
An Academic and Technical Overview on Plant Micropropagation Challenges

 ,
,  ,
,  ,
,  and
and
Abstract
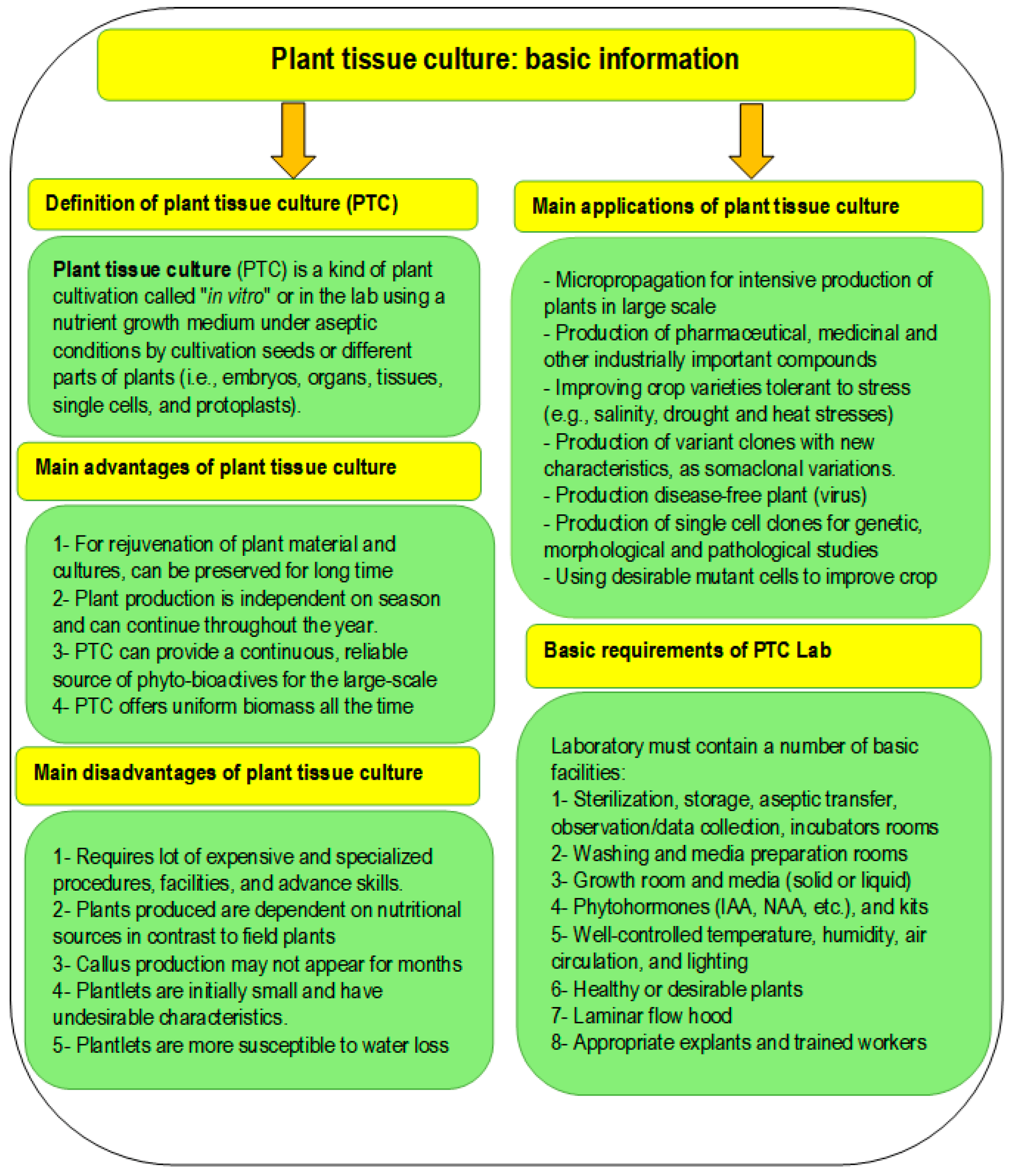
:1. Introduction
2. Methodology of the Review
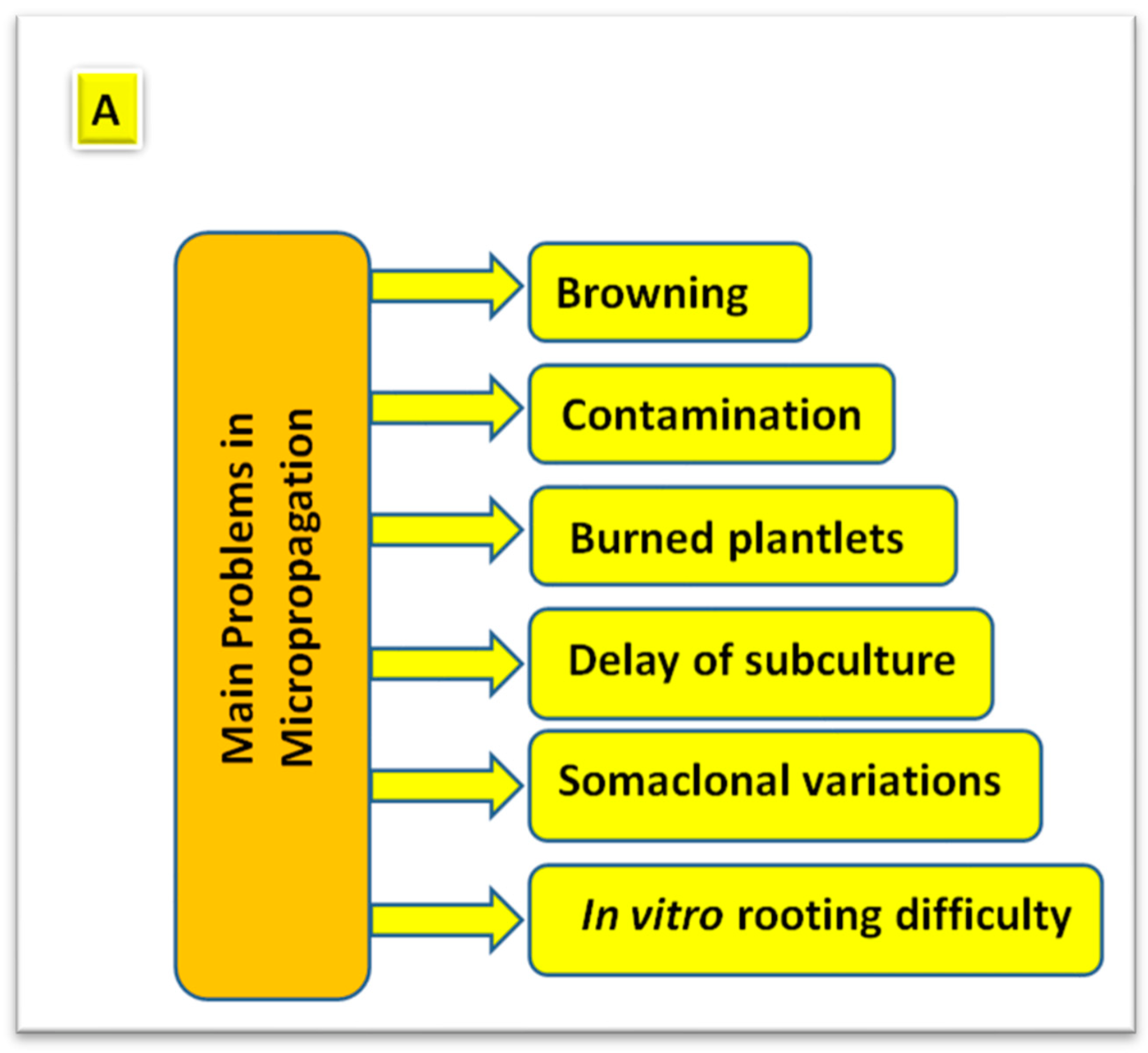
3. Problems Associated with Plant Micropropagation
3.1. Problems Originated Due to Technical Reasons
3.1.1. Contamination of Plant-Tissue Cultures

3.1.2. Delay of Subculture and Burned Plantlets
3.2. Problems Originated Due to Physiological Reasons
3.2.1. Browning of Plant-Tissue Cultures
3.2.2. In Vitro Rooting Difficulty and Failure of Subsequent Acclimatization
3.3. Problems Originated Due to Genetical Reasons
Somaclonal Variation
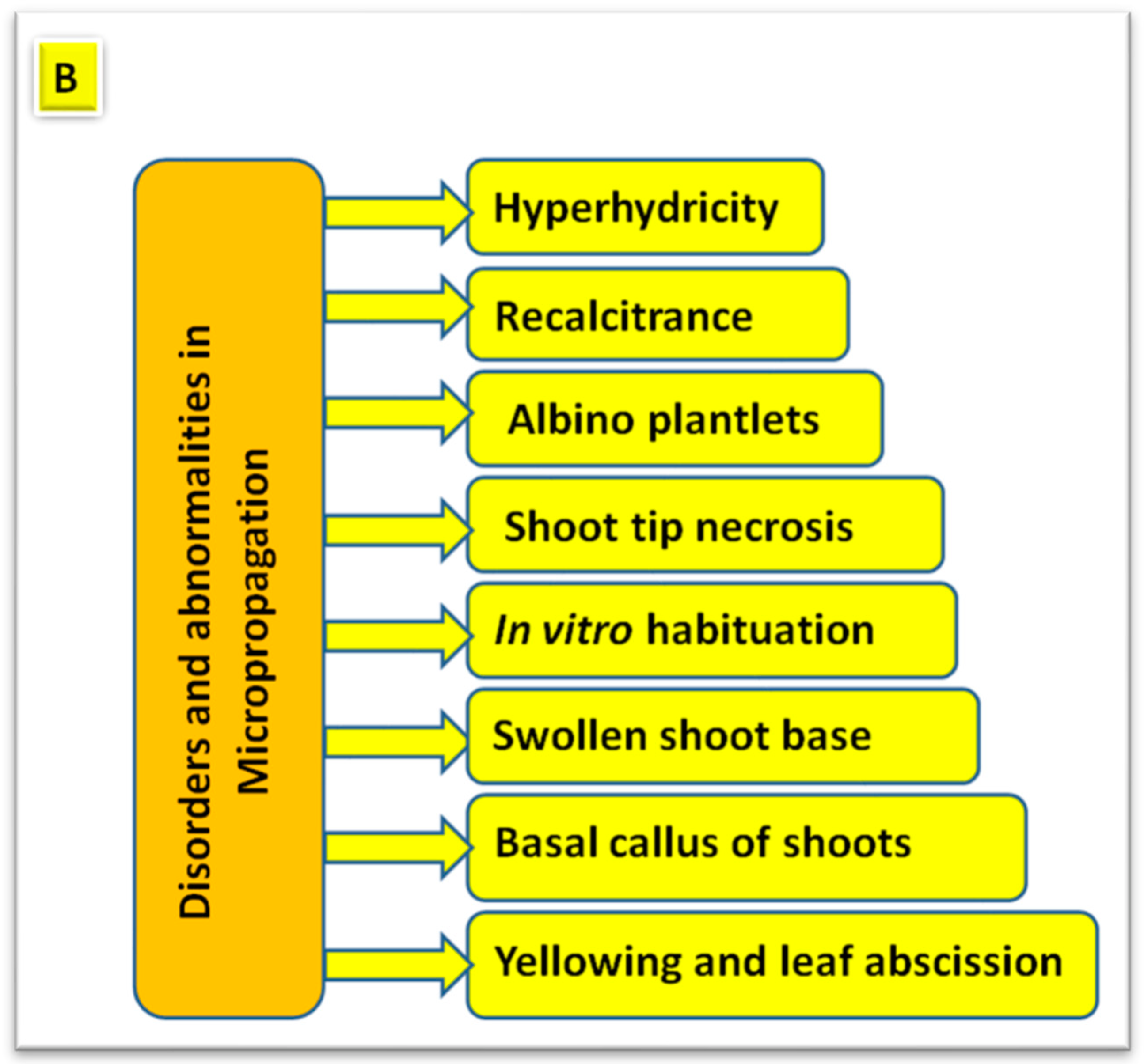
4. Disorders and Abnormalities in Plant Micropropagation
4.1. Hyperhydricity
4.2. Shoot Tip Necrosis
4.3. Albino Plantlets (Albinism) in Anther Cultures
4.4. Recalcitrance in Clonal Micropropagation
4.5. Shoot Abnormalities
4.6. In Vitro Habituation
5. General Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bridgen, M.; Van Houtven, W.; Eeckhaut, T. Plant Tissue Culture Techniques for Breeding. In Ornamental Crops, Handbook of Plant Breeding; Van Huylenbroeck, J., Ed.; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 127–144. [Google Scholar]
- Neumann, K.-H.; Kumar, A.; Imani, J. Plant Cell and Tissue Culture—A Tool in Biotechnology Basics and Application, 2nd ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 1–11. [Google Scholar]
- Viana, C.M.; Freire, D.; Abrantes, P.; Rocha, J.; Pereira, P. Agricultural land systems importance for supporting food security and sustainable development goals: A systematic review. Sci. Total Environ. 2022, 806, 150718. [Google Scholar] [CrossRef] [PubMed]
- Bidabadi, S.S.; Jain, S.M. Cellular, Molecular, and Physiological Aspects of In Vitro Plant Regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Rani, A.; Donovan, N.; Mantria, N. Review: The future of plant pathogen diagnostics in a nursery production system. Biosens. Bioelectron. 2019, 145, 111631. [Google Scholar] [CrossRef] [PubMed]
- Pe, P.P.W.; Naing, A.H.; Soe, M.T.; Kang, H.; Park, K.I.; Kim, C.K. Establishment of meristem culture for virus-free and genetically stable production of the endangered plant Hosta capitata. Sci. Hortic. 2020, 272, 109591. [Google Scholar] [CrossRef]
- Kumar, M.K.; Sandeep, B.V.; Rao, S.P. Development of salt tolerant callus cultures by somatic hybridization between Oryza sativa and mangrove grass Myriostachya wightiana. Ann. Agrar. Sci. 2018, 16, 396–404. [Google Scholar] [CrossRef]
- Wang, M.; Ji, Y.; Feng, S.; Liu, C.; Xiao, Z.; Wang, X.; Wang, Y.; Xia, G. The non-random patterns of genetic variation induced by asymmetric somatic hybridization in wheat. BMC Plant Biol. 2018, 18, 244. [Google Scholar] [CrossRef] [PubMed]
- Park, S. Plant Tissue Culture Techniques and Experiments, 5th ed.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: London, UK, 2021; pp. 1–23. [Google Scholar]
- Naing, A.H.; Adedeji, O.S.; Kim, C.K. Protoplast technology in ornamental plants: Current progress and potential applications on genetic improvement. Sci. Hortic. 2021, 283, 110043. [Google Scholar] [CrossRef]
- Sánchez, M.A.; Coronado, Y.M.; Coronado, A.C.M. Androgenic studies in the production of haploids and doubled haploids in Capsicum spp. Rev. Fac. Nac. Agron. Medellín 2020, 73, 9047–9056. [Google Scholar] [CrossRef]
- Dhiman, K.M.; Sharma, L.; Singh, A.; Sharma, M.M. Ex situ Conservation Using In vitro Methods of an Endangered Plant Sterculia urens Roxb.: A High Volume Trade Plant for Gum. Ind. Crops Prod. 2020, 158, 113015. [Google Scholar] [CrossRef]
- Karakas, F.P.; Bozat, B.G. Fluctuation in secondary metabolite production and antioxidant defense enzymes in in vitro callus cultures of goat’s rue (Galega officinalis) under different abiotic stress treatments. Plant Cell Tissue Organ Cult. 2020, 142, 401–414. [Google Scholar] [CrossRef]
- Soleimani, S.H.; Bernard, F.; Amini, M.; Khavari-Nezhad, R.A. Cadmium accumulation and alkaloid production of Narcissus tazetta plants grown under in vitro condition with cadmium stress. Plant Physiol. Rep. 2020, 25, 51–57. [Google Scholar] [CrossRef]
- Fortini, E.A.; Batista, D.S.; Mamedes-Rodrigues, T.C.; Felipe, S.H.S.; Correia, L.N.F.; Chagas, K.; Silva, P.O.; Rocha, D.I.; Otoni, W.C. Gas exchange rates and sucrose concentrations affect plant growth and production of favonoids in Vernonia condensata grown in vitro. Plant Cell Tissue Organ Cult. 2021, 144, 593–605. [Google Scholar] [CrossRef]
- Kyzioł, A.; Łukasiewicz, S.; Sebastian, V.; Kustrowski, P.; Kozieł, M.; Majda, D.; Cierniak, A. Towards plant-mediated chemistry—Au nanoparticles obtained using aqueous extract of Rosa damascena and their biological activity in vitro. J. Inorg. Biochem. 2021, 214, 111300. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, N.; Herbohn, J.; Harrison, S. Small-scale Forestry Development in Leyte, Philippines: The Central Role of Nurseries. Small-Scale For. 2004, 3, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Degrande, A.; Tadjo, P.; Takoutsing, B.; Asaah, E.; Tsobeng, A.; Zac Tchoundjeu, Z. Getting Trees In to Farmers’ Fields: Success of Rural Nurseries in Distributing High Quality Planting Material in Cameroon. Small-Scale For. 2013, 12, 403–420. [Google Scholar] [CrossRef]
- Marshall, M.; Sutherland, R.; Hulme, P.E. Assessing the role of plant trade networks in the vulnerability of forest nurseries to plant pathogens. Australas. Plant Pathol. 2021, 50, 671–681. [Google Scholar] [CrossRef]
- Seliem, M.K.; Abdalla, N.A.; El-Ramady, H.R. Response of Phalaenopsis Orchid to Selenium and Bio-NanoSelenium: In Vitro Rooting and Acclimatization. Env. Biodiv. Soil Secur. 2020, 4, 277–290. [Google Scholar] [CrossRef]
