



Transformation of Strawberry Plants’ Phenolic Profile after Treatment with a Mechanocomposite Based on Silicon Chelates in the Course of Development under In Vitro, Ex Vitro, and In Vivo Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.1.1. Micropropagation
2.1.2. In Vivo Conditions
2.1.3. Description of the MC, Treatments, and Experimental Design
- In vitro, MC was added to the B5 medium at a concentration of 5.0 mg L−1 before autoclaving (121 °C, 1.2 atm for 20 min) (+MC). The B5 medium without the MC served as a control (−MC). Plant microshoots were collected for analysis at the end of each cultivation cycle (in vitro multiplication and in vitro rooting after 4 and 8 weeks, respectively);
- At the ex vitro acclimation stage, the plants were moistened with the MC slurry at a concentration of 0.3 g L−1 dissolved in a ¼-strength Murashige and Skoog’s (MS) [40] salt solution (27 mL per cell) once a period (4 weeks) (+MC). A control group of plants was moistened with a ¼-strength MS salt solution (−MC). Shoots were sampled for analysis after 4 weeks of ex vitro acclimation of the microclones;
- In the greenhouse, each plant was watered under roots with an MC slurry at a concentration of 0.3 g L−1 in tap water (90 mL per pot) at 50% of full moisture capacity twice a period at regular intervals (2 weeks). Control specimens were moistened with tap water (−MC);
- In the field, root treatment of plants with 0.3 g L−1 MC dissolved in tap water (2.7 L per plant) was carried out at 2 weeks after planting (+MC). Control plants were watered with tap water (−MC). The number of treatments during the growing season was three at regular intervals (3 weeks).
2.2. High Performance Liquid Chromatography (HPLC) Analysis of Phenolic Compounds
2.3. Statistical Analysis
3. Results
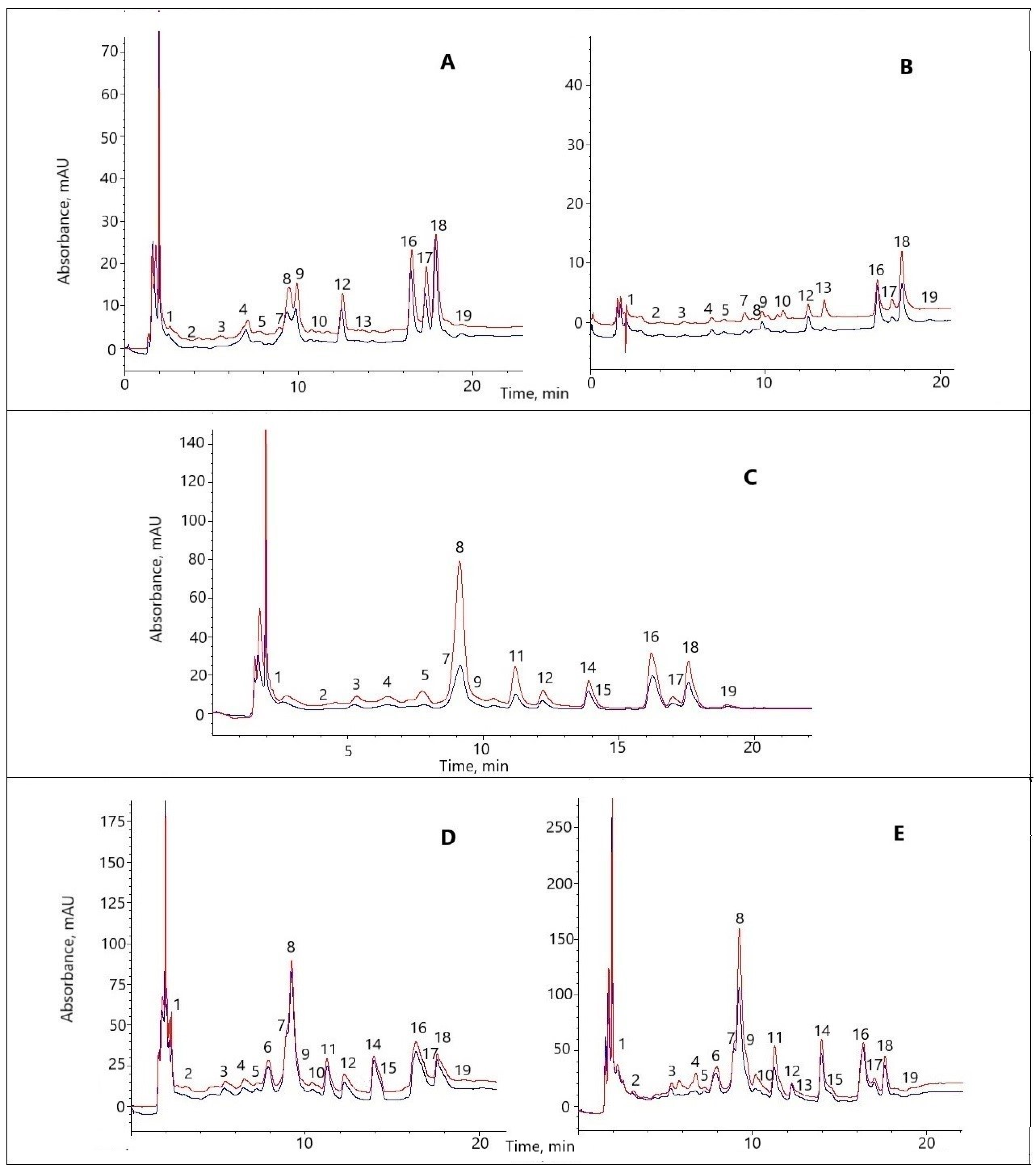
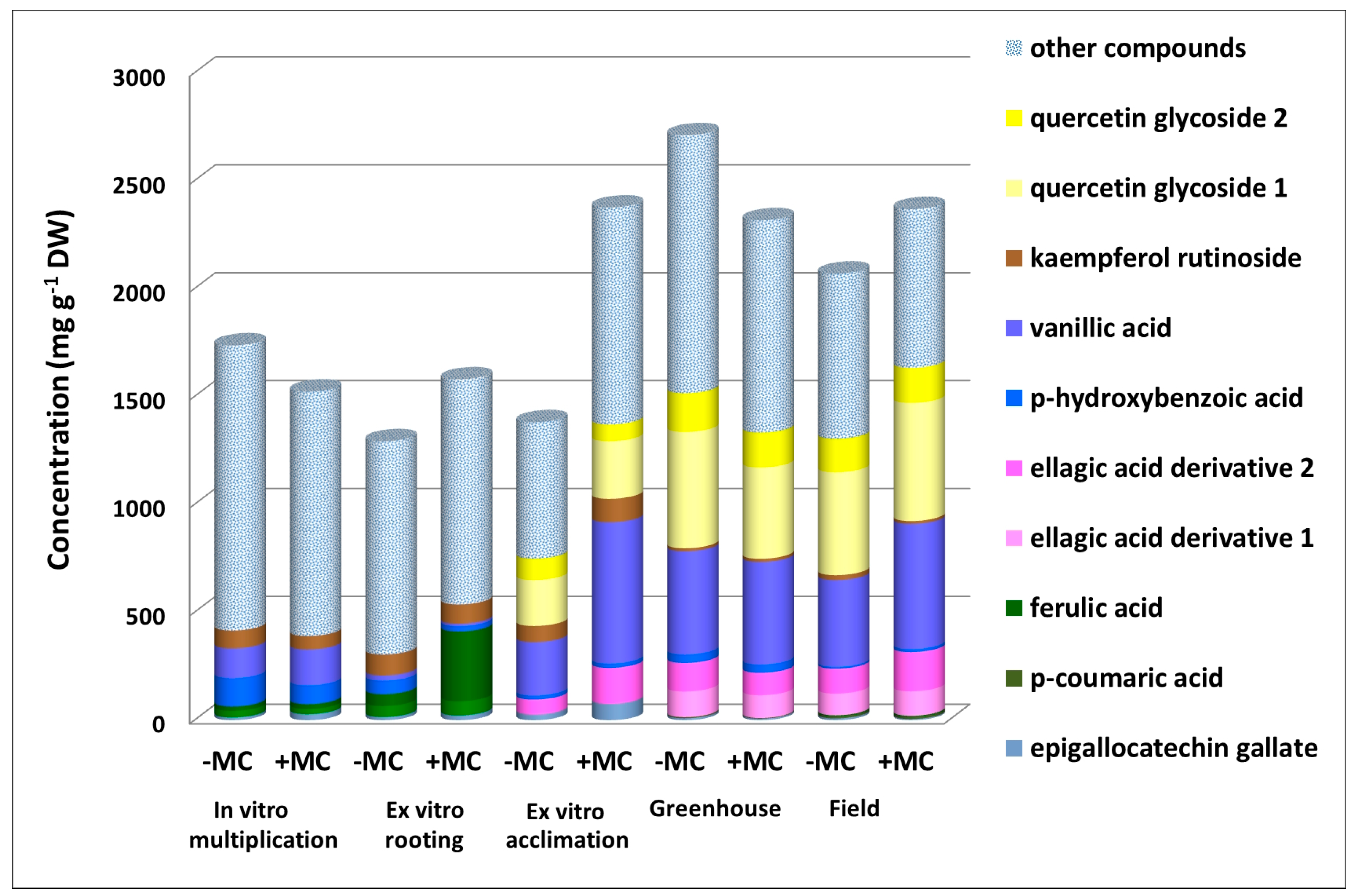
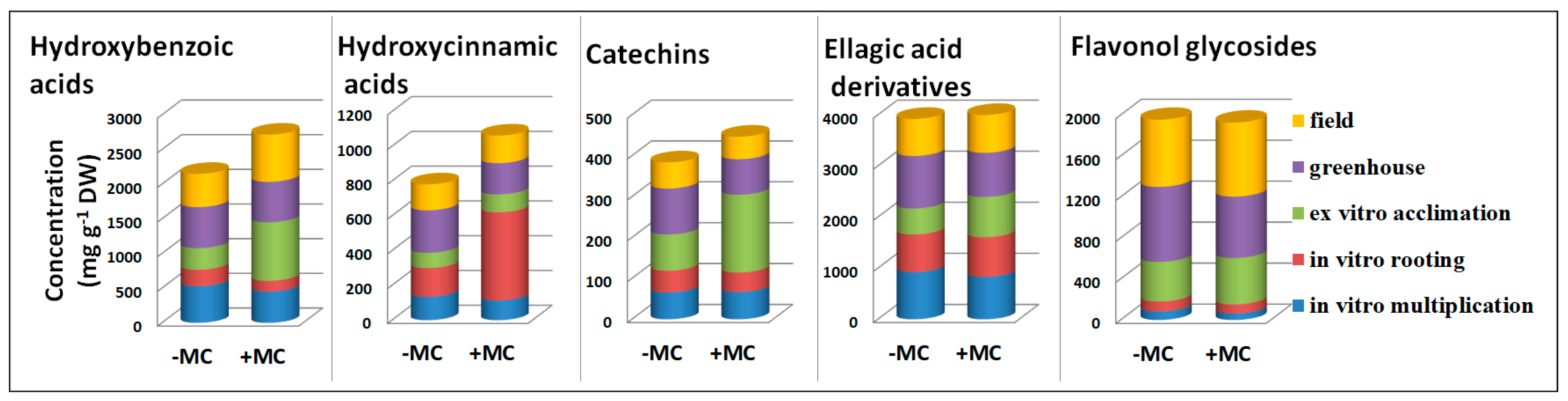
3.1. The Phenolic Profile of the Aboveground Parts of F. × ananassa
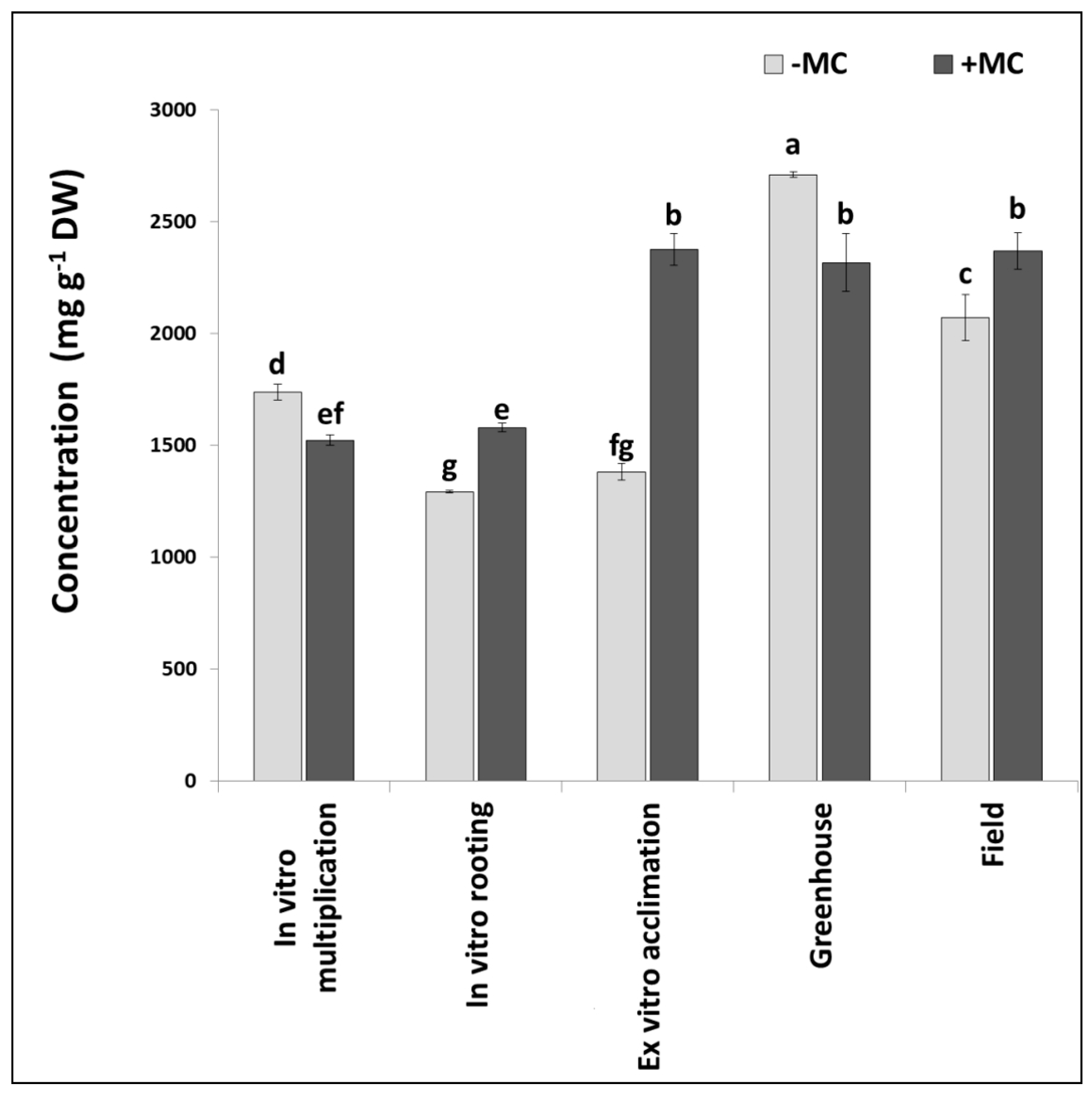
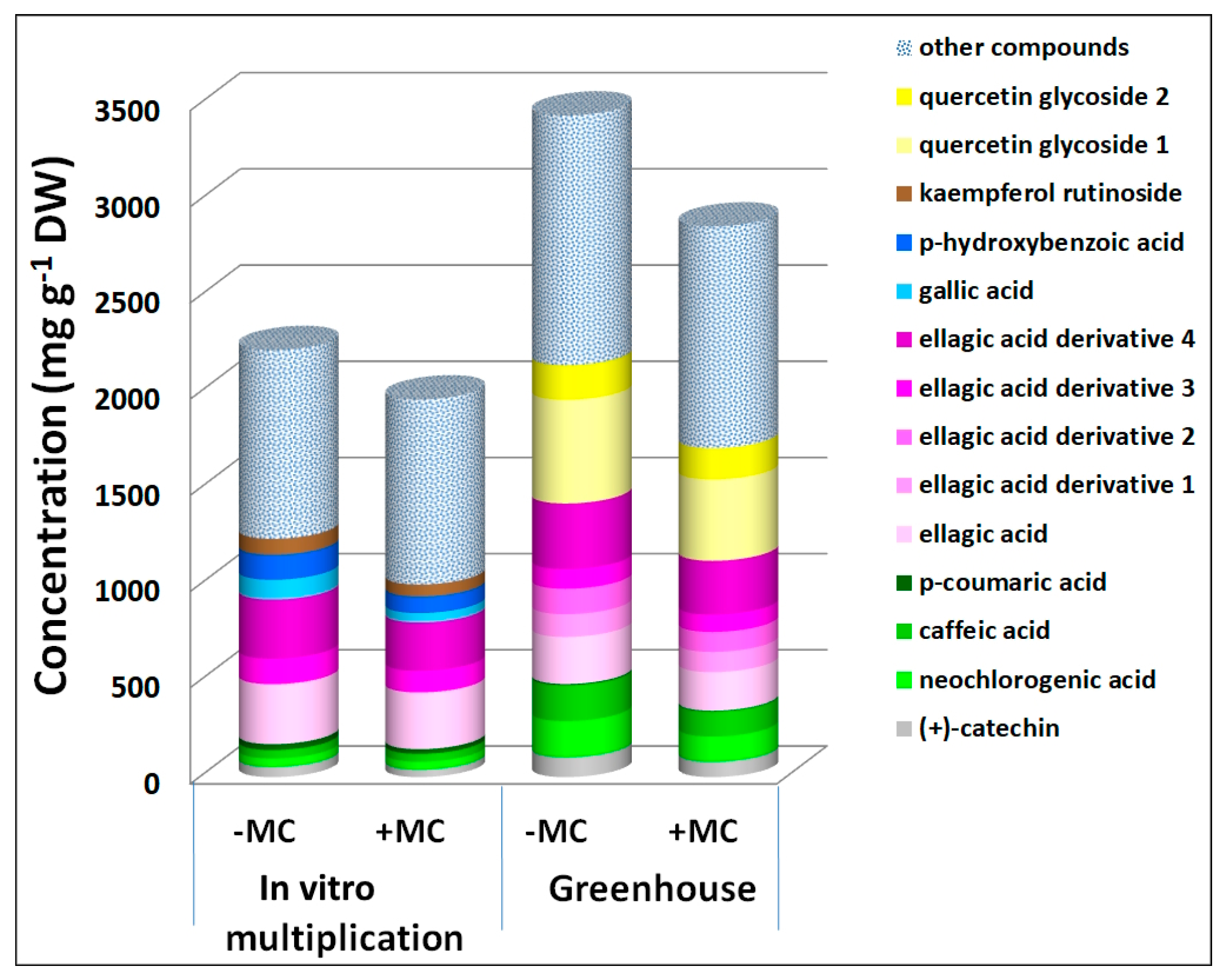
