
Drought Stress in Lentil (Lens culinaris, Medik) and Approaches for Its Management


Abstract
:1. Introduction
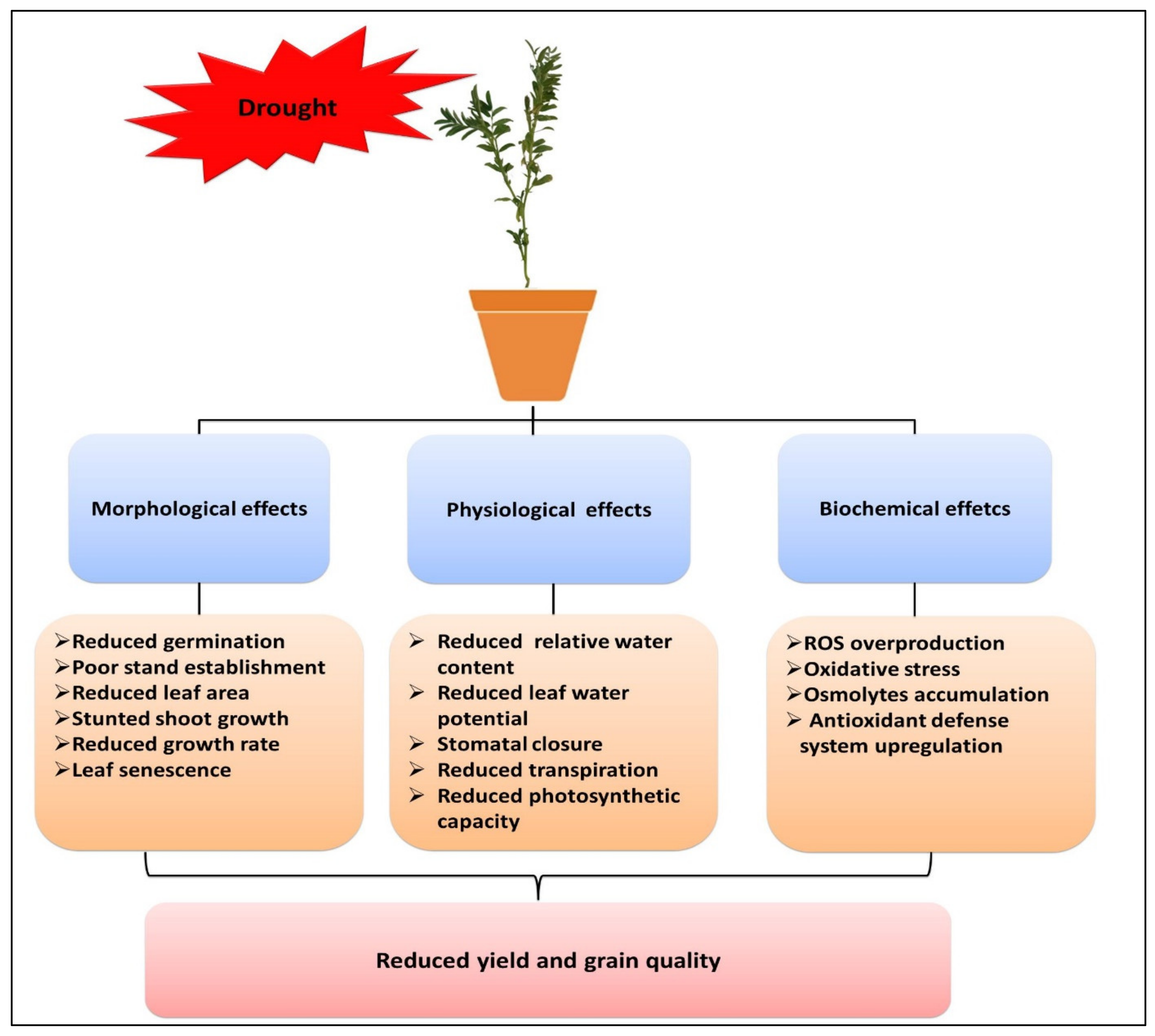
2. Morphological, Physiological, and Biochemical Effects of Drought Stress on Lentil
3. Management of Drought Stress in Lentil
3.1. Conventional Breeding for Improving Lentil Drought Tolerance
3.1.1. Exploitation of Genetic Variation for Selecting Potential Sources of Drought Tolerance
3.1.2. Potential Traits for Screening Drought Tolerance in Lentil
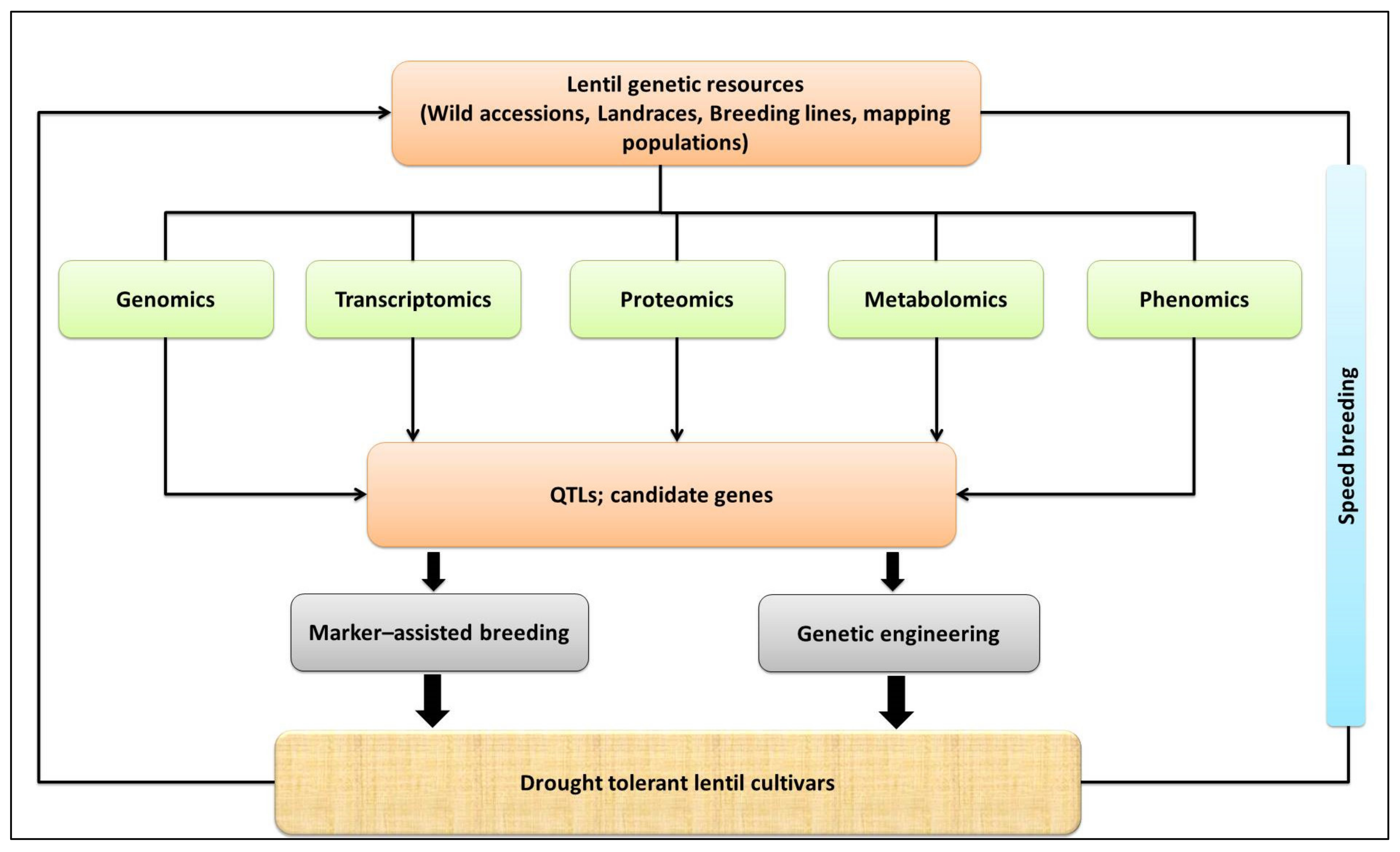
3.2. Biotechnological Approaches for Improving Lentil Drought Tolerance
3.2.1. Identification of Quantitative Trait Loci (QTLs)
3.2.2. High-Throughput Phenotyping
3.2.3. Transcriptomics
3.2.4. Proteomics
3.2.5. Metabolomics
3.2.6. Genetic Engineering
3.2.7. Integration of Biotechnological Tools with Speed Breeding
3.3. Agronomic Interventions to Induce Drought Stress Tolerance in Lentil
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Kaur, P.; Akhter, N.; Wani, S.H.; Saleem, F. Next-Generation Breeding Strategies for Climate-Ready Crops. Front. Plant Sci. 2021, 12, 620420. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAOSTAT. Available online: https://www.fao.org/faostat/en/#data (accessed on 29 September 2022).