- Gaspar, T. Vitrification in Micropropagation. In High-Tech and Micropropagation I; Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1991; Volume 17, pp. 116–126. [Google Scholar]
- Pandey, S.; Sundararajan, S.; Ramalingam, S.; Pant, B. Effects of sodium nitroprusside and growth regulators on callus, multiple shoot induction and tissue browning in commercially important Valeriana jatamansi Jones. Plant Cell Tissue Organ Cult. 2020, 142, 653–660. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Nezami-Alanagh, E.; Barreal, M.E.; Kher, M.M.; Wicaksono, A.; Gulyás, A.; Hidvégi, N.; Magyar-Tábori, K.; Mendler-Drienyovszki, N.; Márton, L.; et al. Shoot tip necrosis of in vitro plant cultures: A reappraisal of possible causes and solutions. Planta 2020, 252, 47. [Google Scholar] [CrossRef]
- Pawełkowicz, M.E.; Skarzyńska, A.; Mróz, T.; Bystrzycki, E.; Pląder, W. Molecular insight into somaclonal variation phenomena from transcriptome profiling of cucumber (Cucumis sativus L.) lines. Plant Cell Tissue Organ Cult. 2021, 145, 239–259. [Google Scholar] [CrossRef]
- Ranghoo-Sanmukhiya, V.M. Somaclonal Variation and Methods Used for Its Detection. In Propagation and Genetic Manipulation of Plants; Siddique, I., Ed.; Springer Nature: Singapore, 2021; pp. 1–18. [Google Scholar]
- Thakur, M.; Rakshandha; Sharma, V.; Chauhan, A. Genetic fidelity assessment of long term in vitro shoot cultures and regenerated plants in Japanese plum cvs Santa Rosa and Frontier through RAPD, ISSR and SCoT markers. S. Afr. J. Bot. 2020, 140, 428–433. [Google Scholar] [CrossRef]
- FAO/IAEA. Low-Cost Options for Tissue Culture Technology in Developing Countries. In Proceedings of the Technical Meeting Organized by the Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, Vienna, Austria, 26–30 August 2002; Available online: https://www-pub.iaea.org/mtcd/publications/pdf/te_1384_web.pdf (accessed on 14 June 2022).
- Loyola-Vargas, V.M.; Ochoa-Alejo, N. An Introduction to Plant Tissue Culture: Advances and Perspectives. In Plant Cell Culture Protocols, Methods in Molecular Biology; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Springer Nature: New York, NY, USA, 2018; Volume 1815, pp. 3–13. [Google Scholar]
- Singh, C.R. Review on problems and its remedy in plant tissue culture. Asian J. Biol. Sci. 2018, 11, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Gangopadhyay, M.; Nandi, S.; Roy, S.K.B. An efficient ex plant sterilization protocol for reducing microbial contamination of Solanum tuberosum CV. ‘Kufri jyoti for establishing micropropagation in rainy season. J. Basic Appl. Plant Sci. 2017, 1, 25. [Google Scholar]
- Jugreet, B.S.; Mahomoodally, M.F. Essential oils from 9 exotic and endemic medicinal plants from Mauritius shows in vitro antibacterial and antibiotic potentiating activities. S. Afr. J. Bot. 2020, 132, 355–362. [Google Scholar] [CrossRef]
- Bhatia, S.; Sharma, K.; Dahiya, R.; Bera, T. Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Academic Press: Cambridge, MA, USA; Elsevier: London, UK, 2015; pp. 31–107. [Google Scholar]
- Taha, H.; Ghazy, U.M.; Gabr, A.M.M.; EL-Kazzaz, A.A.A.; Ahmed, E.M.M.; Haggag, K.M. Optimization of in vitro culture conditions affecting propagation of mulberry plant. Bull. Natl. Res. Cent. 2020, 44, 60. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.D.; Ortega, M.; Orellana, M.; Rodríguez, M.; Folta, K.M.; Torres, M.L. In vitro propagation of the Amazonian medicinal plant guayusa (Ilex guayusa) and effects of light in the growth and development of this shade tolerant plant. Plant Cell Tissue Organ Cult. 2021, 147, 503–517. [Google Scholar] [CrossRef]
- Molinari, L.V.; Souza, D.M.S.C.; Avelar, M.L.M.; Fernandes, S.B.; Gonçalves, D.S.; Faria, J.C.T.; de Carvalho, D.; Brondani, G.E. Effects of chemical sterilization of the culture media, porous membranes and luminosity on in vitro culture of Eucalyptus grandis × Eucalyptus urophylla. J. For. Res. 2021, 32, 1587–1598. [Google Scholar] [CrossRef]
- Andújar, I.; González, N.; García-Ramos, J.C.; Bogdanchikova, N.; Pestryakov, A.; Escalona, M.; Concepción, O. Argovit™ silver nanoparticles reduce contamination levels and improve morphological growth in the in vitro culture of Psidium friedrichsthalianum (O. Berg) Nied. SN Appl. Sci. 2020, 2, 2110. [Google Scholar] [CrossRef]
- Nartop, P. Green Sterilization of Rosmarinus officinalis L. Stem Surfaces with Silver Nanoparticles Synthesized Using Rubia tinctorum L. Cell Culture Extracts. Iran J. Sci. Technol. Trans. A Sci. 2018, 42, 411–414. [Google Scholar] [CrossRef]
- Ahmed, N.; Mohamed, H.F.; Xu, C.; Lin, X.; Huang, L. A novel surface sterilization method using Artemisia dracunculus extract for tissue culturing of endangered species Sargassum fusiforme. Plant Cell Tissue Organ Cult. 2022, 149, 135–145. [Google Scholar] [CrossRef]
- Ahmadpoor, F.; Zare, N.; Asghari, R.; Sheikhzadeh, P. Sterilization protocols and the effect of plant growth regulators on callus induction and secondary metabolites production in in vitro cultures Melia azedarach L. AMB Express 2022, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Urtiga, C.D.C.; Silva-Cardoso, I.M.D.A.; Figueiredo, S.A. Low sodium isocyanurate concentrations as a substitute to medium autoclaving in plant tissue culture. Plant Cell Tissue Organ Cult. 2019, 139, 601–604. [Google Scholar] [CrossRef]
- Rency, A.S.; Pandian, S.; Ramesh, M. Influence of adenine sulphate on multiple shoot induction in Clitoria ternatea L. and analysis of phyto-compounds in in vitro grown plants. Biocatal. Agric. Biotechnol. 2018, 16, 181–191. [Google Scholar] [CrossRef]