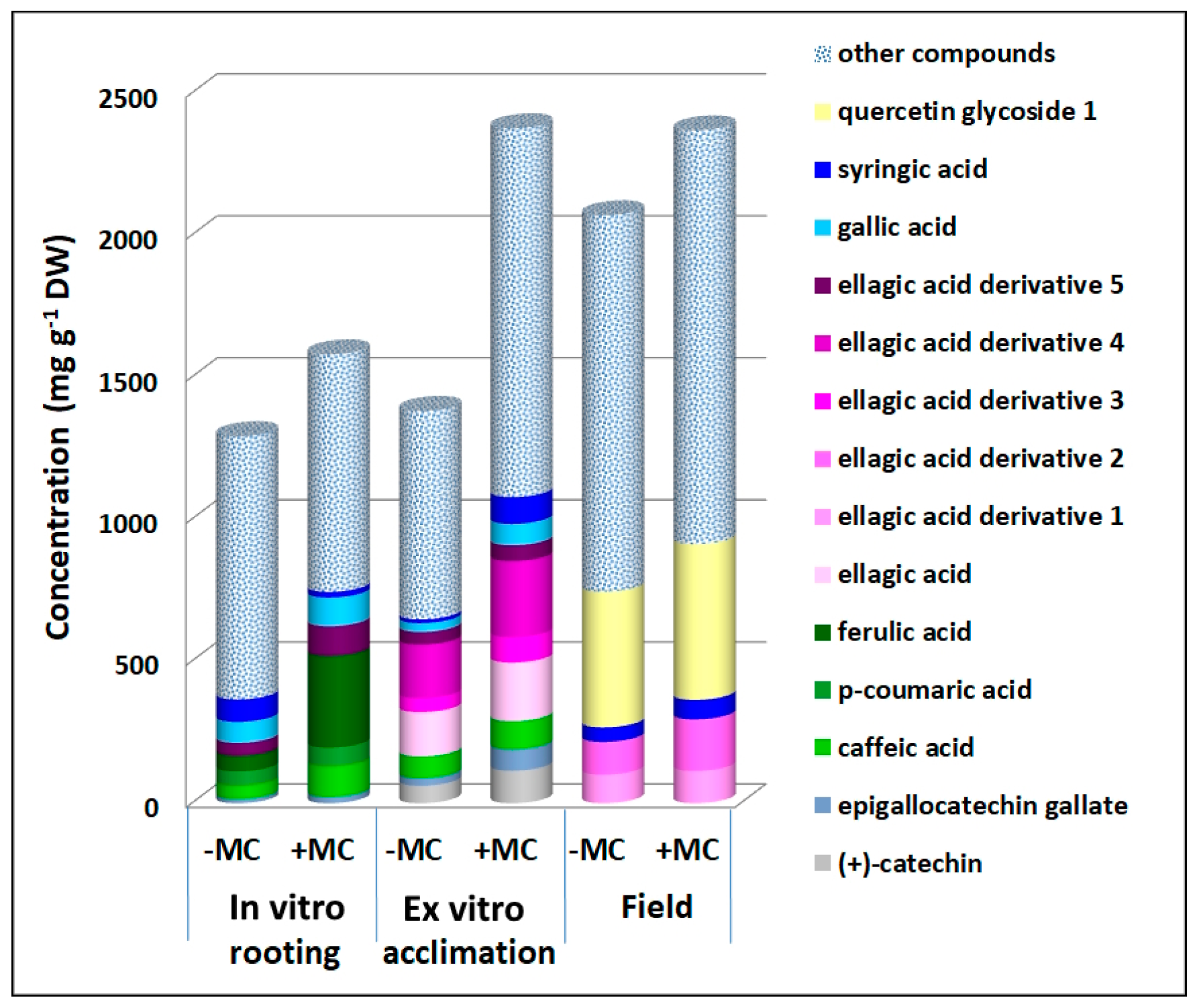
3.2. The Effect of the MC on the Phenolic Profile
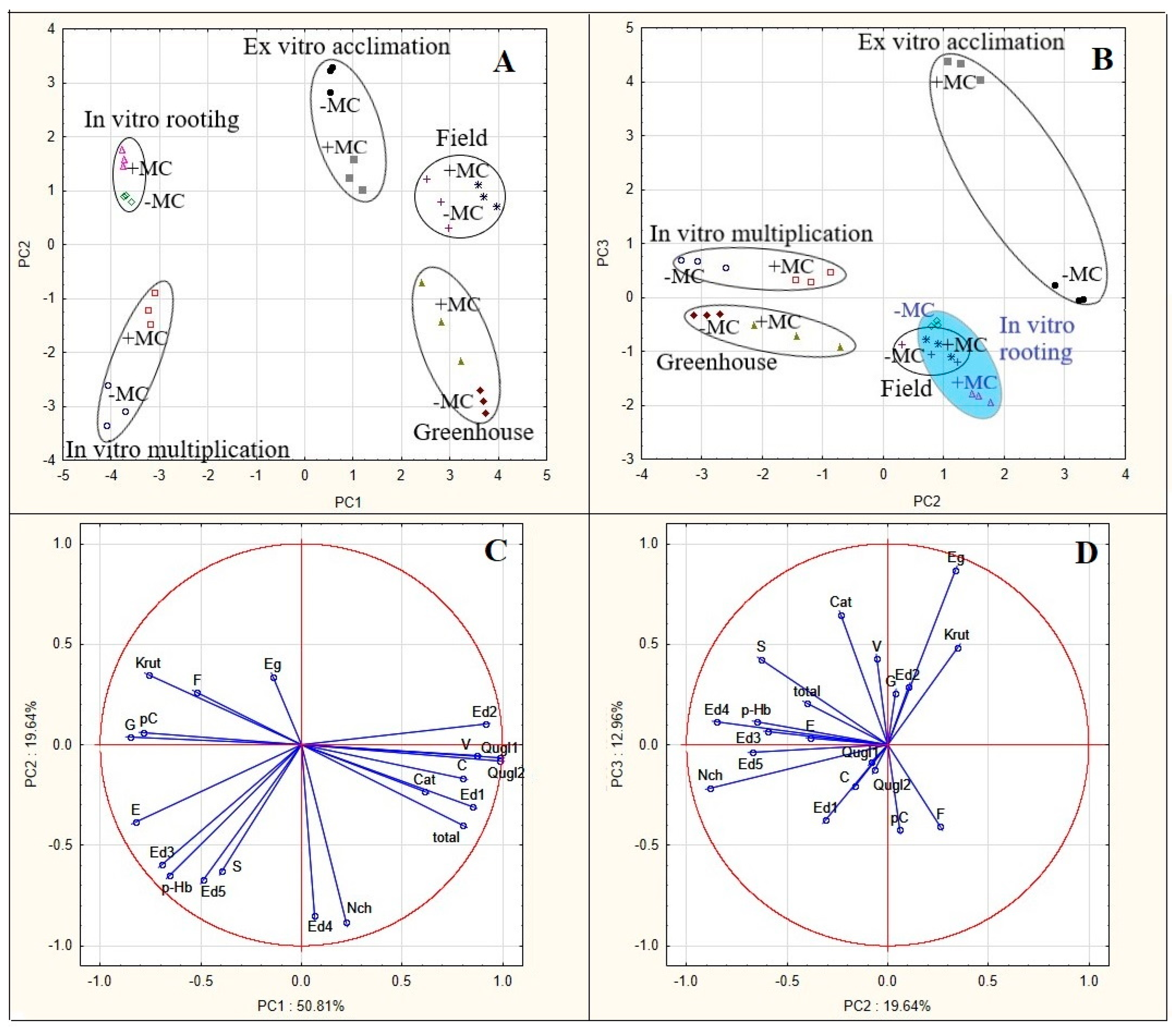
3.3. PCA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Siletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Durán-Lara, E.F.; Valderrama, A.; Marican, A. Natural organic compounds for application in organic farming. Agriculture 2020, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Lomovsky, O.I.; Lomovskiy, I.O.; Orlov, D.V. Mechanochemical solid acid/base reactions for obtaining biologically active preparations and extracting plant materials. Green Chem. Lett. Rev. 2017, 10, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Trofimova, E.G.; Podgorbunskikh, E.M.; Skripkina, T.S.; Bychkov, A.L.; Lomovsky, O.I. Scaling of the mechanochemical process of production of silicon chelates. Bulg. Chem. Commun. 2018, 50, 45–48. [Google Scholar]
- Ambros, E.; Karpova, E.; Kotsupiy, O.; Zaytseva, Y.; Trofmova, E.; Novikova, T. Silicon chelates from plant waste promote in vitro shoot production and physiological changes in strawberry plantlets. Plant Cell Tissue Organ. Cult. 2021, 145, 209–221. [Google Scholar] [CrossRef]
- Savvas, D.; Ntatsi, G. Biostimulant activity of silicon in horticulture. Sci. Hortic. 2015, 196, 66–81. [Google Scholar] [CrossRef]
- Szulc, W.; Rutkowska, B.; Hoch, M.; Ptasinski, D.; Kazberuk, W. Plant available silicon in differentiated fertilizing conditions. Plant Soil Environ. 2019, 65, 233–237. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Tian, D.-D.; Guo, D.-J.; Chen, Z.-L.; Zhong, C.-S.; Nikpay, A.; Singh, M.; Rajput, V.D.; Singh, R.K.; et al. Influence of silicon on biocontrol strategies to manage biotic stress for crop protection, performance, and improvement. Plants 2021, 10, 2163. [Google Scholar] [CrossRef]
- Guerriero, G.; Hausman, J.F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Sci. 2016, 7, 463. [Google Scholar] [CrossRef] [Green Version]
