
Breeding New Premium Quality Cultivars by Citrus Breeding 2.0 in Japan: An Integrative Approach Suggested by Genealogy


Abstract
:1. Introduction
2. Citrus Crossbreeding Effort Aims to Improve Fruit Quality
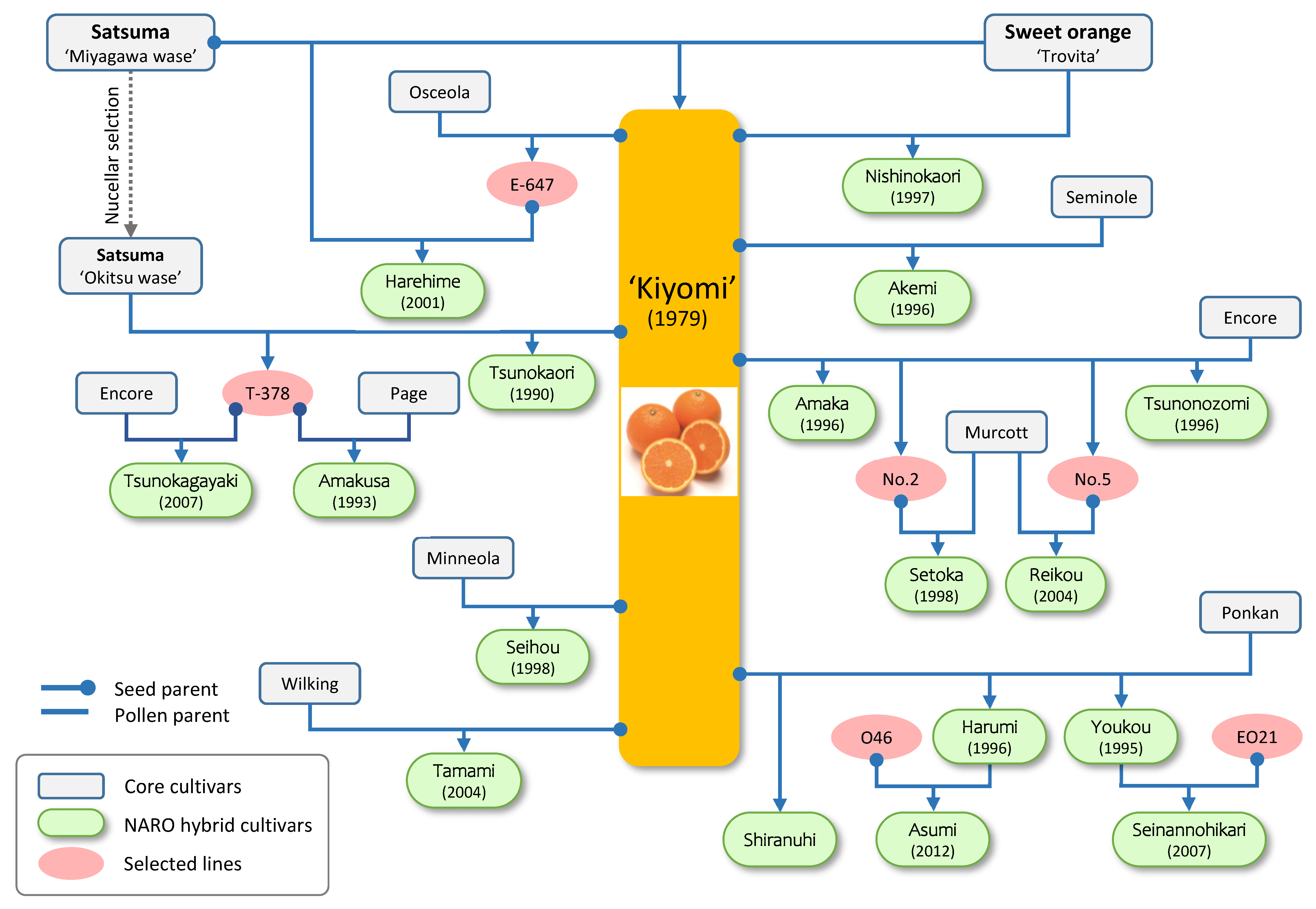
2.1. Past Efforts of the Citrus Breeding Program in NARO
2.2. Constraints of Conventional Crossbreeding
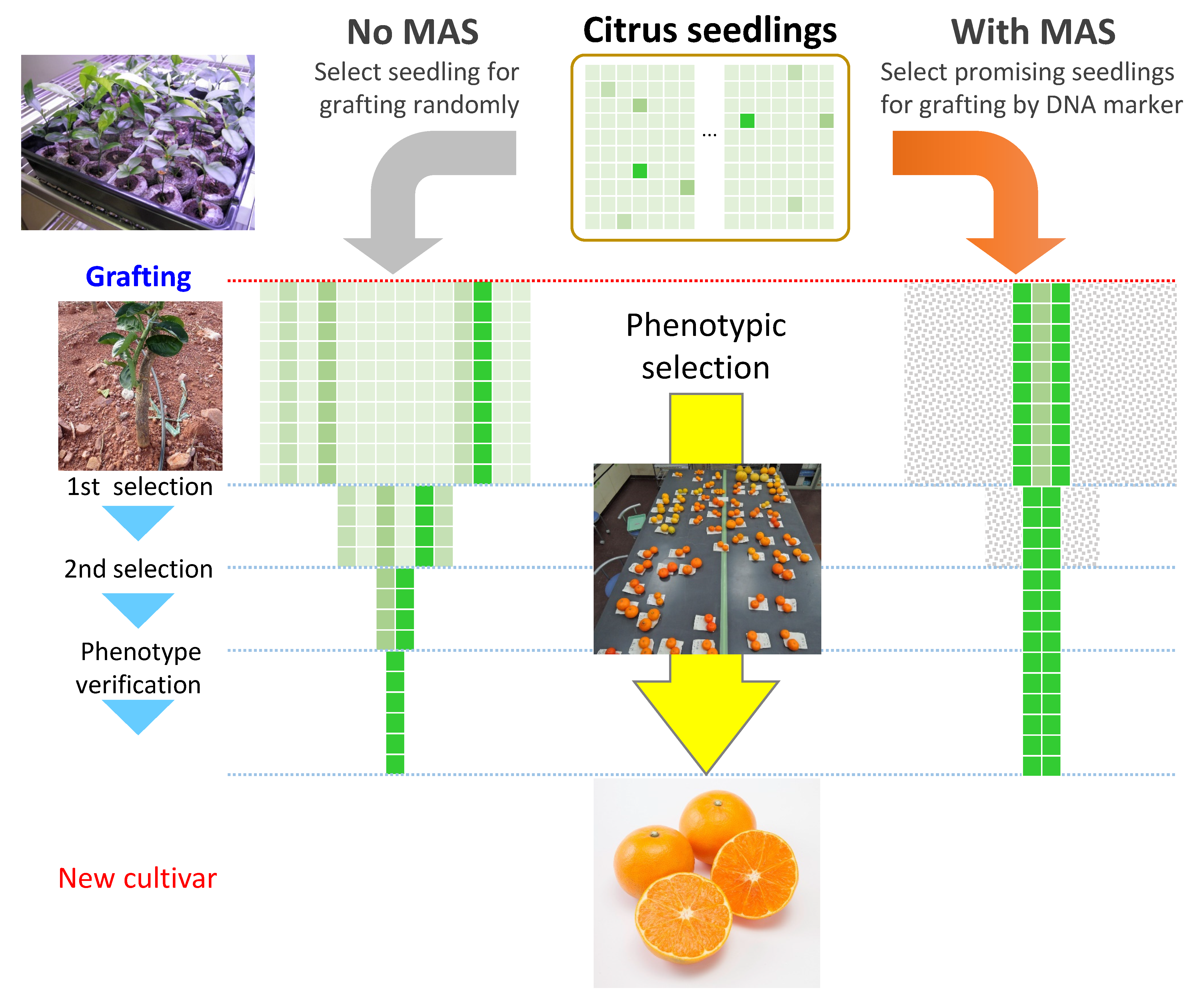
2.3. Pros and Cons of MAS for Improving Fruit Quality and the Demand for Genomic Breeding
3. Suggestions from the Citrus Genealogy for Breeding
3.1. Genomic Breeding Aims to Avoid the Breeding Trilemma
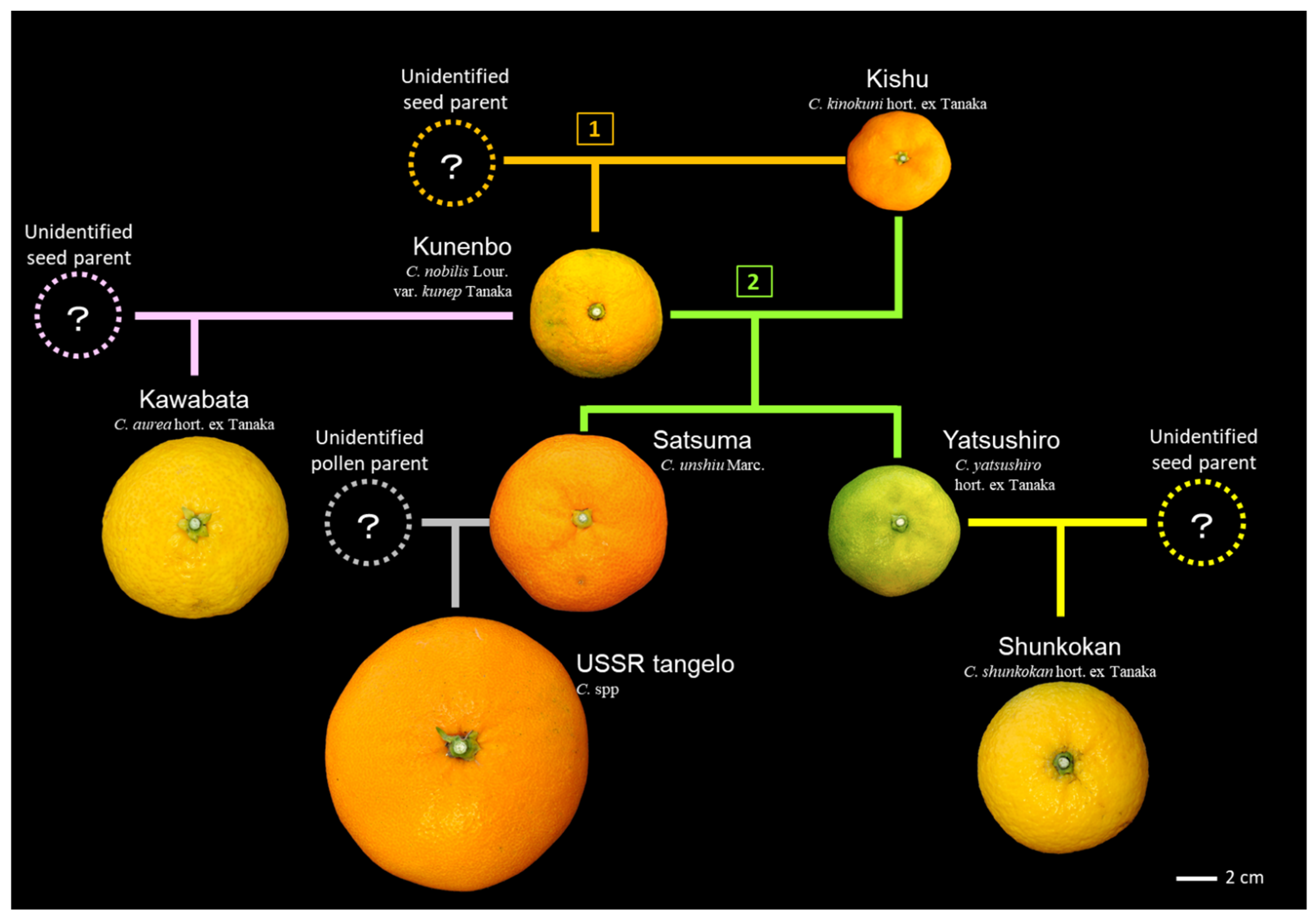
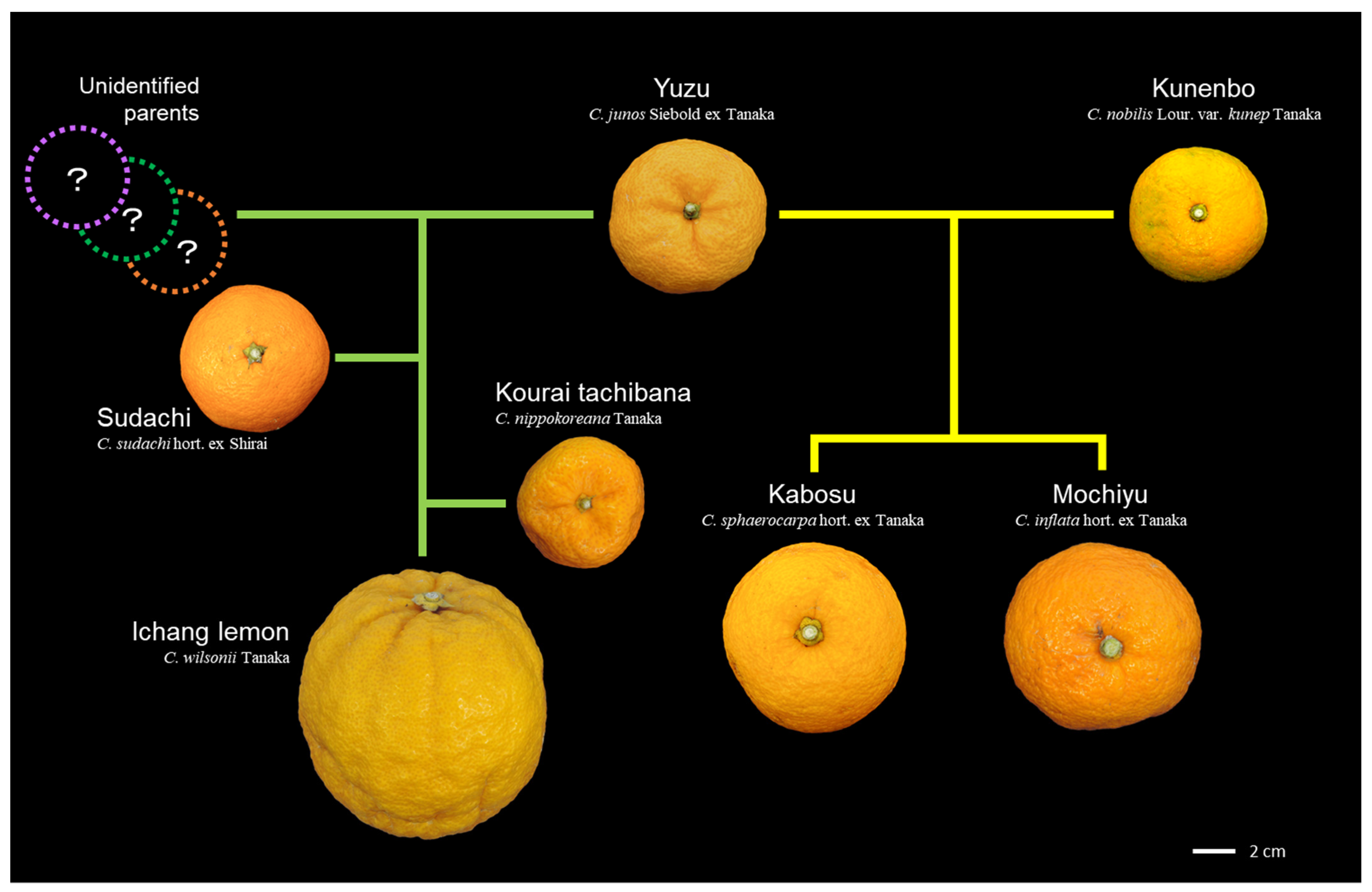
3.2. Lessons from the Revealed Genealogy of Citrus Cultivars