- Kendon, J.P.; Rajaovelona, L.; Sandford, H.; Fang, R.; Bell, J.; Sarasan, V. Collecting near mature and immature orchid seeds for ex situ conservation: ‘in vitro collecting’ as a case study. Bot. Stud. 2017, 58, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Banna, A.N.; El-Mahrouk, M.E.; Dewir, Y.H.; Farid, M.A.; Abou Elyazid, D.M.; Schumacher, H.M. Endophytic Bacteria in Banana In Vitro Cultures: Molecular Identification, Antibiotic Susceptibility, and Plant Survival. Horticulturae 2021, 7, 526. [Google Scholar] [CrossRef]
- Seliem, M.K.; El-Mahrouk, M.E.; El-Banna, A.N.; Hafez, Y.M.; Dewir, Y.H. Micropropagation of Philodendron selloum: Influence of copper sulfate on endophytic bacterial contamination, antioxidant enzyme activity, electrolyte leakage, and plant survival. S. Afr. J. Bot. 2021, 139, 230–240. [Google Scholar] [CrossRef]
- Abdalla, N.A.; Ragab, M.E.; El-Miniawy, S.M.; Arafa, N.M.; Taha, H.S. A New Aspect for In vitro Propagation of Jerusalem Artichoke and Molecular Assessment Using RAPD, ISSR and SCoT Marker Techniques. Egypt. J. Bot. 2021, 61, 203–218. [Google Scholar] [CrossRef]
- Abou Elyazid, D.M.A.; Salama, A.-M.; Zanaty, A.M.E.; Abdalla, N. In Vitro Propagation and Acclimatization of Banana Plants: Antioxidant Enzymes, Chemical Assessments and Genetic Stability of Regenerates as a Response to Copper Sulphate. Plants 2021, 10, 1853. [Google Scholar] [CrossRef]
- Ray, S.S.; Ali, N. Biotic Contamination and Possible Ways of Sterilization: A Review with Reference to Bamboo Micropropagation. Braz. Arch. Biol. Technol. 2016, 59, e160485. [Google Scholar] [CrossRef]
- Alagarsamy, K.; Shamala, L.F.; Wei, S. Influence of media supplements on inhibition of oxidative browning and bacterial endophytes of Camellia sinensis var. sinensis. 3 Biotech 2018, 8, 356. [Google Scholar] [CrossRef]
- Neri, D.; Cioccolanti, T.; Zuccherelli, G.; Navacchi, O.; Giorgi, V.; Lodolini, E.M. Micropropagation Effects on Juvenile Traits, Flower Differentiation, and Tree Architecture in Young Olive Trees. Agronomy 2020, 10, 1742. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, S.H.; Chung, M.Y.; Park, S.K.; Kim, C.K. In vitro propagation method for production of morphologically and genetically stable plants of different strawberry cultivars. Plant Methods 2019, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, E.V.; Kamalapriya, M.; Bansal, S.; Meena, D.P.S.; Agrawal, A. Improved protocol for micropropagation of genetically uniform plants of commercially important cardamom (Elettaria cardamomum Maton). In Vitro Cell. Dev. Biol. Plant. 2021, 57, 409–417. [Google Scholar] [CrossRef]
- Sirikonda, A.; Jogam, P.; Ellendula, R.; Kudikala, H.; Mood, K.; Allini, V.R. In vitro micropropagation and genetic fidelity assessment in Flemingia macrophylla (Willd.) Merr: An ethnomedicinal plant. Vegetos 2020, 33, 286–295. [Google Scholar] [CrossRef]
- El-Mahrouk, M.E.; El-Shereif, A.R.; Hafez, Y.M.; Abdelaal, K.A.; El-Hendawy, A.S.; Migdadi, H.; Al-Obeed, R.S. Micropropagation of Banana: Reversion, Rooting, and Acclimatization of Hyperhydric Shoots. HortScience 2019, 54, 1384–1390. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, E.G.; Batista, T.R.; Stein, V.C.; Balieiro, F.P.; de Abreu, R.J.; Pires, M.F.; de Souza, P.A.; Paiva, L.V. In vitro serial subculture to improve rooting of Eucalyptus urophylla. New Forest 2020, 51, 801–816. [Google Scholar] [CrossRef]
- Siddique, I.; Javed, S.B.; Al-Othman, M.R.; Anis, M. Stimulation of in vitro organogenesis from epicotyl explants and successive micropropagation round in Cassia angustifolia Vahl.: An important source of sennosides. Agrofor. Syst. 2013, 87, 583–590. [Google Scholar] [CrossRef]
- Khamushi, M.; Ardakani, M.D.; Zarei, A.; Aliabad, K.K. An efficient protocol for micropropagation of old cypress of Abarkuh (Cupressus sempervirens var. horizontalis [Mill.]) under in vitro condition. Plant Cell Tissue Organ Cult. 2019, 138, 597–601. [Google Scholar] [CrossRef]
- Vujović, T.; Ružić, D.J.; Cerović, R. In vitro shoot multiplication as influenced by repeated subculturing of shoots of contemporary fruit rootstocks. Hort. Sci. 2012, 39, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Norton, M.E.; Norton, C.R. Change in shoot proliferation with repeated in vitro subculture of shoots of woody species of Rosaceae. Plant Cell Tissue Organ Cult. 1986, 5, 187–197. [Google Scholar] [CrossRef]
- Remphrey, W.R.; Palmer, C.E.; Blouw, M.J. In vitro branching in relation to repeated subculture in two cultivars of Potentilla fruticosa. Plant Cell Tissue Organ Cult. 1993, 32, 235–240. [Google Scholar] [CrossRef]
- Hamad, A.M.; Taha, R.M. Effect of sequential subcultures on in vitro proliferation capacity and shoot formation pattern of pineapple (Ananas comosus L. Merr.) over different incubation periods. Sci. Hortic. 2008, 117, 329–334. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, T.-T.; Zhang, J. Effect of subculture times on genetic fidelity, endogenous hormone level and pharmaceutical potential of Tetrastigma hemsleyanum callus. Plant Cell Tissue Organ Cult. 2015, 122, 67–77. [Google Scholar] [CrossRef]
- Hassanein, A.M.; Salem, J.M.; Faheed, F.A.; El-nagish, A. Effect of anti-ethylene compounds on isoenzyme patterns and genome stability during long term culture of Moringa oleifera. Plant Cell Tissue Organ Cult. 2018, 132, 201–212. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, H.; Liu, K.; Li, L.; Yang, J.; An, X.; Li, P.; Yun, L.; Zhang, Z. The role of JrPPOs in the browning of walnut explants. Plant Biol. 2021, 21, 9. [Google Scholar] [CrossRef]
- Dobránszki, J.; Jámbor-Benczúr, E.; Hudák, I.; Magyar-Tábori, K. Model experiments for establishment of in vitro culture by micrografting in apple. Int. J. Hortic. Sci. 2005, 1, 47–49. [Google Scholar] [CrossRef]