- Carneiro-Carvalho, A.; Aires, A.; Anjos, R.; Martins, L.; Pinto, T.; Peixoto, F.; Gomes-Laranjo, J. The role of silicon fertilization in the synthesis of phenolic compounds on chestnut plants infected with P. cinnamomi and C. parasitica. J. Plant Dis. Prot. 2020, 127, 211–227. [Google Scholar] [CrossRef]
- Vega, I.; Rumpel, C.; Ruíz, A.; Mora, M.d.l.L.; Calderini, D.F.; Cartes, P. Silicon modulates the production and composition of phenols in barley under aluminum stress. Agronomy 2020, 10, 1138. [Google Scholar] [CrossRef]
- Araújo, W.B.S.; Teixeira, G.C.M.; de Mello Prado, R.; Rocha, A.M.S. Silicon mitigates nutritional stress of nitrogen, phosphorus, and calcium deficiency in two forages plants. Sci. Rep. 2022, 12, 6611. [Google Scholar] [CrossRef]
- Kulbat, K. The role of phenolic compounds in plant resistance. Biotechnol. Food Sci. 2016, 80, 97–108. [Google Scholar]
- Šamec, D.; Karalija, E.; Šola, I.; Vujcic Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Matkowski, A. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv. 2008, 26, 548–560. [Google Scholar] [CrossRef]
- Kårlund, A.; Salminen, J.-P.; Koskinen, P.; Ahern, J.R.; Karonen, M.; Tiilikkala, K.; Karjalainen, R.O. Polyphenols in strawberry (Fragaria × ananassa) leaves induced by plant activators. J. Agric. Food Chem. 2014, 62, 4592–4600. [Google Scholar] [CrossRef]
- Dongju, L.; Qing, M.; Yiwen, Z.; Zeyan, P. Phenolic compounds with antioxidant activity from strawberry leaves: A study on microwave-assisted extraction optimization. Prep. Biochem. Biotechnol. 2020, 50, 874–882. [Google Scholar] [CrossRef]
- Kotsupiy, O.; Karpova, E.; Ambros, E. Phenolic compounds in strawberry (Fragaria × ananassa Duch.) microshoots. BIO Web Conf. 2020, 24, 00041. [Google Scholar] [CrossRef]
- Ambros, E.; Karpova, E.; Kotsupiy, O.; Trofimova, E.; Zakabluk, G.; Chernonosov, A.; Koval, V.; Novikova, T. A mechanocomposite based on biogenic silica and green tea flavonoids modulates adaptability of strawberry microclones to in vitro and ex vitro conditions. J. Soil Sci. Plant Nutr. 2022, 1–16. [Google Scholar] [CrossRef]