- Idrissi, O.; Houasli, C.; Amamou, A.; Nsarellah, N. Lentil Genetic Improvement in Morocco: State of Art of the Program, Major Achievements and Perspectives. Moroc. J. Agric. Sci. 2020, 1, 9–13. Available online: https://techagro.org/index.php/MJAS/article/view/816 (accessed on 29 September 2022).
- Idrissi, O.; Sahri, A.; Houasli, C.; Nsarellah, N. Breeding Progress, Adaptation, and Stability for Grain Yield in Moroccan Lentil Improved Varieties. Crop Sci. 2019, 59, 925–936. [Google Scholar] [CrossRef]
- Jan, N.; Rather, A.M.-U.-D.; John, R.; Chaturvedi, P.; Ghatak, A.; Weckwerth, W.; Zargar, S.M.; Mir, R.A.; Khan, M.A.; Mir, R.R. Proteomics for Abiotic Stresses in Legumes: Present Status and Future Directions. Crit. Rev. Biotechnol. 2022, 2, 1–20. [Google Scholar] [CrossRef]
- Kaur, B.; Sandhu, K.S.; Kamal, R.; Kaur, K.; Singh, J.; Röder, M.S.; Muqaddasi, Q.H. Omics for the Improvement of Abiotic, Biotic, and Agronomic Traits in Major Cereal Crops: Applications, Challenges, and Prospects. Plants 2021, 10, 1989. [Google Scholar] [CrossRef]
- Idrissi, O.; Udupa, S.M.; Houasli, C.; De Keyser, E.; Van Damme, P.; De Riek, J. Genetic Diversity Analysis of Moroccan Lentil (Lens culinaris Medik.) Landraces Using Simple Sequence Repeat and Amplified Fragment Length Polymorphisms Reveals Functional Adaptation towards Agro-Environmental Origins. Plant Breed. 2015, 134, 322–332. [Google Scholar] [CrossRef]
- Sarker, A.; Erskine, W.; Singh, M. Variation in Shoot and Root Characteristics and Their Association with Drought Tolerance in Lentil Landraces. Genet. Resour. Crop Evol. 2005, 52, 89–97. [Google Scholar] [CrossRef]
- Lake, L.; Izzat, N.; Kong, T.; Sadras, V.O. High-Throughput Phenotyping of Plant Growth Rate to Screen for Waterlogging Tolerance in Lentil. J. Agron. Crop Sci. 2021, 207, 995–1005. [Google Scholar] [CrossRef]
- Wiraguna, E.; Malik, A.I.; Erskine, W. Waterlogging Tolerance in Lentil (Lens culinaris Medik. Subsp. Culinaris) Germplasm Associated with Geographic Origin. Genet. Resour. Crop Evol. 2017, 64, 579–586. [Google Scholar] [CrossRef]
- Bhandari, K.; Siddique, K.H.; Turner, N.C.; Kaur, J.; Singh, S.; Agrawal, S.K.; Nayyar, H. Heat Stress at Reproductive Stage Disrupts Leaf Carbohydrate Metabolism, Impairs Reproductive Function, and Severely Reduces Seed Yield in Lentil. J. Crop Improv. 2016, 30, 118–151. [Google Scholar] [CrossRef]
- Choukri, H.; Hejjaoui, K.; El-Baouchi, A.; El Haddad, N.; Smouni, A.; Maalouf, F.; Thavarajah, D.; Kumar, S. Heat and Drought Stress Impact on Phenology, Grain Yield, and Nutritional Quality of Lentil (Lens culinaris Medikus). Front. Nutr. 2020, 7, 596307. [Google Scholar] [CrossRef] [PubMed]
- El Haddad, N.; Rajendran, K.; Smouni, A.; Es-Safi, N.E.; Benbrahim, N.; Mentag, R.; Nayyar, H.; Maalouf, F.; Kumar, S. Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress. Agronomy 2020, 10, 1036. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Bhandari, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Impact of Heat Stress during Seed Filling on Seed Quality and Seed Yield in Lentil (Lens culinaris Medikus) Genotypes. J. Sci. Food Agric. 2018, 98, 5134–5141. [Google Scholar] [CrossRef] [PubMed]
- Erskine, W.; Tufail, M.; Russell, A.; Tyagi, M.C.; Rahman, M.M.; Saxena, M.C. Current and Future Strategies in Breeding Lentil for Resistance to Biotic and Abiotic Stresses. Euphytica 1993, 73, 127–135. [Google Scholar] [CrossRef]
- Ghimire, N.H.; Mandal, H.N. Genetic Variability, Genetic Advance, Correlation and Heritability of Cold Tolerance Lentil (Lens culinaris Medic.) Genotypes at High Hill of Nepal. Int. J. Adv. Res. Biol. Sci. 2019, 6, 1–10. [Google Scholar]
- Fardus, J.; Hossain, M.S.; Fujita, M. Modulation of the Antioxidant Defense System by Exogenous L-Glutamic Acid Application Enhances Salt Tolerance in Lentil (Lens culinaris Medik.). Biomolecules 2021, 11, 587. [Google Scholar] [CrossRef]
- Muscolo, A.; Junker, A.; Klukas, C.; Weigelt-Fischer, K.; Riewe, D.; Altmann, T. Phenotypic and Metabolic Responses to Drought and Salinity of Four Contrasting Lentil Accessions. J. Exp. Bot. 2015, 66, 5467–5480. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Singh, C.K.; Tomar, R.S.S.; Sharma, S.; Karwa, S.; Pal, M.; Singh, V.; Sanwal, S.K.; Sharma, P.C. Genetics and Molecular Mapping for Salinity Stress Tolerance at Seedling Stage in Lentil (Lens culinaris Medik). Crop Sci. 2020, 60, 1254–1266. [Google Scholar] [CrossRef]
- Yasir, T.A.; Khan, A.; Skalicky, M.; Wasaya, A.; Rehmani, M.I.A.; Sarwar, N.; Mubeen, K.; Aziz, M.; Hassan, M.M.; Hassan, F.A. Exogenous Sodium Nitroprusside Mitigates Salt Stress in Lentil (Lens culinaris Medik.) by Affecting the Growth, Yield, and Biochemical Properties. Molecules 2021, 26, 2576. [Google Scholar] [CrossRef]
- Singh, D.; Singh, C.K.; Taunk, J.; Gaikwad, K.; Singh, V.; Sanwal, S.K.; Karwa, S.; Singh, D.; Sharma, P.C.; Yadav, R.K. Linking Genome Wide RNA Sequencing with Physio-Biochemical and Cytological Responses to Catalogue Key Genes and Metabolic Pathways for Alkalinity Stress Tolerance in Lentil (Lens culinaris Medikus). BMC Plant Biol. 2022, 22, 99. [Google Scholar] [CrossRef]
- Bansal, R.; Priya, S.; Dikshit, H.K.; Jacob, S.R.; Rao, M.; Bana, R.S.; Kumari, J.; Tripathi, K.; Kumar, A.; Kumar, S. Growth and Antioxidant Responses in Iron-Biofortified Lentil under Cadmium Stress. Toxics 2021, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Abdelrahman, M.; Tran, C.D.; Nguyen, K.H.; Chu, H.D.; Watanabe, Y.; Fujita, M.; Tran, L.-S.P. Modulation of Osmoprotection and Antioxidant Defense by Exogenously Applied Acetate Enhances Cadmium Stress Tolerance in Lentil Seedlings. Environ. Pollut. 2022, 308, 119687. [Google Scholar] [CrossRef] [PubMed]
- Erskine, W.; Saxena, N.P.; Saxena, M.C. Iron Deficiency in Lentil: Yield Loss and Geographic Distribution in a Germplasm Collection. Plant Soil 1993, 151, 249–254. [Google Scholar] [CrossRef]
- Srivastava, S.P.; Bhandari, T.M.S.; Yadav, C.R.; Joshi, M.; Erskine, W. Boron Deficiency in Lentil: Yield Loss and Geographic Distribution in a Germplasm Collection. Plant Soil 2000, 219, 147–151. [Google Scholar] [CrossRef]
- Kaur, S.; Cogan, N.O.; Stephens, A.; Noy, D.; Butsch, M.; Forster, J.W.; Materne, M. EST-SNP Discovery and Dense Genetic Mapping in Lentil (Lens culinaris Medik.) Enable Candidate Gene Selection for Boron Tolerance. Theor. Appl. Genet. 2014, 127, 703–713. [Google Scholar] [CrossRef]
- Rodda, M.S.; Sudheesh, S.; Javid, M.; Noy, D.; Gnanasambandam, A.; Slater, A.T.; Rosewarne, G.M.; Kaur, S. Breeding for Boron Tolerance in Lentil (Lens culinaris Medik.) Using a High-throughput Phenotypic Assay and Molecular Markers. Plant Breed. 2018, 137, 492–501. [Google Scholar] [CrossRef]
