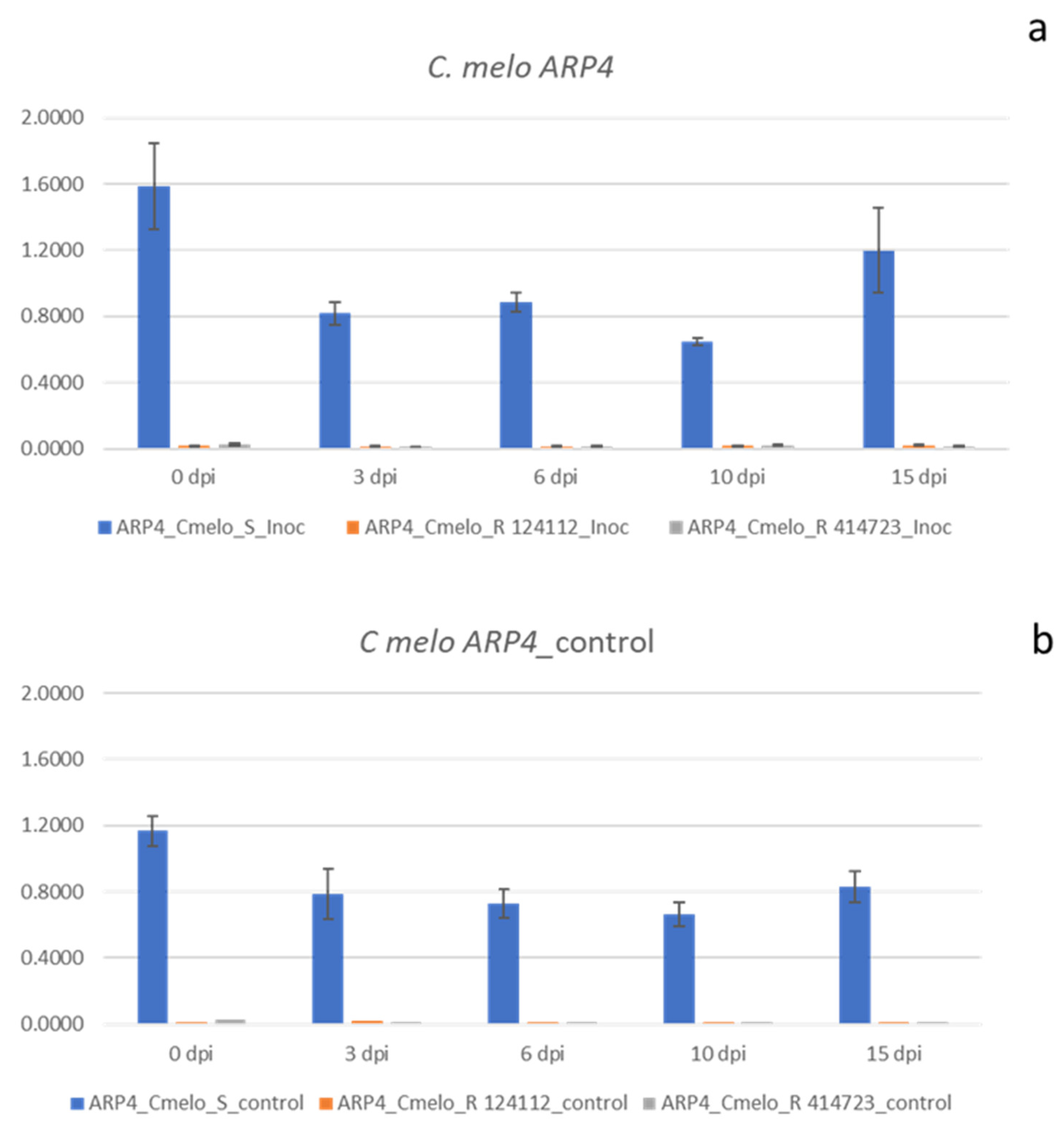
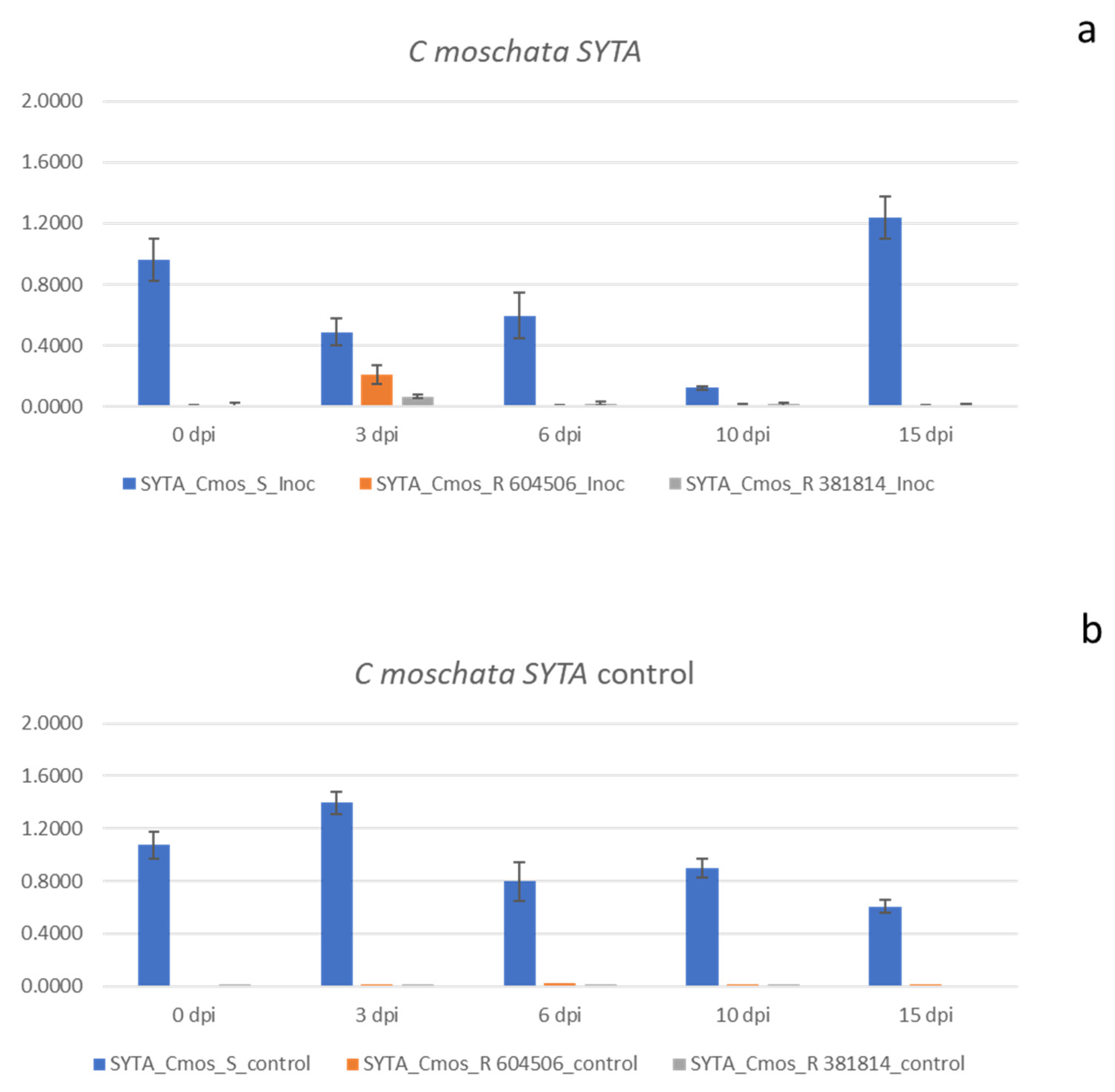
The expression profile of 10 candidate genes was assessed at different dates after ToLCNDV inoculation (0 dpi, 3 dpi, 6 dpi, 10 dpi, and 15 dpi) in three C. melo and three C. moschata contrasting genotypes for resistance. The gene set comprised host factors favoring the virus (proviral) or activating defenses against it (antiviral). Among the genes showing proviral functions, we tested ARP4, TORT, SYTA, and PELOTA. Regarding antiviral factors, six genes comprising SGS, AGO, RDR, and NIK functions were analyzed. Student’s t test showed associated probabilities below 0.05 at all time points considered for ARP4 and SYTA genes in C. melo and C. moschata, respectively (Figure 1 and Figure 2).
The most significant differences between susceptible and resistant accessions were those related to genes involved in proviral functions, such as the ARP4 gene in C. melo and the SYTA gene in C. moschata. In all of the time courses of the experiment (Figure 1), the gene Cm ARP4 was highly upregulated in the susceptible melon inoculated genotype when compared to the resistant inoculated PI 124112 and PI 414723 accessions. The Piel de Sapo cv. showed more than 40-fold higher expression when compared to the accession PI 124112 (ranging from a 42-fold change at 10 dpi to a 94-fold change at 0 dpi). In the case of accession PI 414723, the pattern was similar, with an overexpression higher than 30-fold (ranging from a 34-fold change at 10 dpi to an 83-fold change at 15 dpi). In the case of the inoculated C. moschata accessions, the most interesting results were those obtained for SYTA. In this case, the gene Cmos SYTA in the susceptible C. moschata cultivar was also upregulated when compared to the resistant accession PI 604506 and PI 381814 at all time points evaluated (Figure 2).