- Ambros, E.V.; Toluzakova, S.Y.; Shrainer, L.S.; Trofimova, E.G.; Novikova, T.I. An innovative approach to ex vitro rooting and acclimatization of Fragaria × ananassa Duch. microshoots using a biogenic silica and green-tea-catechin-based mechanocomposite. In Vitro Cell. Dev. Biol. Plant 2018, 54, 436–443. [Google Scholar] [CrossRef]
- Ambros, E.V.; Kotsupy, O.V.; Karpova, E.A.; Trofimova, E.G.; Zaytseva, Y.G.; Novikova, T.I. In vitro adaptive responses of Fragaria ananassa Duch. plantlets induced by the mechanocomposite based on amorphous silica and flavonoids of green tea. Theor. Prikl. Ecol. 2019, 4, 116–122. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-Ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Rahman, A.; Wallis, C.M.; Uddin, W. Silicon-induced systemic defense responses in perennial ryegrass against infection by Magnaporthe oryzae. Phytopathology 2015, 105, 748–757. [Google Scholar] [CrossRef] [Green Version]
- Gengmao, Z.; Shihui, L.; Xing, S.; Yizhou, W.; Zipan, C. The role of silicon in physiology of the medicinal plant (Lonicera japonica L.) under salt stress. Sci. Rep. 2015, 5, 12696. [Google Scholar] [CrossRef] [Green Version]
- Hajiboland, R.; Moradtalab, N.; Eshaghi, Z.; Feizy, J. Effect of silicon supplementation on growth and metabolism of strawberry plants at three developmental stages. N. Z. J. Crop Hortic. Sci. 2018, 46, 144–161. [Google Scholar] [CrossRef]
- Narvekar, A.S.; Tharayil, N. Nitrogen fertilization influences the quantity, composition, and tissue association of foliar phenolics in strawberries. Front. Plant Sci. 2021, 12, 613839. [Google Scholar] [CrossRef]
- Mahmood, T.; Anwar, F.; Abbas, M.; Saari, N. Effect of maturity on phenolics (phenolic acids and flavonoids) profile of strawberry cultivars and mulberry species from Pakistan. Int. J. Mol. Sci. 2012, 13, 4591–4607. [Google Scholar] [CrossRef] [Green Version]