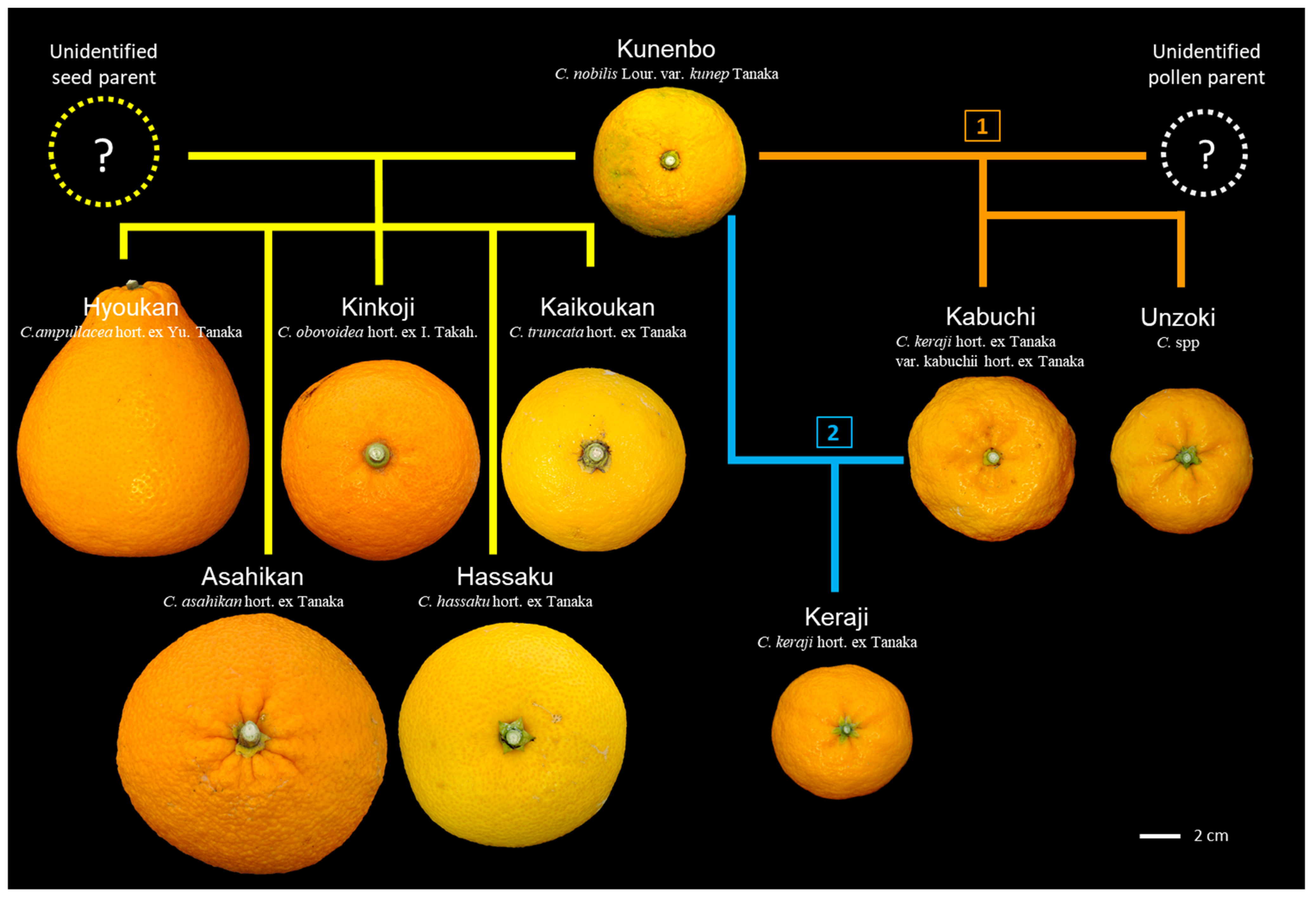
3.3. Hybrids Selected from Kishu and Kunenbo
3.4. Hybrids Selected from Yuzu
3.5. Hybrid Selected from Kunenbo
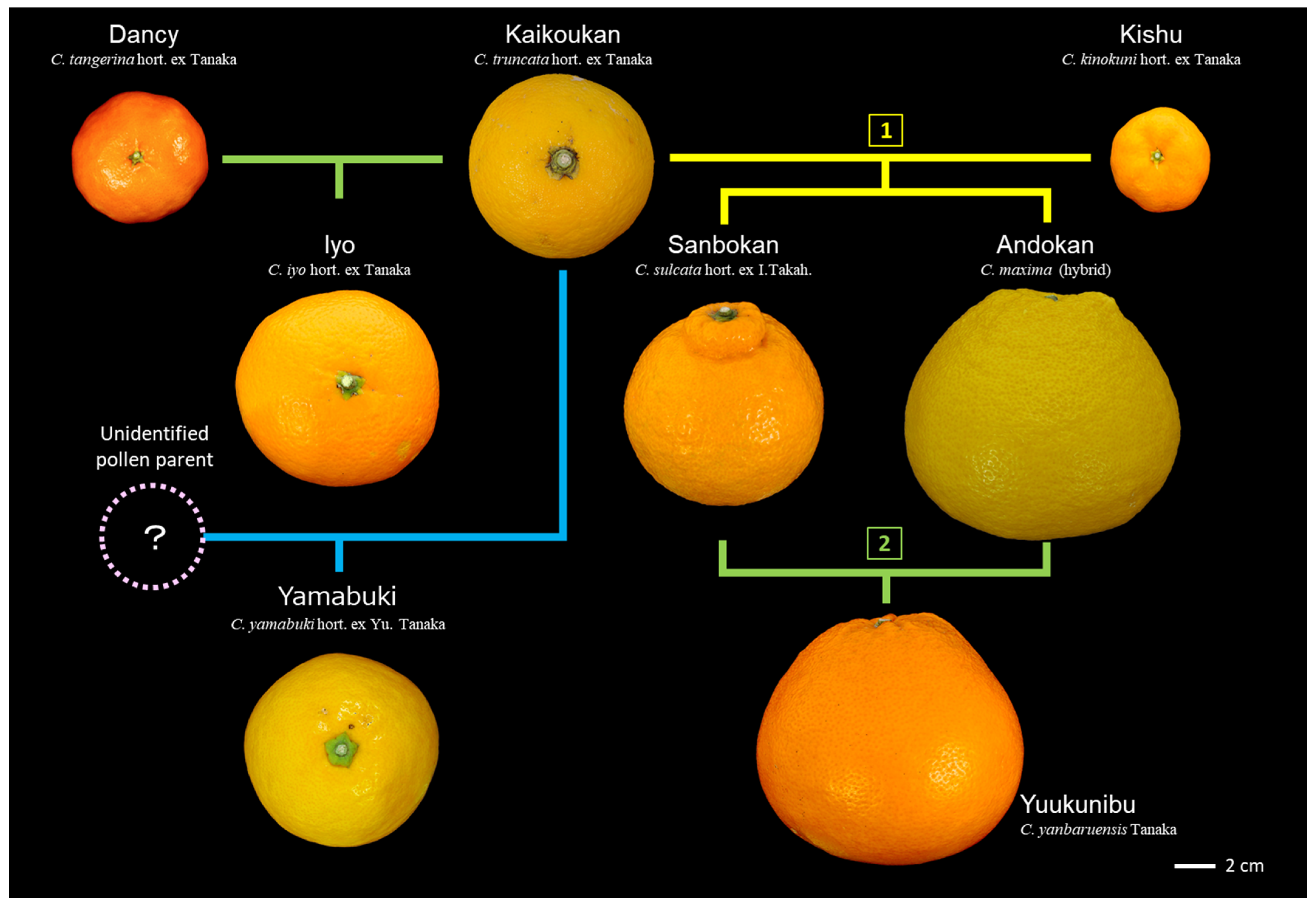
3.6. Hybrids Selected from Kaikoukan
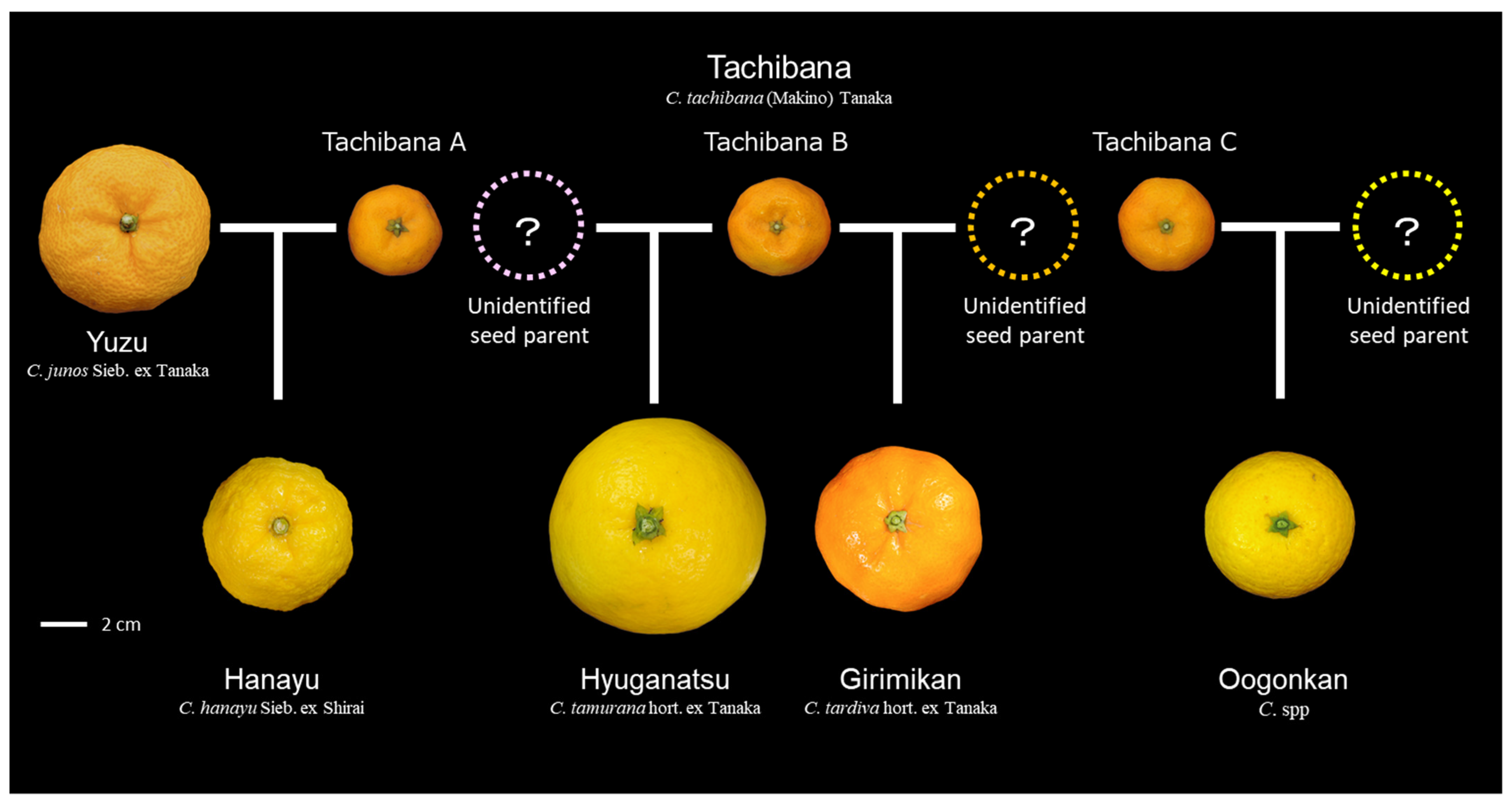
3.7. Hybrids Selected from Tachibana
4. Integration of Genomic Breeding with Genealogy to Avoid the Breeding Trilemma
5. Future Perspective
6. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Webber, H.J. History and development of the citrus industry. In The Citrus Industry I.; Reuther, W., Webber, H.J., Batchelor, L.D., Eds.; University of California Press: Berkeley, CA, USA, 1967; pp. 1–39. [Google Scholar]