Apart from ARP4 in C. melo and SYTA in C. moschata, no significant expression differences were found for the rest of the analyzed genes (Figures S2 and S3)
The range of fold change was wider in the case of C. moschata when compared with C. melo. In the case of accession PI 604506, the fold change ranged from 2.35 to 92 (at 3 dpi and 15 dpi, respectively). In the case of accession PI 381814, it changed from 7 to 142 fold change (at 3 dpi and 15 dpi, respectively). These results were similar to those obtained with healthy control plants, with an overexpressed response in both susceptible accessions of C. melo (Figure 1) and C. moschata (Figure 2) when compared to the resistant genotypes. The counterparts of these genes in the other species, which are ARP4 in C. moschata and SYTA in C. melo, did not show relevant results. ARP4 in C. moschata resulted in quite similar patterns among the three accessions analyzed, whereas SYTA in C. melo did not show a differential response between the susceptible and two resistant genotypes. The gene TORTIFOLIA in C. melo showed a fold change between susceptible and resistant accessions 0.6 to 1.8. In the case of C. moschata, the range values of the fold change oscillated from 0.6 to 1.2. The fold-change range of PELOTA factor between susceptible and resistant accessions remained low, ranging from 0.2 to 1.6. Although the susceptible C. moschata genotype seemed to express a slightly higher amount of transcripts at one of the moments in time considered (10 dpi), this value was of 3.1-fold change in only one of the resistant genotypes (PI 604506), with the other sample times showing fold change values from 0.7 to 2.3.
In the case of genes involved in antiviral defense, the results were not so significant. In this regard, the fold changes observed between susceptible and resistant accessions of both species oscillated within narrower ranges. Nevertheless, three genes showed significant overexpression in at least one resistant genotype when compared to the susceptible one in both species. The genes SGS2, AGO1, and RDR1 showed fold changes higher than 2 units in both species. The resistant C. melo accession with higher contrasting response for these genes was PI 124112, showing an overexpression range of 2 to 4.4 fold change for SGS2, 0.9 to 3.07 fold change for RDR1, and 5.3 fold change at 10 dpi for AGO1 gene. In the case of C. moschata, the most extreme response was shown by the accession PI 381814, with an overexpression ranging from 0.8 to 3.2 fold change for SGS2 and 0.6 to 3.99 for AGO1. In the case of RDR1, both resistant accessions, PI 604506 and PI 381814, showed evident overexpression with ranges of 0.4–5.3 and 1.4 to 10, respectively. Although some implications for resistance control could be inferred from the results of antiviral factors, we decided to sequence only those genes with an unequivocal response in transcript amount quantification in both resistant accessions. Therefore, cDNA of ARP4 gene in C. melo as well as SYTA gene in C. moschata, both belonging to the proviral factors category, were sequenced for the three available accessions in each species.
3.3. cDNA Sequencing of Relevant Genes
Once significant and contrasting results were determined for ARP4 and SYTA gene expression in C. melo and C. moschata, respectively, the coding regions were sequenced in the three accessions of each species. Sequences from ARP4 cDNA of PI 414723, PI 124112, and Piel de Sapo (C. melo) and from SYTA cDNA of PI 604506, PI 381814, and PI 419083 were obtained. The ARP4 gene in C. melo and SYTA gene in C. moschata comprised 21 and 12 exons, respectively.
The lengths of the cDNA fragments amplified in C. melo and C. moschata for the ARP4 and SYTA genes were 1939 and 2024 bp, respectively. In the case of C. melo, the nucleotide sequence variation between the susceptible accession and the two resistant ones consisted of differences in 10 nucleotide bases. From these bases, three were common among resistant accessions and different from the susceptible one, and seven were similar in Piel de Sapo and PI 414723 and differed from the resistant accession PI 124112. The amino acid sequences, however, remained the same among the three accessions for the ARP4 protein, with nucleotide changes leading to synonymous substitutions.
A comparison of the nucleotide sequence among the three C. moschata accessions resulted in 32 SNPs. From this variation in the nucleotide sequence, only three amino acid changes were found among the three inferred protein sequences. Positions 75,339, and 483 showed these differences when comparing resistant (same sequence) and susceptible (different sequence) accessions. The conserved domains of the protein, however, remained identical in the three accessions, with the synaptotagmin-like mitochondrial-lipid-binding protein (SMP) and the C2 domain (https://www.ncbi.nlm.nih.gov/Structure/cdd) being found in all cases.
4. Discussion
In this study, a transcriptional characterization of 10 candidate genes was carried out with the aim of correlating ToLCNDV resistance with transcript amounts changing along different temporal stages. Some of those candidates were host proviral factors that the virus needs to move and replicate inside the plant, and some of them were active antiviral factors related to pathogen silencing and recognition. The molecular characterization of ToLCNDV resistance associated with proviral factors is of utmost importance in species such as C. pepo, where no resistance has been found so far. Taking advantage of an available TILLING platform in the species [28], the knock-out of the gene’s capacity to produce certain proteins needed by the virus and/or the downregulation of transcript production at this target could render a susceptible line resistant. In the case of zucchini, the identification of genetic factors involved in ToLCNDV resistance in melon and pumpkin is, therefore, not only interesting for direct introgression from C. moschata but also to screen for resistant lines in a wide mutation platform.