- Dobránszki, J.; Teixeira da Silva, J.A. Micropropagation of apple—A review. Biotechnol. Adv. 2010, 28, 462–488. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Guly, A.; Magyar-Tabori, K.; Wang, M.R.; Wang, Q.-W.; Dobránszki, J. In vitro tissue culture of apple and other Malus species: Recent advances and applications. Planta 2019, 249, 975–1006. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Gao, C.; Miao, Y.; Liu, Z.; Cui, K. Development of a highly efficient callus induction and plant regeneration system for Dendrocalamus sinicus using hypocotyls as explants. Plant Cell Tissue Organ Cult. 2021, 145, 117–125. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Dobránszki, J.; Hudák, I. Effect of cytokinin content of the regeneration media on in vitro rooting ability of adventitious apple shoots. Sci. Hortic. 2011, 129, 910–913. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Murthy, H.N.; Ammar, M.H.; Alghamdi, S.S.; Al-Suhaibani, N.A.; Alsadon, A.A.; Paek, K.Y. In vitro Rooting of Leguminous Plants: Difficulties, Alternatives, and Strategies for Improvement. Hortic. Environ. Biotechnol. 2016, 57, 311–322. [Google Scholar] [CrossRef]
- Shaik, N.M.; Arha, M.; Nookaraju, A.; Gupta, S.K.; Srivastava, S.; Yadav, A.K.; Kulkarni, P.S.; Abhilash, O.U.; Vishwakarma, R.K.; Singh, S.; et al. Improved method of in vitro regeneration in Leucaena leucocephala. Physiol. Mol. Biol. Plants 2009, 15, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayova, E.; Petrova, M.; Nikolova, M.; Dimitrova, L. Effect of Medium Salt Strength on the Micropropagation, Phenolic Content and Antioxidant Activity of Arnica montana L., Threatened Plant Species. Bio. Bulletin. 2016, 2, 6–13. [Google Scholar]
- Rezali, N.I.; Sidik, N.J.; Saleh, A.; Osman, N.I.; Adam, N.A.M. The effects of different strength of MS media in solid and liquid media on in vitro growth of Typhonium flagelliforme. Asian Pac. J. Trop. Biomed. 2017, 7, 151–156. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Aldubai, A.A.; Kher, M.M.; Alsadon, A.A.; El-Hendawy, S.; Al-Suhaibani, N.A. Optimization of media formulation for axillary shoot multiplication of the red-peeled sweet potato (Ipomoea batatas [L.] Lam.) ‘Abees. Chil. J. Agric. Res. 2020, 80, 3–10. [Google Scholar] [CrossRef]
- Dewir, Y.H.; El-Mahrouk, M.E.; Murthy, H.N.; Yoeup, H.P. Micropropagation of Cattleya: Improved in vitro rooting and acclimatization. Hortic. Environ. Biotechnol. 2015, 56, 89–93. [Google Scholar] [CrossRef]
- Nayak, N.R.; Rath, S.P.; Patnaik, S. In vitro propagation of three epiphytic orchids, Cymbidium aloifolium (L.) Sw., Dendrobium aphyllum (Roxb.), Fisch. and Dendrobium moschatum (Buch-Ham) Sw. through thidiazuron-induced high frequency shoot proliferation. Sci. Hortic. 1997, 71, 243–250. [Google Scholar] [CrossRef]
- Alghamdi, S.S.; Dewir, Y.H.; Khan, M.A.; Migdadi, H.; EL-Harty, E.H.; Aldubai, A.A.; Al-Aizari, A.A. Micropropagation and germplasm conservation of four chickpea (Cicer arietinum L.) genotypes. Chil. J. Agric. Res. 2020, 80, 487–495. [Google Scholar] [CrossRef]
- Hasançebi, S.; Kara, N.T.; Çakir, Ö.; Ari, Ş. Micropropagation and root culture of Turkish endemic Astragalus chrysochlorus (Leguminosae). Turk. J. Botany 2011, 35, 7. [Google Scholar] [CrossRef]
- Isutsa, D.K.; Pritts, M.P.; Mudge, K.W. A protocol for rooting and growing apple rootstock microshoots. Fruit Var. J. 1998, 52, 107–116. [Google Scholar]
- Dobránszki, J.; Magyar-Tábori, K.; Jámbor-Benczúr, E.; Lazányi, J.; Bubán, T.; Szalai, J. Influence of aromatic cytokinins on shoot multiplication and their post-effects on rooting of apple cv. Húsvéti rozmaring. Int. J. Hortic. Sci. 2000, 6, 84–87. [Google Scholar]
- Rescalvo-Morales, A.; Monja-Mio, K.; Robert, M.L.; Sánchez-Teyer, L.F. Telomere length in Agave tequilana Weber plants during the in vitro to ex vitro transition. Plant Cell Tissue Organ Cult. 2019, 136, 133–140. [Google Scholar] [CrossRef]
- Dev, R.; Singh, S.K.; Dayal, V.; Kumar, K.; Singh, T. Standardization of in vitro Hardening Strategies for Tissue Cultured Wine Grape (Vitis vinifera L) Genotypes. Int. J. Curr. Microbiol. App. Sci. 2019, 8, 2108–2117. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Hossain, M.M.; Sharma, M.; Dobránszki, J.; Cardoso, J.C.; Songjun, Z. Acclimatization of in Vitro-derived Dendrobium. Hortic. Plant J. 2017, 3, 110–124. [Google Scholar] [CrossRef]
- Shah, S.; Thapa, B.B.; Chanda, K.; Pradhan, S.; Singh, A.; Varma, A.; Thakur, L.S.; Joshi, P.; Pant, B. Piriformospora indica promotes the growth of the in-vitro-raised Cymbidium aloifolium plantlet and their acclimatization. Plant Signal. Behav. 2019, 14, e1596716. [Google Scholar] [CrossRef] [PubMed]
- Cantabella, D.; Dolcet-Sanjuan, R.; Casanovas, M.; Solsona, C.; Torres, R.; Teixido, N. Inoculation of in vitro cultures with rhizosphere microorganisms improve plant development and acclimatization during immature embryo rescue in nectarine and pear breeding programs. Sci. Hortic. 2020, 273, 109643. [Google Scholar] [CrossRef]
- De Araújo, R.C.; Rodrigues, F.A.; Nadal, M.C.; Ribeiro, M.D.; Antonio, C.A.C.; Rodrigues, V.A.; de Souza, A.C.; Pasqual, M.; Doria, J. Acclimatization of Musa spp. seedlings using endophytic Bacillus spp. and Buttiauxella agrestis strains. Microbiol. Res. 2021, 248, 126750. [Google Scholar] [CrossRef]
- Hoang, N.N.; Kitaya, Y.; Shibuya, T.; Endo, R. Effects of supporting materials in in vitro acclimatization stage on ex vitro growth of wasabi plants. Sci. Hortic. 2020, 261, 109042. [Google Scholar] [CrossRef]
- Acquaah, G. Clonal propagation and in vitro culture. In Principles of Plant Genetics and Breeding, 2nd ed.; Acquaah, G., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 146–170. [Google Scholar]