- Food and Agriculture Organization, Intergovernmental Group on Citrus Fruits. Citrus Fruit Statistics 2015; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Caruso, M.; Smith, M.W.; Froelicher, Y.; Russo, G.; Gmitter, F.G. Traditional breeding. In The Genus Citrus; Elsevier: Duxford, UK, 2020; pp. 129–148. [Google Scholar]
- Nishiura, M. Citrus breeding through nucellar seedling selection. JARQ Jpn. Agric. Res. Q. 1967, 2, 15–19. [Google Scholar]
- Lamo, K.; Ji Bhat, D.; Kour, K.; Singh Solanki, S.P. Mutation Studies in Fruit Crops: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3620–3633. [Google Scholar] [CrossRef]
- Kamatyanatt, M.; Singh, S.K.; Sekhon, B.S. Mutation breeding in citrus—A review. Plant Cell Biotechnol. Mol. Biol. 2021, 22, 1–8. [Google Scholar]
- Roose, M.L.; Williams, T.E. Mutation breeding. In Citrus Genetics, Breeding and Biotechnology; Khan, I.A., Ed.; CABI, CAB International: Oxfordshire, UK, 2007; pp. 345–352. ISBN 978-0851990194. [Google Scholar]
- Cimen, B.; Yesiloglu, T.; Incesu, M.; Yilmaz, B. Studies on mutation breeding in citrus: Improving seedless types of ‘Kozan’ common orange by gamma irradiation. Sci. Hortic. 2021, 278, 109857. [Google Scholar] [CrossRef]
- Iwamasa, M.; Yamaguchi, S.; Kuriyama, T.; Nakamuta, T.; Ehara, T.; Nito, N.; Katayama, Y. Occurrence of very early mutants from the wase (early ripening) satsumas and their characteristics. Bull. Fac. Agric. Saga Univ. 1984, 55, 99–107. [Google Scholar]