- Pešaković, M.; Milenković, S.; Đukić, D.; Mandić, L.; Karaklajić-Stajić, Ž.; Tomić, J.; Miletić, N. Phenolic composition and antioxidant capacity of integrated and conventionally grown strawberry (Fragaria × ananassa Duch.). Hort. Sci. 2016, 43, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, A.B.; Turker, A.U. Effects of regeneration enhancers on micropropagation of Fragaria vesca L. and phenolic content comparison of field-grown and in vitro-grown plant materials by liquid chromatography-electrospray tandem mass spectrometry (LC–ESI-MS/MS). Sci. Hortic. 2014, 169, 169–178. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Sousa, M.J.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Enhancement of nutritional and bioactive compounds by in vitro culture of wild Fragaria vesca L. vegetative parts. Food Chem. 2017, 235, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Hazarika, B.N. Morpho-physiological disorders in in vitro culture of plants. Sci. Hort. 2006, 108, 105–120. [Google Scholar] [CrossRef]
- Pospisilova, J.; Synková, H.; Haisel, D.; Semorádová, Š. Acclimation of plantlets to ex vitro conditions: Effects of air humidity, irradiance, CO2 concentration and abscisic acid (a review). Acta Hort. 2007, 748, 29–38. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Eveleigh, D.E. Culture methods and detection of glucanases in suspension cultures of wheat and barley. Can. J. Biochem. 1968, 46, 417–421. [Google Scholar] [CrossRef]
- Ambros, E.V.; Zaytseva, Y.G.; Krasnikov, A.A.; Novikova, T.I. Optimization of microshoots regeneration systems of Fragaria × ananassa (Rosaceae) genotypes perspectived for Siberian region. Rastit. Mir Aziat. Ross. 2017, 4, 73–80. [Google Scholar] [CrossRef]
- GOST R 53135-2008; Planting Material of Fruit, Berry, Subtropical, Nuts, Citrus Cultures and Tea. Technical Conditions. Standartinform: Moscow, Russia, 2009; 41p.
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kikowska, M.; Thiem, B.; Szopa, A.; Klimek-Szczykutowicz, M.; ·Rewers, M.; ·Sliwinska, E.; Ekiert, H. Comparative analysis of phenolic acids and flavonoids in shoot cultures of Eryngium alpinum L.: An endangered and protected species with medicinal value. Plant Cell Tiss. Organ Cult. 2019, 139, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Sanchez, A.; Monribot-Villanueva, J.L.; Cocotle-Ronzón, Y.; Martínez-Cruz, N.S.; Guerrero-Analco, J.A. Phenolic profile and antioxidant activity from wild and in vitro cultivated Rhynchostele rossii (Orchidaceae). Act. Bot. Mex. 2020, 127, e1665. [Google Scholar] [CrossRef]