- Jevremović, S.; Subotić, A.; Miljković, D.; Trifunović, M.; Petrić, M.; Cingel, A. Clonal fidelity of chrysanthemum cultivars after long term micropropagation by stem segment culture. VII International symposium on in vitro culture and horticultural breeding. Acta Hortic. 2012, 961, 211–216. [Google Scholar] [CrossRef]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.K.; Lee, Z.H.; Mubbarakh, S.A.; Antony, J.J.J.; Chew, B.L.; Subramaniam, S. Effects of plant growth regulators and activated charcoal on somaclonal variations of protocorm-like bodies (PLBs) of Dendrobium Sabin Blue orchid. Biocatal. Agric. Biotechnol. 2019, 22, 101426. [Google Scholar] [CrossRef]
- Bradaï, F.; Sánchez-Romero, C.; Martín, C. Somaclonal variation in olive (Olea europaea L.) plants regenerated via somatic embryogenesis: Influence of genotype and culture age on genetic stability. Sci. Hortic. 2019, 251, 260–266. [Google Scholar] [CrossRef]
- Mirani, A.A.; Teo, C.H.; Markhand, G.S.; Abul-Soad, A.A.; Harikrishna, J.A. Detection of somaclonal variations in tissue cultured date palm (Phoenix dactylifera L.) using transposable element-based markers. Plant Cell Tissue Organ Cult. 2020, 141, 119–130. [Google Scholar] [CrossRef]
- Dhurve, L.; Kumar, A.; Bhaskar, J.; Sobhana, A.; Francies, R.M.; Mathew, D. Wide variability among the ‘Mauritius’ somaclones demonstrates somaclonal variation as a promising improvement strategy in pineapple (Ananas comosus L.). Plant Cell Tissue Organ Cult. 2021, 145, 701–705. [Google Scholar] [CrossRef]
- Noormohammadi, Z.; Habibollahi, H.; Farahani, F.; Sheidai, M. Molecular analysis of somaclonal variation in Linum usitatissimum and L. album. Plant Gene 2020, 24, 100261. [Google Scholar] [CrossRef]
- Solano, M.C.P.; Ruız, J.S.; Arnao, M.R.; Castro, O.C.; Tovar, M.E.G.; Bello, J.J.B. Evaluation of in vitro shoot multiplication and ISSR marker based assessment of somaclonal variants at different subcultures of vanilla (Vanilla planifolia Jacks). Physiol. Mol. Biol. Plants 2019, 25, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Sreelekshmi, R.; Siril, E.A. Effective reversal of hyperhydricity leading to efficient micropropagation of Dianthus chinensis L. 3 Biotech 2021, 11, 95. [Google Scholar] [CrossRef]
- Gao, H.; Xia, X.; An, L.; Xin, X.; Liang, Y. Reversion of hyperhydricity in pink (Dianthus chinensis L.) plantlets by AgNO3 and its associated mechanism during in vitro culture. Plant Sci. 2017, 254, 1–11. [Google Scholar] [CrossRef]
- Park, S.W.; Jeon, J.H.; Kim, H.S.; Park, Y.M.; Aswath, C.; Joung, H. Effect of sealed and vented gaseous microenvironments on the hyperhydricity of potato shoots in vitro. Sci. Hortic. 2004, 99, 199–205. [Google Scholar] [CrossRef]
- Gómez, C.; Ríos, D.; Sánchez-Olate, M. Effect of successive subculture on adventitious caulogenesis of Eucalyptus globulus. Bosque 2007, 28, 13–17. (In Spanish) [Google Scholar] [CrossRef] [Green Version]
- Bayraktar, M.; Hayta-Smedley, S.; Unal, S.; Varol, N.; Gurel, A. Micropropagation and prevention of hyperhydricity in olive (Olea europaea L.) cultivar ‘Gemlik’. S. Afr. J. Bot. 2020, 128, 264–273. [Google Scholar] [CrossRef]
- Zárate-Salazar, J.R.; de Souza, L.M.; de Morais, M.B.; Neto, L.P.; Willadino, L.; Gouveia-Neto, A.; Ulisses, C. Light-emitting diodes and gas exchange facilitation minimize hyperhydricity in Lippia grata: Physiological, biochemical and morpho anatomical aspects. S. Afr. J. Bot. 2020, 135, 164–171. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A.; Muthukrishnan, S. Role of Biogenic Silver Nanoparticles on Hyperhydricity Reversion in Dianthus chinensis L. an In Vitro Model Culture. J. Plant Growth Regul. 2021, 41, 23–39. [Google Scholar] [CrossRef]
- Liu, M.; Jiang, F.; Kong, X.; Tian, J.; Wu, Z.; Wu, Z. Effects of multiple factors on hyperhydricity of Allium sativum L. Sci. Hortic. 2017, 217, 285–296. [Google Scholar] [CrossRef]
- Franck, T.; Kevers, C.; Gaspar, T.; Dommes, J.; Deby, C.; Greimers, R.; Serteyn, D.; Deby-Dupont, G. Hyperhydricity of Prunus avium shoots cultured on gelrite: A controlled stress response. Plant Physiol. Biochem. 2004, 42, 519–527. [Google Scholar] [CrossRef]
- Al-Mayahi, A.M.W.; Ali, A.H. Effects of different types of gelling agents on in vitro organogenesis and some physicochemical properties of date palm buds, Showathy cv. Folia Oecol. 2021, 48, 110–117. [Google Scholar] [CrossRef]
- Quiala, E.; Jiménez-Tello, M.V.; Barbón, R.; Chávez, M.; de Feria, M.; La, O.M.; Pérez, M. Influence of 6-Benzyladenine and gelling agent on the reduction of hyperhydricity in Tectona grandis L. Rev. Colomb. Biotecnol. 2014, 16, 129. [Google Scholar] [CrossRef]
- Ivanova, M.; van Staden, J. Influence of gelling agent and cytokinins on the control of hyperhydricity in Aloe polyphylla. Plant Cell Tissue Organ Cult. 2011, 104, 13–21. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A. Influence of polyamines on hyperhydricity reversion and its associated mechanism during micropropagation of China pink (Dianthus chinensis L.). Physiol. Mol. Biol. Plants 2020, 26, 2035–2045. [Google Scholar] [CrossRef] [PubMed]
- Pence, V.C.; Finke, L.R.; Niedz, R.P. Evaluating a DOE screen to reduce hyperhydricity in the threatened plant, Cycladenia humilis var jonesii. In Vitro Cell. Dev. Biol. Plant 2020, 56, 215–229. [Google Scholar] [CrossRef]
- Gao, H.; Xu, D.; Zhang, H.; Cheng, X.; Yang, Q. Effects of culture medium composition and PEG on hyperhydricity in Dendrobium officinale. In Vitro Cell. Dev. Biol. Plant 2020, 56, 143–149. [Google Scholar] [CrossRef]
- Surakshitha, N.C.; Soorianathasundaram, K.; Ganga, M.; Raveendran, M. Alleviating shoot tip necrosis during in vitro propagation of grape cv. Red Globe. Sci. Hortic. 2019, 248, 118–125. [Google Scholar] [CrossRef]
- Kabylbekova, B.; Kovalchuk, I.; Mukhitdinova, Z.; Turdiyev, T.; Kairova, G.; Madiyeva, G.; Reed, B.M. Reduced major minerals and increased minor nutrients improve micropropagation in three apple cultivars. In Vitro Cell. Dev. Biol. Plant 2020, 56, 335–349. [Google Scholar] [CrossRef]