- Ollitrault, P.; Navarro, L. Citrus. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer: Boston, MA, USA, 2012; pp. 623–662. ISBN 9781441907639. [Google Scholar]
- Raveh, E.; Goldenberg, L.; Porat, R.; Carmi, N.; Gentile, A.; La Malfa, S. Conventional Breeding of Cultivated Citrus Varieties. In The Citrus Genome; Springer Nature: Cham, Switzerland, 2020; pp. 33–48. [Google Scholar]
- Shimizu, T.; Aka Kacar, Y.; Cristofani-Yaly, M.; Curtolo, M.; Machado, M.A. Markers, Maps, and Marker-Assisted Selection. In The Citrus Genome; Gentile, A., La Malfa, S.D.Z., Eds.; Springer: Cham, Switzerland, 2020; pp. 107–139. ISBN 978-3-030-10799-4. [Google Scholar]
- Shimizu, T. Citrus Breeding 2.0: A novel approach integrating deciphered parentage and genomics-assisted selection. JARQ Jpn. Agric. Res. Q. 2019, 53, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Nishiura, M. Citrus breeding and bud selection in Japan. Proc. Fla. State Hort. Soc. 1964, 77, 79–83. [Google Scholar]
- Nishiura, M.; Shichijo, T.; Ueno, I.; Iwamasa, M.; Kihara, T.; Yamada, Y.; Yoshida, T.; Iwasaki, T. New citrus cultivar “Kiyomi” tangor. Bull. Fruit Tree Res. Stn. Ser. B Okitsu 1983, 10, 1–9. (In Japanese) [Google Scholar]
- NARO: National Agriculture and Food Research Organization List of the Citrus Bred Varieties. Available online: https://www.naro.go.jp/laboratory/nifts/kih/hinshu/citrus_cat/index.html (accessed on 10 April 2022). (In Japanese).