The most relevant results regarding gene expression have been those related to two proviral factor genes, ARP4 and SYTA in C. melo and C. moschata, respectively, where the quantity of transcripts has been nearly absent in the case of the resistant accessions when compared to the susceptible ones. In both species, similar results have been found in the inoculated and control plants, suggesting a constitutive mechanism of resistance instead of an induced one.
Plant cells’ rigid walls obstruct virus passage in and out of cells, forcing virus movement through plasmodesmata, channels penetrated by a compressed endoplasmic reticulum membrane and to neighboring cells [43]. The association between viral movement proteins and different host proteins such as ACTIN or SYTA has been reported as crucial for the movement inside the plant of different plant viruses [44]. The implication of both proteins has already been observed in ToLCNDV and other plant virus systems. In this sense, an upregulation of the ARP4 gene in an extreme ToLCNDV susceptible tomato cultivar has been reported [32], and in the case of the single-stranded DNA tomato yellow leaf curl virus (TYLCV), the requirement of the cytoskeleton for the development of viral protein aggregates has also been described [13]. Concerning SYTA, the begomovirus Cabbage leaf curl virus (CaLCuV) infection was delayed, and the cell-to-cell virus movement protein was inhibited in a knockdown line of Arabidopsis [33]. The explanation for these facts is that the dominant-negative SYTA mutant causes depletion of plasma membrane-derived endosomes, produces large intracellular vesicles attached to plasma membrane, and inhibits cell-to-cell trafficking of the virus movement proteins. In this regard, the nearly complete absence of transcripts for these two genes in the two resistant accessions of both species could indicate that the virus has difficulty moving and/or replicating within the host cell.
To discard differences in the coding cDNA region that could be promoting divergence at the protein level, we sequenced the ARP4 and SYTA cDNA in each gene for the three accessions of C. melo and C. moschata, respectively. We found some nucleotide changes, but the inferred amino acid sequences of the translated proteins remained the same in the three C. melo accessions. Regarding C. moschata, some nucleotide changes among accessions were responsible for only three amino acid variations in the protein sequence. The conserved domains, however, remained identical in the proteins of the three analyzed genotypes. Given the functional similarity of the proteins encoded by the ARP4 and SYTA genes between resistant and susceptible accessions in each species, the significant differences in transcript amounts may indicate their molecular role in resistance. These differences in the expression pattern among resistant and susceptible genotypes should be determined by a regulatory mechanism at the level of transcription or post-transcription. Although transcriptional regulation at the level of promoter should be plausible, none of these genes were located on a chromosome where the main ToLCNDV resistance QTLs have been described so far, chromosome 11 of C. melo [8] or chromosome 8 of C. moschata [27]. The ARP4 gen in C. melo is located on chromosome 2 (MELO3C017295.2, Actin related protein 4, Chr02 24576147 .. 24584360), whereas the SYTA gene is located on chromosome 19 of C. moschata (CmoCh19G007430.1 Calcium lipid binding protein, Chr19: 7626721 .. 7634112). If ARP4 or SYTA genes were involved in resistance and the gene promoters were responsible for such expression differences, the genes should be localized together with the ToLCNDV resistance QTLs already reported for the species. This could be plausible for the ARP4 gene, located in the same interval of chromosome 2 of C. melo [8] where the resistance QTL has been described (21.840.921–24.977.990) [14]. Transcriptional regulation through a transcription factor or post-transcriptional regulation can, however, regulate the expression of a gene located in a distant genomic region or even on a different chromosome. Transcriptional gene regulation can decrease RNA synthesis, while post-transcriptional gene silencing can degrade mRNA. According to [45], the role of miRNAs in growth, development, and stress responses cannot be underestimated. This class of tiny non-coding RNAs, which are considered to be important regulators at the post-transcriptional level, regulate the expression of their target genes either by cleaving the target transcripts or by translational inhibition.
In this context, a recent study reported the implication of the module constituted by miRNA 166 (sly-miR166) and its target, a homeobox leucine zipper transcription factor (SlyHB) in ToLCNDV resistance in tomato [46]. This miRNA sly-miR166 is differentially regulated upon ToLCNDV infection in two tomato cultivars contrasting for resistance, being highly expressed in the resistant accession and minimally expressed in the susceptible one, thus leading to the opposite situation in the Homeobox Leucine (HL) transcription factor that showed low and high expression in the resistant and susceptible genotypes, respectively. While the BLAST search of the tomato SlyHB sequence on the C. melo genome showed three homologue sequences in chromosomes where no ToLCNDV QTLs were described, in the case of C. moschata, the homologue fragment CmoCh08G006920.1 (annotated as DNA binding protein) was located in chromosome 8 (4405129 .. 4410067) inside the limits of the ToLCNDV resistance QTL reported by [27]. In plants, miRNAs act on specific target mRNAs in a complete or near-perfect base-pairing manner, resulting in mRNA degradation or translation inhibition [46]. In the case of C. moschata, miR166 is located in exons 4 and 5 of the HL transcription factor gene, pointing to a post-transcriptional regulation event where once the gene is transcribed, it could be cleaved by the miRNA.