- Karpova, E.A.; Nabieva, A.Y.; Fershalova, T.D. Leaf pigments and concentrations of phenolic compound in Begonia grandis plantlets obtained from the floral explants. Rend. Lincei. Sci. Fis. E Naturali. 2021, 32, 921–930. [Google Scholar] [CrossRef]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; de Paiva Foletto-Felipe, M.; Ferrarese-Filho, J.A.O. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Ralph, J.; Bunzel, M.; Marita, J.; Lu, F.; Kim, H.; Schatz, P.; Grabber, J.; Steinhart, H. Peroxidase-dependent cross-linking reactions of p-hydroxycinnamates in plant cell walls. Phytochem. Rev. 2004, 3, 79–96. [Google Scholar] [CrossRef]
- Hukkanen, A.T.; Kokko, H.I.; Buchala, A.J.; McDougall, G.J.; Stewart, D.; Karenlampi, S.O.; Karjalainen, R.O. Benzothiadiazole induces the accumulation of phenolics and improves resistance to powdery mildew in strawberries. J. Agric. Food Chem. 2007, 5, 1862–1870. [Google Scholar] [CrossRef] [Green Version]
- González Moreno, A.; de Cózar, A.; Prieto, P.; Domínguez, E.; Heredia, A. Radiationless mechanism of UV deactivation by cuticle phenolics in plants. Nat. Commun. 2022, 13, 1786. [Google Scholar] [CrossRef]
- Hura, T.; Tyrka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. QTLs for cell wall-bound phenolics in relation to the photosynthetic apparatus activity and leaf water status under drought stress at different growth stages of triticale. Mol. Genet. Genom. 2017, 292, 415–433. [Google Scholar] [CrossRef]
- Reyt, G.; Ramakrishna, P.; Salas-González, I.; Fujita, S.; Love, A.; Tiemessen, D.; Lapierre, C.; Morreel, K.; Calvo-Polanco, M.; Flis, P.; et al. Two chemically distinct root lignin barriers control solute and water balance. Nat. Commun. 