- Shimizu, T. Genomic breeding. In The Genus Citrus; Talon, M., Gmitter, F., Caruso, M., Eds.; Elsevier: Duxford, UK, 2020; pp. 149–169. ISBN 978-0-12-812163-4. [Google Scholar]
- Roose, M.L. Mapping and marker-assisted selection. In Citrus Genetics, Breeding and Biotechnology; Kahn, I.A., Ed.; CABI: Wallingford, UK, 2007; pp. 275–285. ISBN 9780851990194. [Google Scholar]
- Ohta, S.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Kuniga, T.; Yoshioka, T.; Nesumi, H.; Yoshida, T.; Omura, M. PCR primers for marker assisted backcrossing to introduce a CTV resistance gene from Poncirus trifoliata (L.) Raf. into Citrus. J. Jpn. Soc. Hortic. Sci. 2011, 80, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. QTL mapping of male sterility and transmission pattern in progeny of Satsuma mandarin. PLoS ONE 2018, 13, e0200844. [Google Scholar] [CrossRef]
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. Segregation and heritability of male sterility in populations derived from progeny of Satsuma mandarin. PLoS ONE 2016, 11, e0162408. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Endo, T.; Fujii, H.; Nakano, M.; Sugiyama, A.; Daido, G.; Ohta, S.; Yoshioka, T.; Omura, M. MITE insertion-dependent expression of CitRKD1 with a RWP-RK domain regulates somatic embryogenesis in citrus nucellar tissues. BMC Plant Biol. 2018, 18, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oraguzie, N.; Wilcox, P.; Rikkerink, E.; De Silva, N. Linkage Disequilibrium. In Association Mapping in Plants; Oraguzie, N.C., Rikkerink, E.H.A., Gardiner, S.E., de Silva, H.N., Eds.; Springer: New York, NY, USA, 2007; pp. 11–39. ISBN 978-0-387-35844-4. [Google Scholar]
- Iwata, H.; Minamikawa, M.F.; Kajiya-Kanegae, H.; Ishimori, M.; Hayashi, T. Genomics-assisted breeding in fruit trees. Breed. Sci. 2016, 66, 100–115. [Google Scholar] [CrossRef] [Green Version]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T. Genomic selection: The future of marker assisted selection and animal breeding. In Marker Assisted Selection: A Fast Track To Increase Genetic Gain in Plant And Animal Breeding? Lanteri, S., Ed.; Electronic Forum on Biotechnology in Food and Agriculture: Rome, Italy, 2003; pp. 54–59. [Google Scholar]
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Jadon, V.; Tyagi, A.; Mushtaq, M.; Jain, N.; Singh, P.K.; et al. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front. Genet. 2016, 7, 221. [Google Scholar] [CrossRef] [Green Version]
- van Nocker, S.; Gardiner, S.E. Breeding better cultivars, faster: Applications of new technologies for the rapid deployment of superior horticultural tree crops. Hortic. Res. 2014, 1, 14022. [Google Scholar] [CrossRef] [Green Version]
- Desta, Z.A.; Ortiz, R. Genomic selection: Genome-wide prediction in plant improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef]
- Minamikawa, M.F.; Nonaka, K.; Kaminuma, E.; Kajiya-Kanegae, H.; Onogi, A.; Goto, S.; Yoshioka, T.; Imai, A.; Hamada, H.; Hayashi, T.; et al. Genome-wide association study and genomic prediction in citrus: Potential of genomics-assisted breeding for fruit quality traits. Sci. Rep. 2017, 7, 4721. [Google Scholar] [CrossRef]
- Combrink, N.K.; Labuschagne, M.T.; Bijzet, Z. Variation of fruit size and shape in Kiyomi tangor families. Sci. Hortic. 2013, 162, 357–364. [Google Scholar] [CrossRef]
- Combrink, N.K.; Bijzet, Z.; Sippel, A.D.; Booyse, M.; Labuschagne, M.T. Genotypic variation of rind colour in citrus tangor Kiyomi families. Acta Hortic. 2015, 1065, 439–448. [Google Scholar] [CrossRef]
- Kita, M.; Nesumi, H.; Kuniga, T.; Nakajima, N.; Yoshioka, T.; Ohta, S.; Takishita, F.; Nakano, M.; Ogawa, K.; Yoshida, T.; et al. New Citrus Cultivar ‘Aurastar’. J. NARO Res. Dev. 2021, 7, 1–8. (In Japanese) [Google Scholar] [CrossRef]
- Yoshida, T.; Nesumi, H.; Nakajima, N.; Kuniga, T. New citrus parental lines “Kankitsu Chukanbohon Nou 7 Gou” and “Kankitsu Chukanbohon Nou 8 Gou” for breeding new cultivars with immunity to citrus tristeza virus. Jpn. J. Hortic. Sci. 2005, 74, 323. (In Japanese) [Google Scholar]
- Serra, O.; Donoso, J.M.; Picañol, R.; Batlle, I.; Howad, W.; Eduardo, I.; Arús, P. Marker-assisted introgression (MAI) of almond genes into the peach background: A fast method to mine and integrate novel variation from exotic sources in long intergeneration species. Tree Genet. Genomes 2016, 12, 96. [Google Scholar] [CrossRef]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.A.; Sugimoto, C.; Kinjo, H.; Azama, C.; Mitsube, F.; Talon, M.; Gmitter, F.G.; Rokhsar, D.S. Diversification of mandarin citrus by hybrid speciation and apomixis. Nat. Commun. 2021, 12, 4377. [Google Scholar] [CrossRef]
- Shimizu, T.; Kitajima, A.; Nonaka, K.; Yoshioka, T.; Ohta, S.; Goto, S.; Toyoda, A.; Fujiyama, A.; Mochizuki, T.; Nagasaki, H.; et al. Hybrid origins of citrus varieties inferred from DNA marker analysis of nuclear and organelle genomes. PLoS ONE 2016, 11, e0166969. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T. Species Problem in Citrus: A Critical Study of Wild and Cultivated Units of Citrus, Based upon Field Studies in Their Native Homes (Revisio Aurantiacearum IX); Japanese Society for the Promotion of Science: Tokyo, Japan, 1954. [Google Scholar]