The fact that both resistant accessions showed an identical expression pattern with a nearly nonexistent amount of transcripts for the two genes considered, ARP4 in the case of C. melo and SYTA in the case of C. moschata, led us to assume that the genomic region in charge of the regulation of these genes should be relevant for resistance. Future studies to deepen the knowledge of QTLs as well as to decipher the gene regulation process will shed light on the molecular dialogue that sustains ToLCNDV resistance. A temporal stage microscopy phenotyping and/or a phenotypic quantification of candidate gene transcripts when identifying QTL involved, apart from the traditional gross evaluation of symptoms, could help to elucidate the complex resistance mechanism. Moreover, an integrative study of miRNA regulators and their target mRNA transcripts in the first stages of the disease process could allow us to identify the factors involved in the complex gene crosstalk of the regulatory mechanisms of ToCNDV resistance in cucurbits.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9020231/s1, Figure S1: ToLCNDV-ES accumulation in C. melo (a) and C. moschata (b) accessions resistant (R) and susceptible (S) at 0, 3, 6 and 15 days after inoculation (dpi) with viruliferous whiteflies. Values are given as Log 2−ΔΔCt of leaf pooled from four plants. Figure S2: Expression profile of TORTIFOLIA (a), PELOTA (b), SGS1 (c), SGS2 (d), AGO (e), RDR1 (f), RDR6 (g) NIK1 (h) and SYTA (i) genes in C. melo accessions ToLCNDV inoculated at 0, 3, 6, 10 and 15 dpi. Figure S3: Expression profile of TORTIFOLIA (a), PELOTA (b), SGS1 (c), SGS2 (d), AGO (e), RDR1 (f), RDR6 (g) NIK1 (h) and ARP4 (i) genes in C. moschata accessions ToLCNDV inoculated at 0, 3, 6, 10 and 15 dpi.
Author Contributions
Conceptualization, B.R., P.G. and L.R.; methodology, P.G., D.J., L.R. and B.R.; formal analysis, B.R. and L.R.; investigation, B.R., P.G. and L.R.; resources, P.G.; data curation, D.J. and B.R.; writing—original draft preparation, B.R. and L.R.; writing—review and editing, B.R., D.J., P.G. and L.R.; funding acquisition, P.G., L.R. and D.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by project PID2021-125787OR-C32 from Spanish Ministerio de Ciencia e Innovación co-financed by FEDER.
Acknowledgments
B.R. acknowledges the useful contribution of C.I. González-Verdejo to improve the manuscript in regard to gene regulation discussion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jones, R.A.C.; Naidu, A.A. Global Dimensions of Plant Virus Diseases: Current Status and Future Perspectives. Annu. Rev. Virol. 2019, 6, 387–409. [Google Scholar] [CrossRef]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First detection of Tomato leaf curl New Delhi virus infecting zucchini in Spain. Plant Dis. 2014, 98, 857. [Google Scholar] [CrossRef]
- Ruiz, M.L.; Simón, A.; Velasco, L.; García, M.C.; Janssen, D. First report of Tomato leaf curl New Delhi virus infecting tomato in Spain. Plant Dis. 2015, 98, 894. [Google Scholar] [CrossRef]
- Ruiz, L.; Simon, A.; Velasco, L.; Janssen, D. Biological characterization of Tomato leaf curl New Delhi virus from Spain. Plant Pathol. 2017, 66, 376–382. [Google Scholar] [CrossRef]
- Janssen, D.; Simón, A.; Boulares, M.; Ruiz, L. Host Species-Dependent Transmission of Tomato Leaf Curl New Delhi Virus-ES by Bemisia tabaci. Plants 2022, 11, 390. [Google Scholar] [CrossRef]
- López, C.; Ferriol, M.; Picó, M.B. Mechanical transmission of Tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Sáez, C.; Esteras, C.; Martínez, C.; Ferriol, M.; Narinder, P.S.D.; López, C.; Picó, B. Resistance to tomato leaf curl New Delhi virus in melon is controlled by a major QTL located in chromosome 11. Plant Cell Rep. 2017, 36, 1571–1584. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; López, C.; Picó, B. Resistance to tomato leaf curl New Delhi virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Juárez, M.; Rabadan, M.P.; Martinez, L.D.; Tayahi, M.; Grande-Perez, A.; Gomez, P. Natural hosts and genetic diversity of the emerging tomato leaf curl New Delhi virus in Spain. Front. Microbiol. 2019, 10, 140. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Susceptibility Genes to Plant Viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef]