- Yau, S.-K.; Erskine, W. Diversity of Boron-Toxicity Tolerance in Lentil Growth and Yield. Genet. Resour. Crop Evol. 2000, 47, 55–62. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Anastasi, U.; Santonoceto, C.; Maggio, A. Effect of PEG-Induced Drought Stress on Seed Germination of Four Lentil Genotypes. J. Plant Interact. 2014, 9, 354–363. [Google Scholar] [CrossRef]
- Akter, S.; Jahan, I.; Hossain, M.A.; Hossain, M.A. Laboratory-and Field-Phenotyping for Drought Stress Tolerance and Diversity Study in Lentil (Lens culinaris Medik.). Phyton 2021, 90, 949–970. [Google Scholar] [CrossRef]
- Idrissi, O.; Houasli, C.; Udupa, S.M.; De Keyser, E.; Van Damme, P.; De Riek, J. Genetic Variability for Root and Shoot Traits in a Lentil (Lens culinaris Medik.) Recombinant Inbred Line Population and Their Association with Drought Tolerance. Euphytica 2015, 204, 693–709. [Google Scholar] [CrossRef]
- Gupta, D.; Dadu, R.H.; Sambasivam, P.; Bar, I.; Azad, M.; Beera, N.; Ford, R.; Biju, S. Conventional and Biotechnological Approaches for Targeted Trait Improvement in Lentil. In Accelerated Plant Breeding, Volume 3; Springer: Berlin/Heidelberg, Germany, 2020; pp. 67–107. [Google Scholar]
- Gupta, D.; Dadu, R.H.R.; Sambasivam, P.; Bar, I.; Singh, M.; Biju, S. Toward Climate-Resilient Lentils: Challenges and Opportunities. In Genomic Designing of Climate-Smart Pulse Crops; Springer: Berlin/Heidelberg, Germany, 2019; pp. 165–234. [Google Scholar]
- Saha, M.; Bandyopadhyay, P.K.; Sarkar, A.; Nandi, R.; Singh, K.C.; Sanyal, D. Understanding the Impacts of Sowing Time and Tillage in Optimizing the Micro-Environment for Rainfed Lentil (Lens culinaris Medik) Production in the Lower Indo-Gangetic Plain. J. Soil Sci. Plant Nutr. 2020, 20, 2536–2551. [Google Scholar] [CrossRef]
- Ben Ghoulam, S.; Zeroual, A.; Baidani, A.; Idrissi, O. Réponse au déficit hydrique progressif chez la lentille: Vers une différentiation morpho-physiologique entre des accessions sauvages (Lens orientalis), populations locales et lignées avancées (Lens culinaris). Botany 2022, 100, 33–46. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gupta, D. The Use of Infrared Thermal Imaging as a Non-Destructive Screening Tool for Identifying Drought-Tolerant Lentil Genotypes. Plant Physiol. Biochem. 2018, 127, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Singh, C.K.; Taunk, J.; Tomar, R.S.S.; Chaturvedi, A.K.; Gaikwad, K.; Pal, M. Transcriptome Analysis of Lentil (Lens culinaris Medikus) in Response to Seedling Drought Stress. BMC Genom. 2017, 18, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venugopalan, V.K.; Nath, R.; Sengupta, K.; Nalia, A.; Banerjee, S.; Chandran, M.A.S.; Ibrahimova, U.; Dessoky, E.S.; Attia, A.O.; Hassan, M.M.; et al. The Response of Lentil (Lens culinaris Medik.) to Soil Moisture and Heat Stress Under Different Dates of Sowing and Foliar Application of Micronutrients. Front. Plant Sci. 2021, 12, 679469. [Google Scholar] [CrossRef] [PubMed]
- Morgil, H.; Tardu, M.; Cevahir, G.; Kavakli, İ.H. Comparative RNA-Seq Analysis of the Drought-Sensitive Lentil (Lens culinaris) Root and Leaf under Short- and Long-Term Water Deficits. Funct. Integr. Genom. 2019, 19, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Venugopalan, V.; Nath, R.; Sengupta, K.; Pal, A.; Banerjee, S.; Banerjee, P.; Chandran, M.; Roy, S.; Sharma, L.; Hossain, A. Foliar Spray of Micronutrients Alleviates Heat and Moisture Stress in Lentil (Lens culinaris Medik) Grown Under Rainfed Field Conditions. Front. Plant Sci. 2022, 13, 47743. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gupta, D. Silicon Improves Seed Germination and Alleviates Drought Stress in Lentil Crops by Regulating Osmolytes, Hydrolytic Enzymes and Antioxidant Defense System. Plant Physiol. Biochem. 2017, 119, 250–264. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Akbar, M.N.; Iftikhar, Y.; Abbas, M.; Zahid, A.; Fahad, S.; Datta, R.; Ali, M.; Elgorban, A.M.; Ansari, M.J. Rhizobacteria Inoculation and Caffeic Acid Alleviated Drought Stress in Lentil Plants. Sustainability 2021, 13, 9603. [Google Scholar] [CrossRef]
- Idrissi, O.; Chafika, H.; Nsarellah, N. Comparaison de lignées avancées de lentille sous stress hydrique durant la phase de floraison et formation des gousses. Nat. Technol. 2013, 10, 53–61. [Google Scholar]
- Choukri, H.; El Haddad, N.; Aloui, K.; Hejjaoui, K.; El-Baouchi, A.; Smouni, A.; Maalouf, F.; Kumar, S. Effect of High Temperature Stress During the Reproductive Stage on Grain Yield and Nutritional Quality of Lentil (Lens culinaris Medikus). Front. Nutr. 2022, 9, 857469. [Google Scholar] [CrossRef] [PubMed]
- El Haddad, N.; Choukri, H.; Ghanem, M.E.; Smouni, A.; Mentag, R.; Rajendran, K.; Hejjaoui, K.; Maalouf, F.; Kumar, S. High-Temperature and Drought Stress Effects on Growth, Yield and Nutritional Quality with Transpiration Response to Vapor Pressure Deficit in Lentil. Plants 2021, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Kumari, V.V.; Roy, A.; Vijayan, R.; Banerjee, P.; Verma, V.C.; Nalia, A.; Pramanik, M.; Mukherjee, B.; Ghosh, A.; Reja, M. Drought and Heat Stress in Cool-Season Food Legumes in Sub-Tropical Regions: Consequences, Adaptation, and Mitigation Strategies. Plants 2021, 10, 1038. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Farooq, M. The Challenge of Drought Stress for Grain Legumes and Options for Improvement. Arch. Agron. Soil Sci. 2022, 68, 1601–1618. [Google Scholar] [CrossRef]
- Hojjat, S.S.; Ganjali, A. The Effect of Silver Nanoparticle on Lentil Seed Germination under Drought Stress. Int. J. Farming Allied Sci. 2016, 5, 208–212. [Google Scholar]
- Shrestha, R.; Turner, N.C.; Siddique, K.H.M.; Turner, D.W.; Speijers, J. A Water Deficit during Pod Development in Lentils Reduces Flower and Pod Numbers but Not Seed Size. Aust. J. Agric. Res. 2006, 57, 427. [Google Scholar] [CrossRef]
- Gorim, L.Y.; Vandenberg, A. Evaluation of Wild Lentil Species as Genetic Resources to Improve Drought Tolerance in Cultivated Lentil. Front. Plant Sci. 2017, 8, 1129. [Google Scholar] [CrossRef]
- Hamdi, A.; Erskine, W. Reaction of Wild Species of the Genus Lens to Drought. Euphytica 1996, 91, 173–179. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Effects of Drought, Heat and Their Interaction on the Growth, Yield and Photosynthetic Function of Lentil (Lens culinaris Medikus) Genotypes Varying in Heat and Drought Sensitivity. Front. Plant Sci. 2017, 8, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Fujita, M. Drought Stress Responses in Plants, Oxidative Stress, and Antioxidant Defense. In Climate Change and Plant Abiotic Stress Tolerance; Tuteja, N., Gill, S.S., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; pp. 209–237. [Google Scholar] [CrossRef]
- Turner, N.C.; Wright, G.C.; Siddique, K.H.M. Adaptation of Grain Legumes (Pulses) to Water-Limited Environments. Adv. Agron. 2001, 71, 193–231. [Google Scholar]