- Tanaka, T. A Revision of Osmocitrus, a section of the genus Citrus: Revisio Aurantiacearum XIII. Bull. Univ. Osaka Prefect. Ser. B Agric. Biol. 1960, 10, 9–13. [Google Scholar]
- Shimizu, T.; Kitajima, A.; Nonaka, K.; Yoshioka, T.; Ohta, S.; Goto, S.; Kaminuma, E.; Nakamura, Y. A model for the domestication and diversification processes of modern citrus cultivars in Japan. In Proceedings of the IV International Symposium on Citrus Biotechnology, Canelones, Uruguay, 16–18 April 2018; pp. 7–14. [Google Scholar] [CrossRef]
- Shimizu, T.; Tanizawa, Y.; Mochizuki, T.; Nagasaki, H.; Yoshioka, T.; Toyoda, A.; Fujiyama, A.; Kaminuma, E.; Nakamura, Y. Draft sequencing of the heterozygous diploid genome of Satsuma (Citrus unshiu Marc.) using a hybrid assembly approach. Front. Genet. 2017, 8, 180. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T. On the origin, affinity and scientific name of the Satsuma orange (2). Stud. Citrol. 1927, 1, 26–41. (In Japanese) [Google Scholar]
- Tanaka, T. Citrologia: Semi-Centennial Commemoration Papers on Citrus Studies; Citrologia Supporting Foundation for Tanaka International Citrus Institute: Sakai, Osaka, Japan, 1961. [Google Scholar]
- Swingle, W.T. The botany of Citrus and its wild relatives. In The Citrus Industry; Reuther, W., Webber, H.J., Batchelor, L.D., Eds.; University of California Press: Berkeley, CA, USA, 1967; pp. 190–430. [Google Scholar]
- Hirai, M.; Mitsue, S.; Kita, K.; Kajiura, I. A Survey and isozyme analysis of wild mandarin, tachibana (Citrus tachibana (Mak.) Tanaka) growing in Japan. J. Jpn. Soc. Hortic. Sci. 1990, 59, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Nagakura, K.; Okuda, M.; Inagi, H.; Okuda, I.; Endo, S. Evaluation of Genetic Diversity in Wild Tachibana Population of Heda, Shizuoka, Using DNA Marker Analysis, and Stable Maintenance of the Population. Hortic. Res. Jpn. 2020, 19, 141–149. (In Japanese) [Google Scholar] [CrossRef]
- Eduardo, I.; Alegre, S.; Alexiou, K.G.; Arús, P. Resynthesis: Marker-Based Partial Reconstruction of Elite Genotypes in Clonally-Reproducing Plant Species. Front. Plant Sci. 2020, 11, 1205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yin, L.; Wang, M.; Yuan, X.; Liu, X. Factors Affecting the Accuracy of Genomic Selection for Agricultural Economic Traits in Maize, Cattle, and Pig Populations. Front. Genet. 2019, 10, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Year of Release | YTD | Harvest | Fruit Size (g) | Peelability | Seeds | Features |
|---|---|---|---|---|---|---|---|
| Kiyomi | 1979 | 30 | Late March | ~200 | Moderate | SL | A tangor (mandarin x orange). Orange-like aroma and juicy |
| Seihou | 1988 | 17 | Late January | ~200 | Hard | SL | Large fruit with orange-like good flavor and soft flesh |
| Tshunokaori | 1990 | 18 | March–April | ~160 | Easy | SL | High SSC (>13), and no rind-puffing |
| Amakusa | 1993 | 11 | August–January | ~200 | Hard | -- | High SSC (>12), and less alternate bearing |
| Youkou | 1995 | 23 | January–February | 250–300 | Easy | SL | Large fruit with good flavor and soft flesh |
| Harumi | 1996 | 17 | January | 180–200 | Easy | SL | Large fruit with good flavor and soft flesh |
| Akemi | 1996 | 21 | March | 160–180 | Moderate | SL | High SSC (>12), grenadine rind, and soft flesh |
| Amaka | 1996 | 22 | August–January | 200–250 | Easy | SL | Soft fruit with good orange-like aroma |
| Nishinokaori | 1997 | 31 | August–January | 100–180 | Easy | SL | High SSC (>12), orange-like flavor, and soft flesh |
| Setoka | 1998 | 14 | February | 200–280 | Easy | SL | Large fruit with good flavor, high SSC (>12), and soft flesh |
| Harehime | 2001 | 11 | August | 180 | Easy | SL | Early type with good flavor, juicy, and soft flesh |
| Tamami | 2004 | 24 | January | 150 | Easy | -- | High SSC (>12) and good orange-like flavor |
| Reikou | 2004 | 20 | January | 210 | Moderate | SL | High SSC (>12), good flavor, and red-skin |
| Tsunokagayaki | 2008 | 24 | January–February | 180–250 | Easy | SL | High SSC (>13), soft fruit, BCP rich, and suitable for greenhouse production |
| Tsunonozomi | 2011 | 37 | August | 190 | Easy | -- | Early type with good flavor, high SSC (>12), juicy, and less rind puffing |
| Asumi | 2013 | 21 | February | 150 | Moderate | SL | Very high SSC (>15), orange-like flavor, and BCP rich |
| Shiranuhi * | February–March | 230 | Easy | SL | High SSC (>14), soft fruit, good flavor, and unique fruit shape |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, T. Breeding New Premium Quality Cultivars by Citrus Breeding 2.0 in Japan: An Integrative Approach Suggested by Genealogy. Horticulturae 2022, 8, 559. https://doi.org/10.3390/horticulturae8060559
Shimizu T. Breeding New Premium Quality Cultivars by Citrus Breeding 2.0 in Japan: An Integrative Approach Suggested by Genealogy. Horticulturae. 2022; 8(6):559. https://doi.org/10.3390/horticulturae8060559
Chicago/Turabian StyleShimizu, Tokurou. 2022. "Breeding New Premium Quality Cultivars by Citrus Breeding 2.0 in Japan: An Integrative Approach Suggested by Genealogy" Horticulturae 8, no. 6: 559. https://doi.org/10.3390/horticulturae8060559