- Moshe, A.; Belausov, E.; Niehl, A.; Heinlein, M.; Czosnek, H.; Gorovits, R. The Tomato yellow leaf curl virus V2 protein forms aggregates depending on the cytoskeleton integrity and binds viral genomic DNA. Sci. Rep. 2015, 5, 9967. [Google Scholar] [CrossRef]
- Roman B, Gómez P, Picó B, López C, Janssen D (2019) Candidate gene analysis of Tomato leaf curl New Delhi virus resistance in Cucumis melo. Sci. Hortic. 2019, 243, 12–20. [CrossRef]
- Steinberg, G. Tracks for traffic: Microtubules in the plant pathogen Ustilago maydis. New Phytol. 2007, 174, 721–733. [Google Scholar] [CrossRef]
- Pena, E.J.; Ferriol, I.; Sambade, A.; Buschmann, H.; Niehl, A.; Elena, S.F.; Rubio, L.; Heinlein, M. Experimental Virus Evolution Reveals a Role of Plant Microtubule Dynamics and TORTIFOLIA1/SPIRAL2 in RNA Trafficking. PLoS ONE 2014, 9, e105364. [Google Scholar] [CrossRef]
- Uchiyama, A.; Shimada-Beltran, H.; Levy, A.; Zheng, J.Y.; Javia, P.A.; Lazarowitz, S.G. The Arabidopsis synaptotagmin SYTA regulates the cell-to-cell movement of diverse plant viruses. Front. Plant Sci. 2014, 5, 584. [Google Scholar] [CrossRef]
- Robaglia, C.; Caranta, C. Translation initiation factors: A weak link in plant RNA virus infection. Trends Plant Sci. 2006, 11, 40–45. [Google Scholar] [CrossRef]
- Lapidot, M.; Karniel, U.; Gelbart, D.; Fogel, D.; Evenor, D.; Kutsher, Y.; Makhbash, Z.; Nahon, S.; Shlomo, H.; Chen, L.; et al. A Novel Route Controlling Begomovirus Resistance by the Messenger RNA Surveillance Factor Pelota. PLoS Genet. 2015, 11, e1005538A. [Google Scholar] [CrossRef]
- Koeda, S.; Onouchi, M.; Mori, N.; Syafira, P.N.; Nagano, A.J.; Kesumawati, E. A recessive gene pepy-1 encoding Pelota confers resistance to begomovirus isolates of PepYLCIV and PepYLCAV in Capsicum annuum. Theor. Appl. Genet. 2021, 134, 2947–2964. [Google Scholar] [CrossRef]
- Duan, C.G.; Wang, C.H.; Guo, H.S. Application of RNA silencing to plant disease resistance. Silence 2015, 3, 5. [Google Scholar]
- Bai, M.; Yang, G.S.; Chen, W.T.; Mao, Z.C.; Kang, H.X.; Chen, G.H.; Yang, Y.H.; Xie, B.Y. Genome-wide identification of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analyses in response to viral infection and abiotic stresses in Solanum lycopersicum. Gene 2012, 501, 52–62. [Google Scholar] [CrossRef]
- Li, F.; Huang, C.; Li, Z.; Zhou, X. Suppression of RNA Silencing by a Plant DNA Virus Satellite Requires a Host Calmodulin-Like Protein to Repress RDR6 Expression. PLoS Pathog. 2014, 10, e1003921. [Google Scholar]
- Kushwaha, N.; Singh, A.; Basu, S.; Chakraborty, S. Differential response of diverse solanaceous hosts to tomato leaf curl New Delhi virus infection indicates coordinated action of NBS-LRR and RNAi-mediated host defense. Arch. Virol. 2015, 160, 1499–1509. [Google Scholar] [CrossRef]
- Hussain, M.; Mansoor, S.; Iram, S.; Fatima, A.N.; Zafar, Y. The nuclear shuttle protein of Tomato leaf curl New Delhi virus is a pathogenicity determinant. J. Virol. 2005, 79, 4434–4439. [Google Scholar] [CrossRef]
- Machado, J.P.B.; Calil, I.P.; Santos, A.A.; Fontes, E.P.B. Translational control in plant antiviral immunity. Genet. Mol. Biol. 2017, 40, 292–304. [Google Scholar]
- Sáez, C.; Martínez, C.; Montero-Pau, J.; Esteras, C.; Blanca, J.; Sifres, A.; Ferriol, M.; López, C.; Picó, B. A major QTL located in chromosome 8 of Cucurbita moschata is responsible for resistance to tomato leaf curl New Delhi virus (ToLCNDV). Front. Plant Sci. 2020, 11, 207. [Google Scholar]
- Vicente-Dólera, N.; Troadec, C.; Moya, M.; del Río-Celestino, M.; Pomares-Viciana, T.; Bendahmane, A.; Picó, B.; Román, B.; Gómez, P. First TILLING Platform in Cucurbita pepo: A New Mutant Resource for Gene Function and Crop Improvement. PLoS ONE 2014, 9, e112743. [Google Scholar]