- Varshney, R.; Barmukh, R.; Roorkiwal, M.; Qi, Y.; Kholova, J.; Tuberosa, R.; Reynolds, M.; Tardieu, F.; Siddique, K. Breeding Custom-Designed Crops for Improved Drought Adaptation. Adv. Genet. 2021, 2, e202100017. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Shah, W.; Ullah, S.; Ali, S.; Idrees, M.; Khan, M.N.; Ali, K.; Khan, A.; Ali, M.; Younas, F. Effect of Exogenous Alpha-Tocopherol on Physio-Biochemical Attributes and Agronomic Performance of Lentil (Lens culinaris Medik.) under Drought Stress. PLoS ONE 2021, 16, e0248200. [Google Scholar] [CrossRef]
- Mishra, B.K.; Srivastava, J.P.; Lal, J.P.; Sheshshayee, M.S. Physiological and Biochemical Adaptations in Lentil Genotypes under Drought Stress. Russ. J. Plant Physiol. 2016, 63, 695–708. [Google Scholar] [CrossRef]
- Sinha, R.; Pal, A.K.; Singh, A.K. Physiological, Biochemical and Molecular Responses of Lentil (Lens culinaris Medik.) Genotypes under Drought Stress. Indian J. Plant Physiol. 2018, 23, 772–784. [Google Scholar] [CrossRef]
- Abdela, A.A.; Barka, G.D.; Degefu, T. Co-Inoculation Effect of Mesorhizobium Ciceri and Pseudomonas Fluorescens on Physiological and Biochemical Responses of Kabuli Chickpea (Cicer arietinum L.) during Drought Stress. Plant Physiol. Rep. 2020, 25, 359–369. [Google Scholar] [CrossRef]
- Hosseini, S.Z.; Ismaili, A.; Nazarian-Firouzabadi, F.; Fallahi, H.; Nejad, A.R.; Sohrabi, S.S. Dissecting the Molecular Responses of Lentil to Individual and Combined Drought and Heat Stresses by Comparative Transcriptomic Analysis. Genomics 2021, 113, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Erskine, W. Seed-Size Effects on Lentil (Lens culinaris) Yield Potential and Adaptation to Temperature and Rainfall in West Asia. J. Agric. Sci. 1996, 126, 335–341. [Google Scholar] [CrossRef]
- Sánchez-Gómez, D.; Cervera, M.T.; Escolano-Tercero, M.A.; Vélez, M.D.; de María, N.; Diaz, L.; Sánchez-Vioque, R.; Aranda, I.; Guevara, M.Á. Drought Escape Can Provide High Grain Yields under Early Drought in Lentils. Theor. Exp. Plant Physiol. 2019, 31, 273–286. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Bhandari, K.; Kumar, S.; Kumar, J.; Vara Prasad, P.V.; Siddique, K.H.; Nayyar, H. Influence of Drought and Heat Stress, Applied Independently or in Combination during Seed Development, on Qualitative and Quantitative Aspects of Seeds of Lentil (Lens culinaris Medikus) Genotypes, Differing in Drought Sensitivity. Plant Cell Environ. 2019, 42, 198–211. [Google Scholar] [CrossRef] [Green Version]
- Serraj, R. Effects of Drought Stress on Legume Symbiotic Nitrogen Fixation: Physiological Mechanisms. Indian J. Exp. Biol. 2003, 41, 1136–1141. [Google Scholar]
- Idrissi, O.; Udupa, M.S.; De Keyser, E.; Van Damme, P.; De Riek, J. Functional Genetic Diversity Analysis and Identification of Associated Simple Sequence Repeats and Amplified Fragment Length Polymorphism Markers to Drought Tolerance in Lentil (Lens culinaris ssp. Culinaris Medicus) Landraces. Plant Mol. Biol. Report. 2016, 34, 659–680. [Google Scholar] [CrossRef]
- Houasli, C.; Sahri, A.; Nsarellah, N.; Idrissi, O. Chickpea (Cicer arietinum L.) Breeding in Morocco: Genetic Gain and Stability of Grain Yield and Seed Size under Winter Planting Conditions. Euphytica 2021, 217, 159. [Google Scholar] [CrossRef]
- Idrissi, O.; Sakr, B.; Dahan, R.; Houasli, C.; Nsarellah, N.; Udupa, S.M.; Sarker, A. Registration of ‘Chakkouf’ Lentil in Morocco. J. Plant Regist. 2012, 6, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Saker, B. Amélioration Génétique de La Lentille. In La Création Variétale à l’INRA: Méthodes, Acquis et Perspectives|INRA; 2005; Available online: https://www.inra.org.ma/fr/content/la-cr%C3%A9ation-vari%C3%A9tale-%C3%A0-linra-m%C3%A9thodes-acquis-et-perspectives-full-text (accessed on 18 July 2022).
- Babayeva, S.; Akparov, Z.; Damania, A.; Izzatullayeva, V.; Abbasov, M. Genetic Diversity for Drought Tolerance in Lentils from Central Asia and the Caucasus: CACLentil. Albanian J. Agric. Sci. 2014, 13, 1–8. [Google Scholar]
- Coyne, C.J.; Kumar, S.; Wettberg, E.J.B.; Marques, E.; Berger, J.D.; Redden, R.J.; Ellis, T.H.N.; Brus, J.; Zablatzká, L.; Smýkal, P. Potential and Limits of Exploitation of Crop Wild Relatives for Pea, Lentil, and Chickpea Improvement. Legume Sci. 2020, 2, e36. [Google Scholar] [CrossRef] [Green Version]
- Dash, A.P.; De, D.K.; Mohanty, S.; Lenka, D. Screening of Lentil (Lens culinaris Medik.) Genotypes and Correlation Analysis under PEG Imposed Water Stress Condition. Int. J. Bio-Resour. Stress Manag. 2017, 8, 539–547. [Google Scholar] [CrossRef]
- Kumar, J.; Basu, P.S.; Srivastava, E.; Chaturvedi, S.K.; Nadarajan, N.; Kumar, S. Phenotyping of Traits Imparting Drought Tolerance in Lentil. Crop Pasture Sci. 2012, 63, 547. [Google Scholar] [CrossRef]
- Priya, S.; Bansal, R.; Kumar, G.; Dikshit, H.K.; Kumari, J.; Pandey, R.; Singh, A.K.; Tripathi, K.; Singh, N.; Kumari, N.K.P.; et al. Root Trait Variation in Lentil (Lens culinaris Medikus) Germplasm under Drought Stress. Plants 2021, 10, 2410. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, S.; Basandrai, A.K.; Basandrai, D.; Malhotra, N.; Saxena, D.R.; Gupta, D.; Sarker, A.; Singh, K. Evaluation and Identification of Wild Lentil Accessions for Enhancing Genetic Gains of Cultivated Varieties. PLoS ONE 2020, 15, e0229554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Sharma, S.K.; Singh, B.; Malhotra, N.; Chandora, R.; Sarker, A.; Singh, K.; Gupta, D. Widening the Genetic Base of Cultivated Gene Pool Following Introgression from Wild Lens Taxa. Plant Breed. 2018, 137, 470–485. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Dikshit, H.K.; Singh, R. A New Phenotyping Technique for Screening for Drought Tolerance in Lentil (Lens culinaris Medik.). Plant Breed. 2013, 132, 185–190. [Google Scholar] [CrossRef]
- Ullah, A.; Romdhane, L.; Rehman, A.; Farooq, M. Adequate Zinc Nutrition Improves the Tolerance against Drought and Heat Stresses in Chickpea. Plant Physiol. Biochem. 2019, 143, 11–18. [Google Scholar] [CrossRef]
- Silim, S.N.; Saxena, M.C.; Erskine, W. Adaptation of Lentil to the Mediterranean Environment. I. Factors Affecting Yield Under Drought Conditions. Exp. Agric. 1993, 29, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, R.; Turner, N.C.; Siddique, K.H.M.; Turner, D.W. Physiological and Seed Yield Responses to Water Deficits among Lentil Genotypes from Diverse Origins. Aust. J. Agric. Res. 2006, 57, 903. [Google Scholar] [CrossRef]
- Akter, S.; Jahan, I.; Hossain, M.A.; Hossain, M.A. Variability for Agromorphological Traits, Genetic Parameters, Correlation and Path Coefficient Analyses in Lentil (Lens culinaris Medik.). Res. Plant Biol. 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Saxena, M.C. Plant Morphology, Anatomy and Growth Habit. In The Lentil: Botany, Production and Uses; Erskine, W., Muehlbauer, F.J., Sarker, A., Sharma, B., Eds.; CABI Press: Wallingford, UK, 2009; pp. 34–46. [Google Scholar]