- Al-Aizari, A.A.; Al-Obeed, R.S.; Mohamed, M.A.H. Improving micropropagation of some grape cultivars via boron, calcium and phosphate. Electron. J. Biotechnol. 2020, 48, 95–100. [Google Scholar] [CrossRef]
- Nezami-Alanagh, E.; Garoosi, G.A.; Landín, M.; Gallego, P.P. Computer-based tools provide new insight into the key factors that cause physiological disorders of pistachio rootstocks cultured in vitro. Sci. Rep. 2019, 9, 9740. [Google Scholar] [CrossRef] [PubMed]
- Cheong, E.J.; Na, M.; Jeong, U. The effect of endophytic bacteria on in vitro shoot growth of Prunus yedoensis and its identification and elimination. In Vitro Cell. Dev. Biol. Plant 2020, 56, 200–206. [Google Scholar] [CrossRef]
- Thakur, M.; Sharma, V.; Luharch, R. Propagation of plum (Prunus salicina L.) cultivar Frontier in vitro through control of shoot tip necrosis (STN) and validation of genetic integrity using ISSR markers. Plant Physiol. Rep. 2021, 26, 238–246. [Google Scholar] [CrossRef]
- Martin, K.P.; Zhang, C.L.; Slater, A.; Madassery, J. Control of shoot necrosis and plant death during micro-propagation of banana and plantains (Musa spp.). Plant Cell Tissue Organ Cult. 2007, 88, 51–59. [Google Scholar] [CrossRef]
- Naaz, A.; Shahzad, A.; Anis, M. Effect of adenine sulphate interaction on growth and development of shoot regeneration and inhibition of shoot tip necrosis under in vitro condition in adult Syzygium cumini L.—A multipurpose tree. Appl. Biochem. Biotechnol. 2014, 173, 90–102. [Google Scholar] [CrossRef]
- Park, J.S.; Naing, A.H.; Kim, C.K. Effects of ethylene on shoot initiation, leaf yellowing, and shoot tip necrosis in roses. Plant Cell Tissue Organ Cult. 2016, 127, 425–431. [Google Scholar] [CrossRef]
- Kumari, M.; Clarke, H.J.; Small, I.; Siddique, K.H.M. Albinism in Plants: A Major Bottleneck in Wide Hybridization, Androgenesis and Doubled Haploid Culture. CRC Crit. Rev. Plant Sci. 2009, 28, 393–409. [Google Scholar] [CrossRef]
- Isah, T. Changes in the biochemical parameters of albino, hyperhydric and normal green leaves of Caladium bicolor cv. “Bleeding hearts” in vitro long-term cultures. J. Photochem. Photobiol. B Biol. 2019, 191, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Sara, H.C.; René, G.H.; Rosa, U.C.; Angela, K.G.; Clelia, D.P. Agave angustifolia albino plantlets lose stomatal physiology function by changing the development of the stomatal complex due to a molecular disruption. Mol. Genet Genom. 2020, 295, 787–805. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.K.; Wang, K.; Zhang, W.; Liu, H.Y.; Du, L.P.; Hu, H.R.; Ye, X.G. Comprehensive analysis of differently expressed genes and proteins in albino and green plantlets from a wheat anther culture. Biol. Plant. 2017, 61, 255–265. [Google Scholar] [CrossRef]
- Harada, T.; Sato, T.; Asaka, D.; Matsukawa, I. Large-scale deletions of rice plastid DNA in anther culture. Theor. Appl. Genet. 1991, 81, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Yaronskaya, E.; Ziemann, V.; Walter, G.; Averina, N.; Borner, T.; Grimm, B. Metabolic control of the tetrapyrrole biosynthetic pathway for porphyrin distribution in the barley mutant albostrians. Plant J. 2003, 35, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Schmitz-Linneweber, C.; Williams-Carrier, R.E.; Williams-Voelker, P.M.; Kroeger, T.S.; Vichas, A.; Barkan, A. A pentatricopeptide repeat protein facilitates the trans-splicing of the maize chloroplast rps12 pre-mRNA. Plant Cell 2006, 18, 2650–2663. [Google Scholar] [CrossRef] [Green Version]
- Day, A.; Ellis, T.H.N. Deleted forms of plastid DNA in albino plants from cereal anther culture. Curr. Genet. 1985, 9, 671–678. [Google Scholar] [CrossRef]
- Dunford, R.; Walden, R. Plastid genome structure and plastid related transcript levels in albino barley plants derived from anther culture. Curr. Genet. 1991, 20, 339–347. [Google Scholar] [CrossRef]
- Migneault, A.; Sandhu, H.; Per McCord, P.; Zhao, D.; Erickson, J. Albinism in Sugarcane: Significance, Research Gaps, and Potential Future Research Developments. Sugar Tech. 2019, 21, 536–541. [Google Scholar] [CrossRef]
- Canonge, J.; Roby, C.; Hamon, C.; Potin, P.; Pfannschmidt, T.; Philippot, M. Occurrence of albinism during wheat androgenesis is correlated with repression of the key genes required for proper chloroplast biogenesis. Planta 2021, 254, 123. [Google Scholar] [CrossRef] [PubMed]
- Jacquard, C.; Nolin, F.; Hecart, C.; Rashal, D.G.I.; Dhondt-Cordelier, S.; Rajbir, S.; Devaux, S.P.; Mazeyrat-Gourbeyre, F.; Clement, C. Microspore embryogenesis and programmed cell death in barley: Effects of copper on albinism in recalcitrant cultivars. Plant Cell Rep. 2009, 28, 1329–1339. [Google Scholar] [CrossRef]
- Ćalić, D.; Bohanec, B.; Devrnja, N.; Milojević, J.; Tubić, L. Impact of abscisic acid in overcoming the problem of albinism in horse chestnut androgenic embryos. Trees 2013, 27, 755–762. [Google Scholar] [CrossRef]
- Gailis, A.; Samsone, I.; Šēnhofa, S.; Girgžde, E.; Kāpostiņš, R.; Jansons, A. Silver birch (Betula pendula Roth.) culture initiation in vitro and genotype determined differences in micropropagation. New Forests 2021, 52, 791–806. [Google Scholar] [CrossRef]
- Bonga, J.M.; Klimaszewska, K.K.; von Aderkas, P. Recalcitrance in clonal propagation, in particular of conifers. Plant Cell Tissue Organ Cult. 2010, 100, 241–254. [Google Scholar] [CrossRef]
- Bonga, J.M. Can explant choice help resolve recalcitrance problems in in vitro propagation, a problem still acute especially for adult conifers? Trees 2017, 31, 781–789. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A. The role of thin cell layers in regeneration and transformation in orchids. Plant Cell Tissue Organ Cult. 2013, 113, 149–161. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Dobranszki, J. Plant thin cell layers: A 40-year celebration. J. Plant Growth Regul. 2013, 32, 922–943. [Google Scholar] [CrossRef]