- Janssen, D.; Simon, A.; Crespo, O.; Ruiz, L. Genetic population structure of Bemisia tabaci in Spain associated with Tomato leaf curl New Delhi virus. Plant Protect. Sci. 2017, 53, 25–31. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta Ct) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Su, S.; Liu, Z.; Chen, C.; Zhang, Y.; Wang, X.; Zhu, L.; Miao, L.; Wang, X.C.; Yuan, M. Cucumber mosaic virus movement protein severs actin filaments to increase the plasmodesmal size exclusion limit in tobacco. Plant Cell 2010, 22, 1373–1387. [Google Scholar] [CrossRef]
- Naqvi, A.R.; Sarwat, M.; Pradhan, B.; Choudhury, N.R.; Haq, Q.M.; Mukherjee, S.K. Differential expression analyses of host genes involved in systemic infection of Tomato leaf curl New Delhi virus (ToLCNDV). Virus Res. 2011, 160, 395–399. [Google Scholar]
- Lewis, J.D.; Lazarowitz, S. Arabidopsis synaptotagmin SYTA regulates endocytosis and virus movement protein cell-to-cell transport. Proc. Natl. Acad. Sci. USA 2010, 107, 2491–2496. [Google Scholar]
- Levy, A.; Zheng, J.Y.; Lazarowitz, S.C. Synaptotagmin SYTA forms ER-Plasma membrane Junctions that are recruited to plasmodesmata for plant virus movement. Curr. Biol. 2015, 25, 2018–2025. [Google Scholar] [CrossRef]
- Kumar, V.; Naqvi, A.R. Tomato leaf curl New Delhi virus (ToLCNDV) encoded AC2 associates with host mirnas by directly interacting with AGO1. J. RNAi Gene Silencing 2016, 12, 515–520. [Google Scholar]
- Basu, S.; Kushwaha, N.K.; Singh, A.K.; Sahu, P.P.; Kumar, R.V.; Chakraborty, S. Dynamics of a geminivirus-encoded pre-coat protein and host RNA-dependent RNA polymerase 1 in regulating symptom recovery in tobacco. J. Exp. Bot. 2018, 69, 2085–2102. [Google Scholar] [CrossRef]
- Leibman, D.; Kravchik, M.; Wolf, D.; Haviv, S.; Weissberg, M.; Ophir, R.; Paris, H.S.; Palukaitis, P.; Ding, S.W.; Gaba, V.; et al. Differential expression of cucumber RNA-dependent RNA polymerase 1 genes during antiviral defence and resistance. Mol. Plant. Pathol. 2018, 19, 300–312. [Google Scholar] [CrossRef]
- Hussain, M.; Mansoor, S.; Iram, S.; Zafar, Y.; Briddon, R.W. The Hypersensitive Response to Tomato leaf curl New Delhi virus Nuclear Shuttle Protein Is Inhibited by Transcriptional Activator Protein. Mol. Plant-Microbe Interact. 2007, 20, 1581–1588. [Google Scholar]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.; Moorman, A.F. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar]
- Kong, Q.; Yuan, J.; Niu, P.; Xie, J.; Jiang, W.; Huang, Y.; Bie, Z. Screening suitable reference genes for normalization in reverse transcription quantitative Real-Time PCR analysis in melon. PLoS ONE 2014, 9, e87197. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar]
- Ishikawa, K.; Hashimoto, M.; Yusa, A.; Koinuma, H.; Kitazawa, Y.; Netsu, O.; Yamaji, Y.; Namba, S. Dual targeting of a virus movement protein to ER and plasma membrane subdomains is essential for plasmodesmata localization. PLoS Pathog. 2017, 13, e1006463. [Google Scholar]
- Levy, A.; Tilsner, J. Creating Contacts Between Replication and Movement at Plasmodesmata–A Role for Membrane Contact Sites in Plant Virus Infections. Front. Plant Sci. 2020, 11, 862. [Google Scholar]
- Prasad, A.; Sharma, N.; Muthamilarasan, M.; Rana, S.; Prasad, M. Recent advances in small RNA mediated plant-virus interactions. Crit. Rev. Biotechnol. 2019, 39, 587–601. [Google Scholar] [CrossRef]
- Prasad, A.; Sharma, N.; Chirom, O.; Prasad, M. The sly-miR166-SlyHB module acts as a susceptibility factor during ToLCNDV infection. Theor. Appl. Genet. 2022, 135, 233–242. [Google Scholar] [CrossRef]
Figure 1.
Expression profile of ARP4 gene in C. melo accessions. C. melo plants ToLCNDV inoculated at 0, 3, 6, 10 and 15 dpi (a). Control plants of C. melo accessions (non inoculated) (b). Student’s t test associated probability in all cases p ≤ 0.05.
Figure 1.
Expression profile of ARP4 gene in C. melo accessions. C. melo plants ToLCNDV inoculated at 0, 3, 6, 10 and 15 dpi (a). Control plants of C. melo accessions (non inoculated) (b). Student’s t test associated probability in all cases p ≤ 0.05.

Figure 2.
Expression profile of SYTA gene in C. moschata accessions. C. moschata plants ToLCNDV inoculated at 0, 3, 6, 10 and 15 dpi (a). Control plants of C. moschata accessions (b). Student’s t test associated probability in all cases, p ≤ 0.05.