- Gorim, L.Y.; Vandenberg, A. Variation in Total Root Length and Root Diameter of Wild and Cultivated Lentil Grown under Drought and Re-Watered Conditions. Plant Genet. Resour. Charact. Util. 2019, 17, 45–53. [Google Scholar] [CrossRef]
- Gorim, L.Y.; Vandenberg, A. Can Wild Lentil Genotypes Help Improve Water Use and Transpiration Efficiency in Cultivated Lentil? Plant Genet. Resour. Charact. Util. 2018, 16, 459–468. [Google Scholar] [CrossRef]
- Mishra, B.K.; Srivastava, J.P.; Lal, J.P. Drought Resistance in Lentil (Lens culinaris Medik.) in Relation to Morphological, Physiological Parameters and Phenological Developments. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2288–2304. [Google Scholar] [CrossRef]
- Ashraf, M.; Bokhari, M.H.; Chishti, S.N. Variation in Osmotic Adjustment of Accessions of Lentil (Lens culinaris Medic.) in Response to Drought Stress. Acta Bot. Neerl. 1992, 41, 51–62. [Google Scholar] [CrossRef]
- Ashraf, M. Thermotolerance in Plants: Potential Physio-Biochemical and Molecular Markers for Crop Improvement. Environ. Exp. Bot. 2021, 186, 104454. [Google Scholar] [CrossRef]
- Shunmugam, A.; Kannan, U.; Jiang, Y.; Daba, K.; Gorim, L. Physiology Based Approaches for Breeding of Next-Generation Food Legumes. Plants 2018, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Allahmoradi, P.; Mansourifar, C.; Saiedi, M. Effect of Different Water Deficiency Levels on Some Antioxidants at Different Growth Stages of Lentil (Lens culinaris L.). Adv. Environ. Biol. 2013, 7, 535–543. [Google Scholar]
- Ahmadi, A.; Dehaghi, M.A.; Fotokian, M.H.; Sedghi, M.; Far, C.M. Evaluation of stress tolerance indices in a number of advanced genotypes of lentil (Lens culinaris medik) under rainfed and low irrigation conditions. Plant Arch. 2019, 19, 490–499. [Google Scholar]
- Siahsar, B.A.; Ganjali, S.; Allahdoo, M. Evaluation of Drought Tolerance Indices and Their Relationship with Grain Yield of Lentil Lines in Drought-Stressed and Irrigated Environments. Aust. J. Basic Appl. Sci. 2010, 4, 4336–4346. [Google Scholar]
- Dwivedi, S.L.; Siddique, K.H.M.; Farooq, M.; Thornton, P.K.; Ortiz, R. Using Biotechnology-Led Approaches to Uplift Cereal and Food Legume Yields in Dryland Environments. Front. Plant Sci. 2018, 9, 1249. [Google Scholar] [CrossRef]
- Bett, K.; Ramsay, L.; Chan, C.; Sharpe, A.G.; Cook, D.R.; Varma, R. OP06: The Lentil Genome–from the Sequencer to the Field. In Proceedings of the PAG XXIV: Plant and Animal Genomics Conference, San Diego, CA, USA, 18–20 April 2016. Available online: https://mel.cgiar.org/reporting/download/hash/gJFbVXUc (accessed on 18 August 2022).
- Ramsay, L.; Koh, C.S.; Kagale, S.; Gao, D.; Kaur, S.; Haile, T.; Gela, T.S.; Chen, L.-A.; Cao, Z.; Konkin, D.J. Genomic Rearrangements Have Consequences for Introgression Breeding as Revealed by Genome Assemblies of Wild and Cultivated Lentil Species. bioRxiv 2021. [preprint]. [Google Scholar] [CrossRef]
- Saha, G.C.; Sarker, A.; Chen, W.; Vandemark, G.J.; Muehlbauer, F.J. Inheritance and Linkage Map Positions of Genes Conferring Resistance to Stemphylium Blight in Lentil. Crop Sci. 2010, 50, 1831–1839. [Google Scholar] [CrossRef]
- Saha, G.C.; Sarker, A.; Chen, W.; Vandemark, G.J.; Muehlbauer, F.J. Inheritance and Linkage Map Positions of Genes Conferring Agromorphological Traits in Lens culinaris Medik. Int. J. Agron. 2013, 2013, 618926. [Google Scholar] [CrossRef] [Green Version]
- Dadu, R.H.R.; Bar, I.; Ford, R.; Sambasivam, P.; Croser, J.; Ribalta, F.; Kaur, S.; Sudheesh, S.; Gupta, D. Lens Orientalis Contributes Quantitative Trait Loci and Candidate Genes Associated with Ascochyta Blight Resistance in Lentil. Front. Plant Sci. 2021, 12, 703283. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Taylor, P.W.J.; Inder, P.; Phan, H.T.T.; Ellwood, S.R.; Mathur, P.N.; Sarker, A.; Ford, R. Integration of EST-SSR Markers of Medicago Truncatula into Intraspecific Linkage Map of Lentil and Identification of QTL Conferring Resistance to Ascochyta Blight at Seedling and Pod Stages. Mol. Breed. 2012, 30, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Polanco, C.; Sáenz de Miera, L.E.; González, A.I.; García, P.; Fratini, R.; Vaquero, F.; Vences, F.J.; Pérez de la Vega, M. Construction of a High-Density Interspecific (Lens culinaris x L. Odemensis) Genetic Map Based on Functional Markers for Mapping Morphological and Agronomical Traits, and QTLs Affecting Resistance to Ascochyta in Lentil. PLoS ONE 2019, 14, e0214409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubeena; Taylor, P.W.J.; Ades, P.K.; Ford, R. QTL Mapping of Resistance in Lentil (Lens culinaris ssp. Culinaris) to Ascochyta Blight (Ascochyta Lentis). Plant Breed. 2006, 125, 506–512. [Google Scholar]
- Sudheesh, S.; Rodda, M.S.; Davidson, J.; Javid, M.; Stephens, A.; Slater, A.T.; Cogan, N.O.; Forster, J.W.; Kaur, S. SNP-Based Linkage Mapping for Validation of QTLs for Resistance to Ascochyta Blight in Lentil. Front. Plant Sci. 2016, 7, 1604. [Google Scholar] [CrossRef] [Green Version]
- Hamwieh, A.; Udupa, S.M.; Choumane, W.; Sarker, A.; Dreyer, F.; Jung, C.; Baum, M. A Genetic Linkage Map of Lens Sp. Based on Microsatellite and AFLP Markers and the Localization of Fusarium Vascular Wilt Resistance. Theor. Appl. Genet. 2005, 110, 669–677. [Google Scholar] [CrossRef]
- Gela, T.; Ramsay, L.; Haile, T.A.; Vandenberg, A.; Bett, K. Identification of Anthracnose Race 1 Resistance Loci in Lentil by Integrating Linkage Mapping and Genome-wide Association Study. Plant Genome 2021, 14, e20131. [Google Scholar] [CrossRef]
- Gela, T.S.; Koh, C.S.; Caron, C.T.; Chen, L.-A.; Vandenberg, A.; Bett, K.E. QTL Mapping of Lentil Anthracnose (Colletotrichum lentis) Resistance from Lens Ervoides Accession IG 72815 in an Interspecific RIL Population. Euphytica 2021, 217, 70. [Google Scholar] [CrossRef]
- Ma, Y.; Marzougui, A.; Coyne, C.J.; Sankaran, S.; Main, D.; Porter, L.D.; Mugabe, D.; Smitchger, J.A.; Zhang, C.; Amin, M. Dissecting the Genetic Architecture of Aphanomyces Root Rot Resistance in Lentil by QTL Mapping and Genome-Wide Association Study. Int. J. Mol. Sci. 2020, 21, 2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, C.K.; Singh, D.; Tomar, R.S.S.; Karwa, S.; Upadhyaya, K.C.; Pal, M. Molecular Mapping of Aluminium Resistance Loci Based on Root Re-Growth and Al-Induced Fluorescent Signals (Callose Accumulation) in Lentil (Lens culinaris Medikus). Mol. Biol. Rep. 2018, 45, 2103–2113. [Google Scholar] [CrossRef]
- Singh, C.K.; Singh, D.; Sharma, S.; Chandra, S.; Tomar, R.S.S.; Kumar, A.; Upadhyaya, K.C.; Pal, M. Mechanistic Association of Quantitative Trait Locus with Malate Secretion in Lentil (Lens culinaris Medikus) Seedlings under Aluminium Stress. Plants 2021, 10, 1541. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Singh, C.K.; Singh Tomar, R.S.; Pal, M. Genetics and Molecular Mapping of Heat Tolerance for Seedling Survival and Pod Set in Lentil. Crop Sci. 2017, 57, 3059–3067. [Google Scholar] [CrossRef]