2021, 12, 2320. [Google Scholar] [CrossRef]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Surech Kumar, C. Syringic acid (SA)—A review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure–antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Temocico, G.; Fierascu, I.; Ortan, A.; Babeanu, N.E. Fragaria genus: Chemical composition and biological activities. Molecules 2020, 25, 498. [Google Scholar] [CrossRef] [Green Version]
- Bernatoniene, J.; Kopustinskiene, D.M. The role of catechins in cellular responses to oxidative stress. Molecules. 2018, 23, 965. [Google Scholar] [CrossRef] [Green Version]
- Maksimovic, J.D.; Bogdanovic, J.; Maksimovic, V.; Nikolic, M. Silicon modulates the metabolism and utilization of phenolic compounds in cucumber (Cucumis sativus L.) grown at excess manganese. J. Plant Nutr. Soil Sci. 2007, 170, 739–744. [Google Scholar] [CrossRef]
- Summers, C.B.; Felton, G.W. Prooxidant effects of phenolic acids on the generalist herbivore Helicoverpa zea (Lepidoptera: Noctuidae): Potential mode of action for phenolic compounds in plant anti-herbivore chemistry. Insect. Biochem. Mol. Biol. 1994, 24, 943–953. [Google Scholar] [CrossRef]
- War, A.R.; Sharma, S.P.; Sharma, H.C. Differential induction of flavonoids in groundnut in response to Helicoverpa armigera and Aphis craccivora infestation. Int. J. Insect Sci. 2020, 8, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Yamshi, A.; Bajguz, A.; Hayat, S. The role of quercetin in plants. Plant Physiol. Biochem. 2021, 166, 10–19. [Google Scholar] [CrossRef]
- Shetty, R.; Jensen, B.; Shetty, N.P.; Hansen, M.; Hansen, C.W.; Starkey, K.R.; Jørgensen, H.J.L. Silicon induced resistance against powdery mildew of roses caused by Podosphaera pannosa. Plant Pathol. 2012, 61, 120–131. [Google Scholar] [CrossRef]
- Kårlund, A.; Hanhineva, K.; Lehtonen, M.; McDougall, G.J.; Stewart, D.; Karjalainen, R.O. Non-targeted metabolite profiling highlights the potential of strawberry leaves as a resource for specific bioactive compounds. J. Sci. Food Agric. 2017, 97, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpova, E.A.; Fershalova, T.D.; Petruk, A.A. Flavonoids in adaptation of Begonia grandis Dryander subsp. grandis introduced in West Siberia (Novosibirsk). J. Stress Physiol. Biochem. 2016, 12, 44–56. [Google Scholar]