- Parthibhan, S.; Rao, M.V.; Teixeira da Silva, J.A.; Kumar, T.S. Somatic embryogenesis from stem thin cell layers of Dendrobium aqueum. Biol. Plant. 2018, 62, 439–450. [Google Scholar] [CrossRef]
- Bairwa, V.K.; Kachhwaha, S.; Kothari, S.L. Phloroglucinol Mediated Shoot Bud Elongation in Capsicum annuum L. Natl. Acad. Sci. Lett. 2012, 35, 331–335. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, S.; Duguid, S.; Dribnenki, P.; Kenaschuk, E. Effect of sucrose concentration on elongation of shoots from flax anther culture. Plant Cell Tissue Organ Cult. 2003, 72, 181–183. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Nurmansyah; Naidoo, Y.; Teixeira da Silva, J.A. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Ha, N.T.M.H.; Do, C.M.; Hoang, T.T.; Ngo, N.D.; Bui, L.V.; Nhut, D.T. The effect of cobalt and silver nanoparticles on overcoming leaf abscission and enhanced growth of rose (Rosa hybrida L. ‘Baby Love’) plantlets cultured in vitro. Plant Cell Tissue Organ Cult. 2020, 141, 393–405. [Google Scholar] [CrossRef]
- Mahmoud, L.M.; Grosser, J.W.; Dutt, M. Silver compounds regulate leaf drop and improve in vitro regeneration from mature tissues of Australian finger lime (Citrus australasica). Plant Cell Tissue Organ Cult. 2020, 141, 455–464. [Google Scholar] [CrossRef]
- Jin, J.; Essemine, J.; Duan, J.; Xie, Q.; Zhu, J.; Cai, W. Regeneration of active endogenous IAA in rice calli following acclimation to 2, 4 D free medium. Plant Growth Regul. 2021, 93, 203–220. [Google Scholar] [CrossRef]
- Pischke, M.S.; Huttlin, E.L.; Hegeman, A.D.; Sussman, M.R. A Transcriptome-Based Characterization of Habituation in Plant Tissue Culture. Plant Physiol. 2006, 140, 1255–1278. [Google Scholar] [CrossRef] [Green Version]
- Meins, F., Jr. Habituation: Heritable variation in the requirement of cultured plant cells for hormones. Annu. Rev. Genet 1989, 23, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Gautheret, G. Hétéro-auxine et cultures de tissues végétaux. Bull. Soc. Chim. Biol. 1942, 24, 13–47. [Google Scholar]
- Jin, J.; Essemine, J.; Duan, J.; Zhu, J.; Cai, W. Rice calli may decelerate its metabolism to adapt hormone free medium. Plant Cell Tissue Organ Cult. 2021, 145, 223–238. [Google Scholar] [CrossRef]
- Wikandari, R.; Manikharda; Baldermann, S.; Ningrum, A.; Taherzadeh, M.J. Application of cell culture technology and genetic engineering for production of future foods and crop improvement to strengthen food security. Bioengineered 2021, 12, 11305–11330. [Google Scholar] [CrossRef]
- Sudheer, W.N.; Thiruvengadam, M.; Nagella, P. A comprehensive review on tissue culture studies and secondary metabolite production in Bacopa monnieri L. Pennell: A nootropic plant. Crit. Rev. Biotechnol. 2022, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.K.; Thakur, A.K.; Kumar, P. Agricultural Biotechnology: Latest Research and Trends; Springer: Singapore, 2021; pp. 1–36. [Google Scholar] [CrossRef]
- Gupta, S.; Chaturvedi, P. Commercial Scale Tissue Culture for Horticulture and Plantation Crops; Springer: Singapore, 2022; pp. 1–21. [Google Scholar] [CrossRef]
- Nhut, D.T.; Tung, H.T.; Yeung, E.C.-T. Plant Tissue Culture: New Techniques and Application in Horticultural Species of Tropical Region; Springer: Singapore, 2022; pp. 1–24. [Google Scholar] [CrossRef]
- Moyo, M.; Bairu, M.W.; Amoo, S.O.; Van Staden, V. Plant biotechnology in South Africa: Micropropagation research endeavours, prospects and challenges. S. Afr. J. Bot. 2011, 77, 996–1011. [Google Scholar] [CrossRef] [Green Version]
- Nithya, V.; Kamalam, M. Standardization of a protocol for micropropagation of Eupatorium glandulosum L. an important medicinal plant. Plant Cell Tissue Organ Cult. 2021, 146, 339–344. [Google Scholar] [CrossRef]
- Rafiq, S.; Rather, Z.A.; Bhat, R.A.; Nazki, I.T.; AL-Harbi, M.S.; Banday, N.; Farooq, I.; Samra, B.N.; Khan, M.H.; Ahmed, A.F.; et al. Standardization of in vitro micropropagation procedure of Oriental Lilium Hybrid Cv. ‘Ravenna’. Saudi J. Biol. Sci. 2021, 28, 7581–7587. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.S.; Rather, Z.A.; Nazki, I.T.; Banday, N.; Wani, T.; Rafiq, S.; Farooq, I.; Noureldeen, A.; Darwish, H. Standardization of in vitro micropropagation of Winter Jasmine (Jasminum nudiflorum) using nodal explants. Saudi J. Biol. Sci. 2022, 29, 3425–3431. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Studied Plants | Surface-Sterilization Methods and Sterilized Items | Main Findings of the Study | Refs. |
|---|---|---|---|
| Potato (Solanum tuberosum L.) | Ultraviolet-C radiation to explants for 10 min | The external contamination of explants has been reduced. | [29] |
| Mulberry (Morus alba L.) | Mercuric chloride at 0.2% for 10 min to shoot tips and auxiliary buds | Minimum percentage of contamination and highly percentage of survival in correlation with shoot development were noticed for all mulberry cultivars under investigation. | [33] |
| Guayusa (Ilex guayusa Loes.) | 70% ethanol for 2 min + 2.5% sodium hypochlorite + five drops of Tween-20 for 25 min to apical stem segment | Surface sterilization with ethanol and sodium hypochlorite resulted in 100% surface-sterilized stakes. | [34] |
| Eucalyptus (Eucalyptus obliqua) | Active chlorine added to culture media at 0.005% for establishment and 0.003% for shoot multiplication and elongation | Active chlorine at 0.005% led to the lowest fungal contamination rate in the establishment stage, where at 0.003% resulted in maximum number of shoots per explant and the greatest shoot length in the multiplication stage. | [35] |