- Ates, D.; Sever, T.; Aldemir, S.; Yagmur, B.; Temel, H.Y.; Kaya, H.B.; Alsaleh, A.; Kahraman, A.; Ozkan, H.; Vandenberg, A. Identification QTLs Controlling Genes for Se Uptake in Lentil Seeds. PLoS ONE 2016, 11, e0149210. [Google Scholar]
- Ates, D.; Aldemir, S.; Yagmur, B.; Kahraman, A.; Ozkan, H.; Vandenberg, A.; Tanyolac, M.B. QTL Mapping of Genome Regions Controlling Manganese Uptake in Lentil Seed. G3 Genes Genomes Genet. 2018, 8, 1409–1416. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Singh, C.K.; Taunk, J.; Tomar, R.S.S. Genetic Analysis and Molecular Mapping of Seedling Survival Drought Tolerance Gene in Lentil (Lens culinaris Medikus). Mol. Breed. 2016, 36, 58. [Google Scholar] [CrossRef]
- Varshney, R.K.; Thudi, M.; Nayak, S.N.; Gaur, P.M.; Kashiwagi, J.; Krishnamurthy, L.; Jaganathan, D.; Koppolu, J.; Bohra, A.; Tripathi, S. Genetic Dissection of Drought Tolerance in Chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2014, 127, 445–462. [Google Scholar] [CrossRef]
- Bharadwaj, C.; Tripathi, S.; Soren, K.R.; Thudi, M.; Singh, R.K.; Sheoran, S.; Roorkiwal, M.; Patil, B.S.; Chitikineni, A.; Palakurthi, R. Introgression of “QTL-hotspot” Region Enhances Drought Tolerance and Grain Yield in Three Elite Chickpea Cultivars. Plant Genome 2021, 14, e20076. [Google Scholar] [CrossRef]
- Varshney, R.K.; Gaur, P.M.; Chamarthi, S.K.; Krishnamurthy, L.; Tripathi, S.; Kashiwagi, J.; Samineni, S.; Singh, V.K.; Thudi, M.; Jaganathan, D. Fast-track Introgression of “QTL-hotspot” for Root Traits and Other Drought Tolerance Traits in JG 11, an Elite and Leading Variety of Chickpea. Plant Genome 2013, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Idrissi, O.; Udupa, S.M.; De Keyser, E.; McGee, R.J.; Coyne, C.J.; Saha, G.C.; Muehlbauer, F.J.; Van Damme, P.; De Riek, J. Identification of Quantitative Trait Loci Controlling Root and Shoot Traits Associated with Drought Tolerance in a Lentil (Lens culinaris Medik.) Recombinant Inbred Line Population. Front. Plant Sci. 2016, 7, 1174. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Eltaher, S.; Alqudah, A.M.; Belamkar, V.; Baenziger, P.S. Combined GWAS and QTL Mapping Revealed Candidate Genes and SNP Network Controlling Recovery and Tolerance Traits Associated with Drought Tolerance in Seedling Winter Wheat. Genomics 2022, 114, 110358. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, M.; Fakheri, B.A.; Aghnoum, R.; Mahdi Nezhad, N.; Ataei, R. GWAS Analysis in Spring Barley (Hordeum Vulgare L.) for Morphological Traits Exposed to Drought. PLoS ONE 2018, 13, e0204952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Feng, H.; Wu, D.; Yan, S.; Zhang, P.; Wang, W.; Zhang, J.; Ye, J.; Dai, G.; Fan, Y. Using High-Throughput Multiple Optical Phenotyping to Decipher the Genetic Architecture of Maize Drought Tolerance. Genome Biol. 2021, 22, 185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Wang, X.; Li, Y.; Li, Y.; Gou, Z.; Zhao, X.; Hong, H.; Ren, H.; Qi, X. Identification of Genes for Drought Resistance and Prediction of Gene Candidates in Soybean Seedlings Based on Linkage and Association Mapping. Crop J. 2021, 10, 830–839. [Google Scholar] [CrossRef]
- Singh, A.; Dikshit, H.K.; Mishra, G.P.; Aski, M.; Kumar, S. Association Mapping for Grain Diameter and Weight in Lentil Using SSR Markers. Plant Gene 2019, 20, 100204. [Google Scholar] [CrossRef]
- Gupta, P.K.; Kulwal, P.L.; Jaiswal, V. Association Mapping in Plants in the Post-GWAS Genomics Era. Adv. Genet. 2019, 104, 75–154. [Google Scholar]
- Chen, Y.; Palta, J.A.; Wu, P.; Siddique, K.H. Crop Root Systems and Rhizosphere Interactions. Plant Soil 2019, 439, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Marsh, J.I.; Hu, H.; Gill, M.; Batley, J.; Edwards, D. Crop Breeding for a Changing Climate: Integrating Phenomics and Genomics with Bioinformatics. Theor. Appl. Genet. 2021, 134, 1677–1690. [Google Scholar] [CrossRef]
- Yang, W.; Feng, H.; Zhang, X.; Zhang, J.; Doonan, J.H.; Batchelor, W.D.; Xiong, L.; Yan, J. Crop Phenomics and High-Throughput Phenotyping: Past Decades, Current Challenges, and Future Perspectives. Mol. Plant 2020, 13, 187–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Yang, W.; Chang, Y.; Ma, X.; Tu, H.; Xiong, F.; Jiang, N.; Feng, H.; Huang, C.; Yang, P. Genome-Wide Association Studies of Image Traits Reveal Genetic Architecture of Drought Resistance in Rice. Mol. Plant 2018, 11, 789–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Tu, H.; Bai, B.; Yang, C.; Zhao, B.; Guo, Z.; Liu, Q.; Zhao, H.; Yang, W.; Xiong, L. Combining UAV-RGB High-Throughput Field Phenotyping and Genome-Wide Association Study to Reveal Genetic Variation of Rice Germplasms in Dynamic Response to Drought Stress. New Phytol. 2021, 232, 440–455. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative Aerial and Ground Based High Throughput Phenotyping for the Genetic Dissection of NDVI as a Proxy for Drought Adaptive Traits in Durum Wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Feng, H.; Guo, C.; Yang, S.; Huang, W.; Xiong, X.; Liu, J.; Chen, G.; Liu, Q.; Xiong, L. High-Throughput Phenotyping Accelerates the Dissection of the Dynamic Genetic Architecture of Plant Growth and Yield Improvement in Rapeseed. Plant Biotechnol. J. 2020, 18, 2345–2353. [Google Scholar] [CrossRef]
- Rane, J.; Raina, S.K.; Govindasamy, V.; Bindumadhava, H.; Hanjagi, P.; Giri, R.; Jangid, K.K.; Kumar, M.; Nair, R.M. Use of Phenomics for Differentiation of Mungbean (Vigna Radiata L. Wilczek) Genotypes Varying in Growth Rates per Unit of Water. Front. Plant Sci. 2021, 12, 692534. [Google Scholar] [CrossRef] [PubMed]
- Sivasakthi, K.; Thudi, M.; Tharanya, M.; Kale, S.M.; Kholová, J.; Halime, M.H.; Jaganathan, D.; Baddam, R.; Thirunalasundari, T.; Gaur, P.M. Plant Vigour QTLs Co-Map with an Earlier Reported QTL Hotspot for Drought Tolerance While Water Saving QTLs Map in Other Regions of the Chickpea Genome. BMC Plant Biol. 2018, 18, 29. [Google Scholar] [CrossRef]
- Marzougui, A.; Ma, Y.; Zhang, C.; McGee, R.J.; Coyne, C.J.; Main, D.; Sankaran, S. Advanced Imaging for Quantitative Evaluation of Aphanomyces Root Rot Resistance in Lentil. Front. Plant Sci. 2019, 10, 383. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, R.; Kahrood, H.V.; Dimech, A.M.; Noy, D.M.; Rosewarne, G.M.; Smith, K.F.; Cogan, N.O.I.; Kaur, S. Development and Application of Image-Based High-Throughput Phenotyping Methodology for Salt Tolerance in Lentils. Agronomy 2020, 10, 1992. [Google Scholar] [CrossRef]
- Dissanayake, R.; Cogan, N.O.; Smith, K.F.; Kaur, S. Application of Genomics to Understand Salt Tolerance in Lentil. Genes 2021, 12, 332. [Google Scholar] [CrossRef]
- Patrignani, A.; Ochsner, T.E. Canopeo: A Powerful New Tool for Measuring Fractional Green Canopy Cover. Agron. J. 2015, 107, 2312–2320. [Google Scholar] [CrossRef] [Green Version]