- Fraser, D.P.; Sharma, A.; Fletcher, T.; Budge, S.; Moncrieff, C.; Dodd, A.N.; Franklin, K.A. UV-B antagonises shade avoidance and increases levels of the flavonoid quercetin in coriander (Coriandrum sativum). Sci. Rep. 2017, 7, 17758. [Google Scholar] [CrossRef] [Green Version]
- Duckstein, S.M.; Lotter, E.M.; Meyer, U.; Lindequist, U.; Stintzing, F.C. Phenolic constituents from Alchemilla vulgaris L. and Alchemilla mollis (Buser) Rothm. at different dates of harvest. Z. Naturforsch. C J. Biosci. 2012, 67, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Grochowski, D.M.; Skalicka-Woźniak, K.; Orhan, I.E.; Xiao, J.; Locatelli, M.; Piwowarski, J.P.; Granica, S.; Tomczyk, M. A comprehensive review of agrimoniin. Ann. N. Y. Acad. Sci. 2017, 1401, 166–180. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotsupiy, O.; Karpova, E.; Trofimova, E.; Novikova, T.; Ambros, E. Transformation of Strawberry Plants’ Phenolic Profile after Treatment with a Mechanocomposite Based on Silicon Chelates in the Course of Development under In Vitro, Ex Vitro, and In Vivo Conditions. Horticulturae 2023, 9, 157. https://doi.org/10.3390/horticulturae9020157
Kotsupiy O, Karpova E, Trofimova E, Novikova T, Ambros E. Transformation of Strawberry Plants’ Phenolic Profile after Treatment with a Mechanocomposite Based on Silicon Chelates in the Course of Development under In Vitro, Ex Vitro, and In Vivo Conditions. Horticulturae. 2023; 9(2):157. https://doi.org/10.3390/horticulturae9020157
Chicago/Turabian StyleKotsupiy, Olga, Evgeniya Karpova, Elena Trofimova, Tatyana Novikova, and Elena Ambros. 2023. "Transformation of Strawberry Plants’ Phenolic Profile after Treatment with a Mechanocomposite Based on Silicon Chelates in the Course of Development under In Vitro, Ex Vitro, and In Vivo Conditions" Horticulturae 9, no. 2: 157. https://doi.org/10.3390/horticulturae9020157