Figure 2.
Expression profile of SYTA gene in C. moschata accessions. C. moschata plants ToLCNDV inoculated at 0, 3, 6, 10 and 15 dpi (a). Control plants of C. moschata accessions (b). Student’s t test associated probability in all cases, p ≤ 0.05.


{kind=link}
{kind=link}
Table 1.
Primer sequences used in RT-PCR for amplifying candidate genes in C melo and C. moschata.
| Target Gene | Sequence | Protein Coding Gene | ||
|---|---|---|---|---|
| Cucumis melo | Cucurbita moschata | Cucumis melo | Cucurbita moschata | |
| Cm ARP4/ Cmos ARP4 Actin related protein | F: TCGAAGGCAATGTTGTAGCA R: TATTGCCCCCATTAATTTGC | F: GAAGAACATGGGGCTTCCTA R: GGATCAAGCGTCCAGAACAG | MELO3C017295T1_Ch02 | CmoCh10G002600 |
| Cm TORT/ Cmos TORT Tortifolia | F: GAATTGACTGCGGAAGCATT R: TGTCCATATCACCTGCCTGA | F: TGTCGAGAAACGGACAGATG R: CTTCTGTTACCTCCGCCTTG | MELO3C018075.2.1_Ch04 | CmoCh09G009400.1 |
| Cm SYTA/ Cmos SYTA Synaptotagmin | F: GACATCAAGTCAGCCCCAAC R: CCAAGCTTTCCTTGTTTTGC | F: GCCATTGGTTCCAAGCTTTC R: CCCCAACAAGTTTTAGGCCA | MELO3C013361.2.1_Ch01 | CmoCh19G007430.1 |
| Cm PELOTA/ Cmos PELOTA RNA survell. factor Pelota | F: TGACGGCCATAACAGTGTCT R: CTGGTAGCGTCAAGATGGTG | F: AGTGTCAAGATGGTGCCACT R: TTCCTAACGGTGACAGCCAT | MELO3C008594.2.1_Ch05 | CmoCh05G008420.1 |
| Cm SGS1/ Cmos SGS1 Suppressor of gene silencing 1 | F: GGATCTGATTATGTGGCCTCC R: ATCCATTGCTTTGTTCCCCA | F: TGATTATGTGGCCTCCTCTAGTT R: TTACTATCCATTGCCTTGTTTCC | MELO3C012650_Ch01 | CmoCh08G011180 |
| Cm SGS2/ Cmos SGS2 Suppressor of gene silencing 2 | F: ACAAGTCCTCTCAGAAGCCT R: GATTTAGGGTCGGAGGGAGG | F: CCACCAGGTCGTCTCAGAAG R: AGGGAGGATTGTTGGTGGAG | MELO3C012650.2_Ch01 | CmoCh17G002200 |
| Cm AGO1/ Cmos AGO1 Argonaute | F: TCATGGCTGAATTGGTGAGA R: AAAGGAAGCTCACCAGCTGTA | MELO3C006494.2.1_Ch06 | CmoCh14G017940.1 | |
| Cm RDR1/ Cmos RDR1 RdRNA-pol | F: TGGAGCTCCTCGGATATATAAAA R: CCCATTGATCATCAGGTACTTC | MELO3C026815.2.1_Ch10 | CmoCh04G000890.1 | |
| Cm RDR6/ Cmos RDR6 RdRNA-pol | F: TGACCGCAGCAAGTATGGA R: GGTCACGATCTTCCCTGTCT | F: GCAGTGAGTATGGAGCTTTGG R: GGCATCGTCACAATCTTCCC | MELO3C011257.2.1_Ch03 | CmoCh05G003850.1 |
| Cm NIK1/ Cmos NIK1 NSP-interacting kinases | F: AAAACGCTGTGGATCCATGT R: CGACAAGCTACCGGACAAGT | F: TTGGGATGAAAACGCTGTGG R: GGCTACCCGACAAGTTCTGA | MELO3C012187_Ch10 | CmoCh18G010900 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Román, B.; Gómez, P.; Janssen, D.; Ruiz, L. Insights into the Key Genes in Cucumis melo and Cucurbita moschata ToLCNDV Resistance. Horticulturae 2023, 9, 231. https://doi.org/10.3390/horticulturae9020231
AMA Style
Román B, Gómez P, Janssen D, Ruiz L. Insights into the Key Genes in Cucumis melo and Cucurbita moschata ToLCNDV Resistance. Horticulturae. 2023; 9(2):231. https://doi.org/10.3390/horticulturae9020231
Chicago/Turabian StyleRomán, Belén, Pedro Gómez, Dirk Janssen, and Leticia Ruiz. 2023. "Insights into the Key Genes in Cucumis melo and Cucurbita moschata ToLCNDV Resistance" Horticulturae 9, no. 2: 231. https://doi.org/10.3390/horticulturae9020231
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.