- Zeroual, A.; Baidani, A.; Idrissi, O. Use of canopeo, a smart phone based application, as a nondestructive and simple tool to estimate drought tolerance in lentil. In Proceedings of the ISPEC 9th International Conference on Agriculture, Animal Sciences and Rural Development, Burdur, Turkey, 19–20 March 2022. [Google Scholar]
- Idrissi, O.; Draye, X. High-Throughput Phenotyping for Drought Tolerance-Related Root Traits in Lentil: Steps Ahead for the Development of Efficient Screening Protocol for Climate Change Resilient Varieties. In Proceedings of the International Conference Climate Resilient Agriculture: Ways of Adaptation, Rabat, Morocco, 12 December 2019. [Google Scholar]
- Yang, Y.; Saand, M.A.; Huang, L.; Abdelaal, W.B.; Zhang, J.; Wu, Y.; Li, J.; Sirohi, M.H.; Wang, F. Applications of Multi-Omics Technologies for Crop Improvement. Front. Plant Sci. 2021, 12, 1846. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Bohra, A.; Nayyar, H. Advances in “Omics” Approaches to Tackle Drought Stress in Grain Legumes. Plant Breed. 2020, 139, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Taunk, J.; Singh, C.K.; Chaudhary, P.; Gaikwad, K.; Yadav, R.K.; Singh, D.; Pal, M. Comparative RNA Sequencing for Deciphering Nodes of Multiple Abiotic Stress Tolerance in Lentil (Lens culinaris Medikus). Plant Gene 2022, 31, 100373. [Google Scholar] [CrossRef]
- Yi, S.; Zhou, X.; Sun, Z.; Li, X.-P.; Li, W. Cloning and Functional Characterization of a Citrate Synthase Gene of Rhododendron Micranthum Turcz. S. Afr. J. Bot. 2022, 147, 915–925. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Fakhar, A.Z.; Sharif, R.; Chen, H.; Zhang, C.; Ju, L.; Fotopoulos, V.; Siddique, K.H.; Singh, R.K. Smart Reprograming of Plants against Salinity Stress Using Modern Biotechnological Tools. Crit. Rev. Biotechnol. 2022, 15, 1–28. [Google Scholar] [CrossRef]
- Tiwari, M.; Singh, B.; Min, D.; Jagadish, S.K. Omics Path to Increasing Productivity in Less-Studied Crops Under Changing Climate—Lentil a Case Study. Front. Plant Sci. 2022, 13, 813985. [Google Scholar] [CrossRef]
- Malik, J.A.; Mishra, G.; Hajam, Y.A.; Lone, R.; Quazi, S. Metabolome Analyses in Response to Diverse Abiotic Stress. In Omics Approach to Manage Abiotic Stress in Cereals; Springer: Berlin/Heidelberg, Germany, 2022; pp. 103–117. [Google Scholar]
- Razzaq, A.; Sadia, B.; Raza, A.; Khalid Hameed, M.; Saleem, F. Metabolomics: A Way Forward for Crop Improvement. Metabolites 2019, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Raza, A. Metabolomics: A Systems Biology Approach for Enhancing Heat Stress Tolerance in Plants. Plant Cell Rep. 2020, 41, 741–763. [Google Scholar] [CrossRef]
- Foti, C.; Kalampokis, I.F.; Aliferis, K.A.; Pavli, O.I. Metabolic Responses of Two Contrasting Lentil Genotypes to PEG-Induced Drought Stress. Agronomy 2021, 11, 1190. [Google Scholar] [CrossRef]
- Khatib, F.; Makris, A.; Yamaguchi-Shinozaki, K.; Kumar, S.; Sarker, A.; Erskine, W.; Baum, M. Expression of the DREB1A Gene in Lentil (Lens culinaris Medik. Subsp. Culinaris) Transformed with the Agrobacterium System. Crop Pasture Sci. 2011, 62, 488. [Google Scholar] [CrossRef]
- Ashraf, M. Inducing Drought Tolerance in Plants: Recent Advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Baloglu, M.C.; Altunoglu, Y.C.; Baloglu, P.; Yildiz, A.B.; Türkölmez, N.; Çiftçi, Y.Ö. Gene-Editing Technologies and Applications in Legumes: Progress, Evolution, and Future Prospects. Front. Genet. 2022, 13, 859437. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, A.; Gill, R.A.; Hassan, M.U.; Mahmood, A.; Qari, S.; Zaman, Q.U.; Ilyas, M.; Aamer, M.; Batool, M.; Li, H. A Critical Review: Recent Advancements in the Use of CRISPR/Cas9 Technology to Enhance Crops and Alleviate Global Food Crises. Curr. Issues Mol. Biol. 2021, 43, 1950–1976. [Google Scholar] [CrossRef]
- Bhowmik, P.; Konkin, D.; Polowick, P.; Hodgins, C.L.; Subedi, M.; Xiang, D.; Yu, B.; Patterson, N.; Rajagopalan, N.; Babic, V. CRISPR/Cas9 Gene Editing in Legume Crops: Opportunities and Challenges. Legume Sci. 2021, 3, e96. [Google Scholar] [CrossRef]
- Sarker, R.H.; Biswas, A.; Mustafa, B.M.; Mahbub, S.; Hoque, M.I. Agrobacterium-Mediated Transformation of Lentil (Lens culinaris Medik. Plant Tissue Cult. 2003, 13, 1–12. [Google Scholar]
- Sarker, R.H.; Das, S.K.; Hoque, M.I. In Vitro Flowering and Seed Formation in Lentil (Lens culinaris Medik.). Vitro Cell. Dev. Biol.-Plant 2012, 48, 446–452. [Google Scholar] [CrossRef]
- Foti, C.; Pavli, O.I. High-Efficiency Agrobacterium Rhizogenes-Mediated Transgenic Hairy Root Induction of Lens culinaris. Agronomy 2020, 10, 1170. [Google Scholar] [CrossRef]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.-D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D. Speed Breeding Is a Powerful Tool to Accelerate Crop Research and Breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B. Breeding Crops to Feed 10 Billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Mobini, S.H.; Lulsdorf, M.; Warkentin, T.D.; Vandenberg, A. Plant Growth Regulators Improve in Vitro Flowering and Rapid Generation Advancement in Lentil and Faba Bean. Vitro Cell. Dev. Biol.-Plant 2015, 51, 71–79. [Google Scholar] [CrossRef]
- Lulsdorf, M.M.; Banniza, S. Rapid Generation Cycling of an F2 Population Derived from a Cross between Lens culinaris Medik. and Lens Ervoides (Brign.) Grande after Aphanomyces Root Rot Selection. Plant Breed. 2018, 137, 486–491. [Google Scholar] [CrossRef]
- Maglia, F.; Bermejo, C.; Palacios, L.T.; Cointry, E. Speed Breeding Para La Multiplicación de Colecciones Activas En Lenteja (Lens culinaris Medik.). J. Basic Appl. Genet. 2020, 31, 123–145. [Google Scholar]
- Idrissi, O.; Sahri, A.; Udupa, S.; Kumar, S. Single Seed Descent under Extended Photoperiod as a Simple, Rapid and Efficient Breeding Method for Accelerated Genetic Gain in Lentil. In Proceedings of the Third International Legume Society Conference ILS3, Poznań, Polska, 21–24 May 2019. [Google Scholar]
- Idrissi, O. Application of Extended Photoperiod in Lentil: Towards Accelerated Genetic Gain in Breeding for Rapid Improved Variety Development. Moroc. J. Agric. Sci. 2020, 1, 14–19. [Google Scholar]
- Mobini, S.H.; Lulsdorf, M.; Warkentin, T.D.; Vandenberg, A. Low Red: Far-Red Light Ratio Causes Faster in Vitro Flowering in Lentil. Can. J. Plant Sci. 2016, 96, 908–918. [Google Scholar] [CrossRef] [Green Version]
- Bamouh, A. Productivity, Profitability and Farmer’s Adoption Potential of Direct Seeding of Lentils in Zaer Region (Morocco). Moroc. J. Agric. Sci. 2020, 1, 181–185. [Google Scholar]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H. Drought Stress in Grain Legumes during Reproduction and Grain Filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Kassam, A.; Friedrich, T.; Derpsch, R.; Lahmar, R.; Mrabet, R.; Basch, G.; González-Sánchez, E.J.; Serraj, R. Conservation Agriculture in the Dry Mediterranean Climate. Field Crops Res. 2012, 132, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, A.; Farooq, M.; Anees-Ur-Rehman, A.U.; Yadav, R.; Siddique, K. Agronomic Innovations for Enhancing the Yield Potential of Agricultural Crops. Indian J. Agron. 2021, 66, S191–S197. [Google Scholar]