| Guava (Psidium guajava L.) | Silver nanoparticles (AgNPs) at 50 mg L−1 directly to shoot tips for 5 min or at 5 mg L−1 added to culture medium | AgNPs at 50 mg L−1 yielded a contamination rate of 40%, where at 5 mg L−1 reduced shoot contamination rate to 50% compared to controls (80%) and enhanced multiplication rate of the shoots by 180%, as an alternative method for surface sterilization of explants which are easily damaged by commonly used surface sterilizing. | [36] |
| Rosmarinus officinalis L. | Bio-synthesized of silver nanoparticles via Rubia tinctorum L. using cell culture were applied for surface sterilization of stem explants | Sterile explant percentages varied between 40 and 97% and no browning was observed. This method could be used in surface sterilization of explants which have a browning problem caused by their phenolic contents. | [37] |
| Sargassum fusiforme | A crude extract of a medicinal herbal plant Tarragon (Artemisia dracunculus) was used for surface sterilization of explants (leaf, stipe, and stolon) cultured in vitro | The crude extract of A. dracunculus showed a high microbial sterilization effect with (90, 80, and 20%) for leaves, stipes and stolons, respectively. It has very low toxicity to plant tissues compared to chemical sterilants. | [38] |
| Chinaberry (Melia azedarach L.) | Dipping leaf explants in 2 g L−1 benomyl for 2 h + 7% hydrogen peroxide (H2O2) for 10 min + 2% NaOCl for 12 min for surface sterilization | The lowest contamination percentage of explants and browning as well as the highest percentage of callus induction and growth were observed. | [39] |
| Carnation (Dianthus caryophyllus) | Sodium dichloroisocyanurate (NaDCC) was applied as a medium sterilizer to culture medium at 0.02 g L−1 instead of autoclave sterilization | Contamination rate recorded below 5%, sodium isocyanurate has the potential to substitute media autoclaving in plant-tissue culture. | [40] |
| Butterfly pea Clitoria ternatea L. | 0.1% Bavistin solution + 70% ethanol, and 0.1% HgCl2 was used for surface sterilization of nodal explants | Microbial contamination was eliminated and then surface sterilized nodal explants were used for shoot multiplication induction. | [41] |
| Orchid (Angraecum rutenbergianm Kraenzl) | 0.5% (w/v) NaDCC solution + 2 mL L−1 Plant Preservative Mixture (PPM™) was used for surface sterilization of Seed capsules | 87.5% of the total number of capsules was disinfected and the seeds inside them were clean after 3 months of culture. Both NaDCC and PPM were essential to suppress microbial growth. | [42] |
| Applied Substance (Name and Concentration) | Plant Species/Cultivar | Success of Decontamination | Refs. |
|---|---|---|---|
| Kanamycin and streptomycin sulphate at 10 μg ml−1 each were added to shoot multiplication medium +2 mg L−1 BAP +10 mg L−1 adenine sulfate | Guadua angustifolia Kunth | Bacterial growth was inhibited and intensive formation of high-quality shoots was observed. | [48] |
| Antibiotics (timentin at 150 mg L−1 + gentamycin at 30 mg L−1 were added to culture medium | Camellia sinensis var. sinensis | They were effective to eliminate bacterial endophytic up to 24 days with 0% contamination. | [49] |
| Antibiotic, cefotaxime at 62.5 mg L−1 was supplemented to ½ Murashige and Skoog (MS) medium for establishment | Jerusalem artichoke (Helianthus tuberosus L.) | It recorded 0% contamination 100% survival of stem nodes cultures. | [45] |
| Copper sulfate (CuSO4 5H2O) at 60 mg L−1 was added to MS medium +3 mg L−1 BA + 1 mg L−1 KIN for shoot multiplication | Banana (Musa sp.) | The growth of the endophytic bacteria was inhibited by recording 0% contamination. | [46] |
| Copper sulfate (CuSO4 5H2O) at 70 mg L−1 was supplemented to MS medium + 5 mg L−1 BA for shoot multiplication | Philodendron selloum | It eliminated the endogenous bacteria contamination to 0%, without decline in growth of in vitro shoots. | [44] |
| Plant Species/Cultivar | Applied Substance (Name and Concentration) | Success (%) | Refs. |
|---|---|---|---|
| Pink (Dianthus chinensis L.) | AgNO3 at 29.4 µM L−1 supplemented to culture medium | 67% of the hyperhydric Dianthus chinensis L. in vitro plantlets have been revert to normal appearance. | [97] |
| Garlic (Allium sativum L.) | Ascorbic acid at 250 µM + salicylic acid at 50 µM + spermidine at 10 µM + H2O2 at 50 µM | Hyperhydricity was relieved. | [103] |
| Olive (Olea europaea L.) cv. ‘Gemlik’ | Agar-agar at 0.65% w/v supplemented to culture medium | Hyperhydricity was prevented by changing the gelling agent from gelrite to Agar-Agar. | [100] |
| China pink (Dianthus chinensis L.) | Polyamines as spermine at 5 µM supplemented to MS medium | It significantly reduced the hyperhydrated shoots to 0.33% compared to control treatment (100%), and the number of healthy reverted shoots was maximum (11.0). | [108] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, N.; El-Ramady, H.; Seliem, M.K.; El-Mahrouk, M.E.; Taha, N.; Bayoumi, Y.; Shalaby, T.A.; Dobránszki, J. An Academic and Technical Overview on Plant Micropropagation Challenges. Horticulturae 2022, 8, 677. https://doi.org/10.3390/horticulturae8080677
Abdalla N, El-Ramady H, Seliem MK, El-Mahrouk ME, Taha N, Bayoumi Y, Shalaby TA, Dobránszki J. An Academic and Technical Overview on Plant Micropropagation Challenges. Horticulturae. 2022; 8(8):677. https://doi.org/10.3390/horticulturae8080677
Chicago/Turabian StyleAbdalla, Neama, Hassan El-Ramady, Mayada K. Seliem, Mohammed E. El-Mahrouk, Naglaa Taha, Yousry Bayoumi, Tarek A. Shalaby, and Judit Dobránszki. 2022. "An Academic and Technical Overview on Plant Micropropagation Challenges" Horticulturae 8, no. 8: 677. https://doi.org/10.3390/horticulturae8080677