- Saxena, M.C.; Murinda, M.V.; Turk, M.; Trabulsi, N. Productivity and Water-Use of Lentil as Affected by Date of Sowing. Lentil Exp. Lens. 1983, 10, 28–29. [Google Scholar]
- Bejiga, G. Effect of Sowing Date on the Yield of Lentil (Lens culinaris Medik.). J. Agron. Crop Sci. 1991, 167, 135–140. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Loss, S.P.; Pritchard, D.L.; Regan, K.L.; Tennant, D.; Jettner, R.L.; Wilkinson, D. Adaptation of Lentil (Lens culinaris Medik.) to Mediterranean-Type Environments: Effect of Time of Sowing on Growth, Yield, and Water Use. Aust. J. Agric. Res. 1998, 49, 613–626. [Google Scholar] [CrossRef]
- Maphosa, L.; Anwar, M.R.; Luckett, D.J.; Ip, R.H.; Chauhan, Y.S.; Richards, M.F. Impact of Sowing Time and Genotype on Water Use Efficiency of Lentil (Lens culinaris Medick.). Agronomy 2022, 12, 1542. [Google Scholar] [CrossRef]
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Díaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H. Phytohormones Trigger Drought Tolerance in Crop Plants: Outlook and Future Perspectives. Front. Plant Sci. 2022, 12, 799318. [Google Scholar] [CrossRef] [PubMed]
- Jogawat, A.; Yadav, B.; Lakra, N.; Singh, A.K.; Narayan, O.P. Crosstalk between Phytohormones and Secondary Metabolites in the Drought Stress Tolerance of Crop Plants: A Review. Physiol. Plant. 2021, 172, 1106–1132. [Google Scholar] [CrossRef]
- Tavanti, T.R.; de Melo, A.A.R.; Moreira, L.D.K.; Sanchez, D.E.J.; dos Santos Silva, R.; da Silva, R.M.; Dos Reis, A.R. Micronutrient Fertilization Enhances ROS Scavenging System for Alleviation of Abiotic Stresses in Plants. Plant Physiol. Biochem. 2021, 160, 386–396. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gupta, D. Silicon Modulates Nitro-Oxidative Homeostasis along with the Antioxidant Metabolism to Promote Drought Stress Tolerance in Lentil Plants. Physiol. Plant. 2021, 172, 1382–1398. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gonzalez Viejo, C.; Torrico, D.D.; Inayat, S.; Gupta, D. Silicon Supplementation Improves the Nutritional and Sensory Characteristics of Lentil Seeds Obtained from Drought-stressed Plants. J. Sci. Food Agric. 2021, 101, 1454–1466. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Bhardwaj, A.; Bhandari, K.; Jha, U.; Vara Prasad, P.V.; Singh, S.; Kumar, S.; Siddique, K.H.; Nayyar, H. Selenium Supplementation to Lentil (Lens culinaris Medik.) under Combined Heat and Drought Stress Improves Photosynthetic Ability, Antioxidant Systems, Reproductive Function and Yield Traits. Plant Soil 2022, 1–17. [Google Scholar] [CrossRef]
- Amirnia, R.; Ghiyasi, M.; Siavash Moghaddam, S.; Rahimi, A.; Damalas, C.A.; Heydarzadeh, S. Nitrogen-Fixing Soil Bacteria Plus Mycorrhizal Fungi Improve Seed Yield and Quality Traits of Lentil (Lens culinaris Medik). J. Soil Sci. Plant Nutr. 2019, 19, 592–602. [Google Scholar] [CrossRef]
- Jorge, G.L.; Kisiala, A.; Morrison, E.; Aoki, M.; Nogueira, A.P.O.; Emery, R.N. Endosymbiotic Methylobacterium Oryzae Mitigates the Impact of Limited Water Availability in Lentil (Lens culinaris Medik.) by Increasing Plant Cytokinin Levels. Environ. Exp. Bot. 2019, 162, 525–540. [Google Scholar] [CrossRef]
- Singh, O.S.; Singh, R.S. Effects of Phosphorus and Glomus Fasciculatus Inoculation on Nitrogen Fixation, P Uptake and Yield of Lentil (Lens culinaris Medic) Grown on an Unsterilized Sandy Soil. Environ. Exp. Bot. 1986, 26, 185–190. [Google Scholar] [CrossRef]
- Taha, K.; El Attar, I.; Hnini, M.; Raif, A.; Béna, G.; Aurag, J. Beneficial Effect of Rhizobium Laguerreae Co-Inoculated with Native Bacillus Sp. and Enterobacter Aerogenes on Lentil Growth under Drought Stress. Rhizosphere 2022, 22, 100523. [Google Scholar] [CrossRef]
- Farooq, M.; Usman, M.; Nadeem, F.; ur Rehman, H.; Wahid, A.; Basra, S.M.; Siddique, K.H. Seed Priming in Field Crops: Potential Benefits, Adoption and Challenges. Crop Pasture Sci. 2019, 70, 731–771. [Google Scholar] [CrossRef]
- Farooq, M.; Romdhane, L.; Al Sulti, M.K.; Rehman, A.; Al-Busaidi, W.M.; Lee, D.-J. Morphological, Physiological and Biochemical Aspects of Osmopriming-Induced Drought Tolerance in Lentil. J. Agron. Crop Sci. 2020, 206, 176–186. [Google Scholar] [CrossRef]
- GHASSEMI-GOLEZANI, K.; Aliloo, A.A.; Valizadeh, M.; MOGHADDAM, M. Effects of Hydro and Osmo-Priming on Seed Germination and Field Emergence of Lentil (Lens culinaris Medik.). Not. Bot. Horti Agrobot. Cluj-Napoca 2008, 36, 29–33. [Google Scholar]
- Sağlam, S.; Day, S.; Kaya, G.; Gürbüz, A. Hydropriming Increases Germination of Lentil (Lens culinaris Medik.) under Water Stress. Not. Sci. Biol. 2010, 2, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Mrabet, R.; Moussadek, R.; Fadlaoui, A.; Van Ranst, E. Conservation Agriculture in Dry Areas of Morocco. Field Crops Res. 2012, 132, 84–94. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.K.; Halder, S.; Mondal, K.; Singh, K.C.; Nandi, R.; Ghosh, P.K. Response of Lentil (Lens Culinaries) to Post-Rice Residual Soil Moisture under Contrasting Tillage Practices. Agric. Res. 2018, 7, 463–479. [Google Scholar] [CrossRef]
- Das, A.; Layek, J.; Ramkrushna, G.I.; Rangappa, K.; Lal, R.; Ghosh, P.K.; Choudhury, B.U.; Mandal, S.; Ngangom, B.; Dey, U. Effects of Tillage and Rice Residue Management Practices on Lentil Root Architecture, Productivity and Soil Properties in India’s Lower Himalayas. Soil Tillage Res. 2019, 194, 104313. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Accession | References |
|---|---|
| ILL 6002 | [9] |
| JL 1, IPL 98/193, DPL 53 | [77] |
| ILL 123613, ILL 123466 | [74] |
| ILL 7835, ILL 7835, ILL 6075, ILL 7814, ILL 7804, ILL 7833, ILL 8029, ILL 6338, ILL 6104, ILL 7814, ILL 6362 | [14] |
| ILL-10700, ILL-10823, FLIP-96-51 | [81] |
| BM-1247, BM-1227, BM-981, BM-502 | [31] |
| IC559713, IC559696, IC560051, IC560246, IC559647, IC560032, IC559769, IC559757, IC559744, IC835822, IC560337 | [78] |
| Digger, Cumra, Indianhead, ILL5588, ILL6002, ILL5582 | [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeroual, A.; Baidani, A.; Idrissi, O. Drought Stress in Lentil (Lens culinaris, Medik) and Approaches for Its Management. Horticulturae 2023, 9, 1. https://doi.org/10.3390/horticulturae9010001
Zeroual A, Baidani A, Idrissi O. Drought Stress in Lentil (Lens culinaris, Medik) and Approaches for Its Management. Horticulturae. 2023; 9(1):1. https://doi.org/10.3390/horticulturae9010001
Chicago/Turabian StyleZeroual, Abdelmonim, Aziz Baidani, and Omar Idrissi. 2023. "Drought Stress in Lentil (Lens culinaris, Medik) and Approaches for Its Management" Horticulturae 9, no. 1: 1. https://doi.org/10.3390/horticulturae9010001