
Antifungal Activity of Volatile Organic Compounds from Bacillus velezensis CE 100 against Colletotrichum gloeosporioides

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antagonistic Bacteria and Pathogenic Fungi
2.2. Antifungal Activity of VOCs from Bacillus velezensis CE 100 against Spore Germination and Mycelial Growth of Colletotrichum gloeosporioides
2.3. Collection and Identification of VOCs Emitted by Bacillus velezensis CE 100 Using HS-SPME/GC-MS
2.4. Antifungal Activity of the VOCs Produced by B. velezensis CE 100 against the Germination of Spore and Mycelial Growth of Colletotrichum gloeosporioides
2.5. Statistical Analysis
3. Results
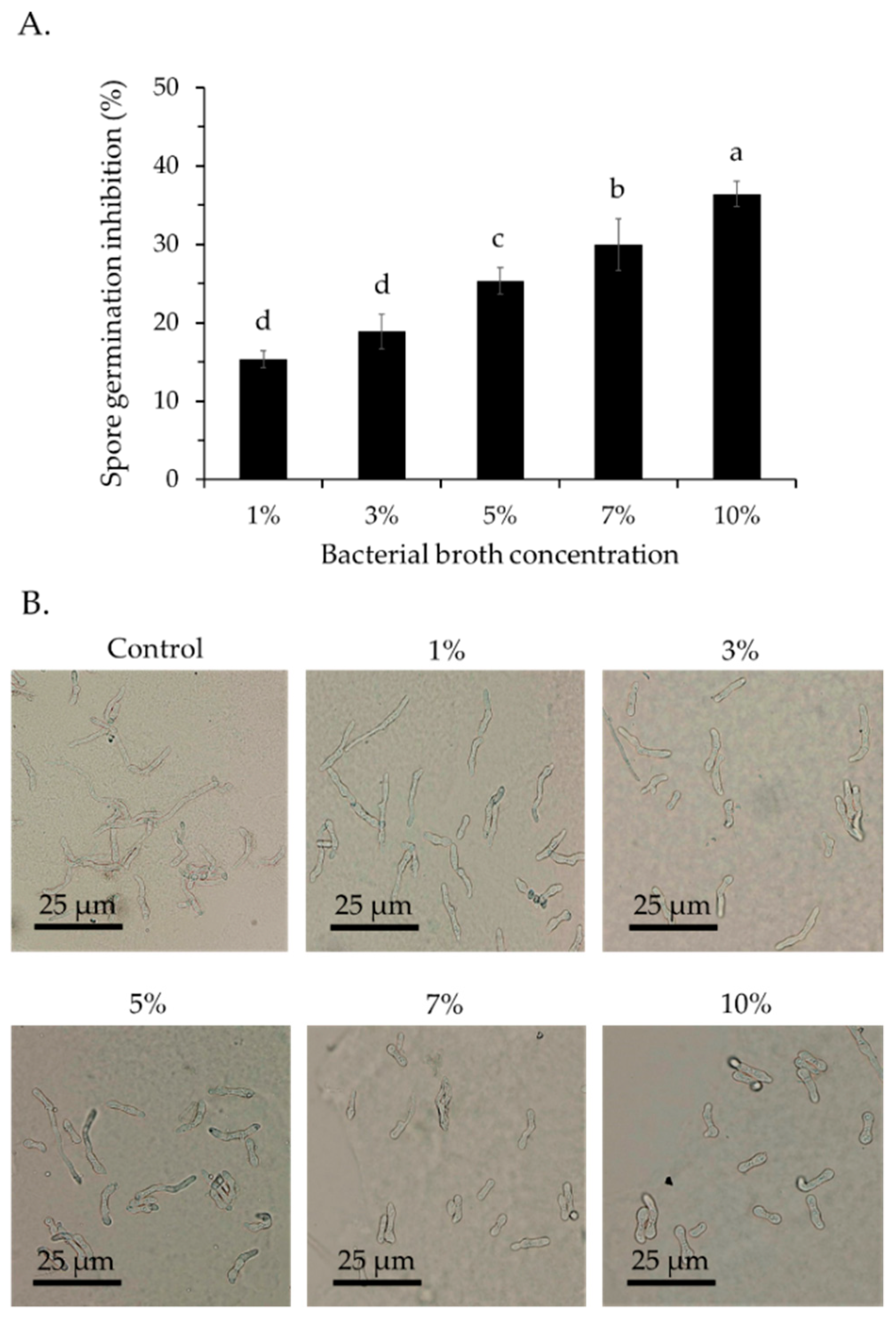
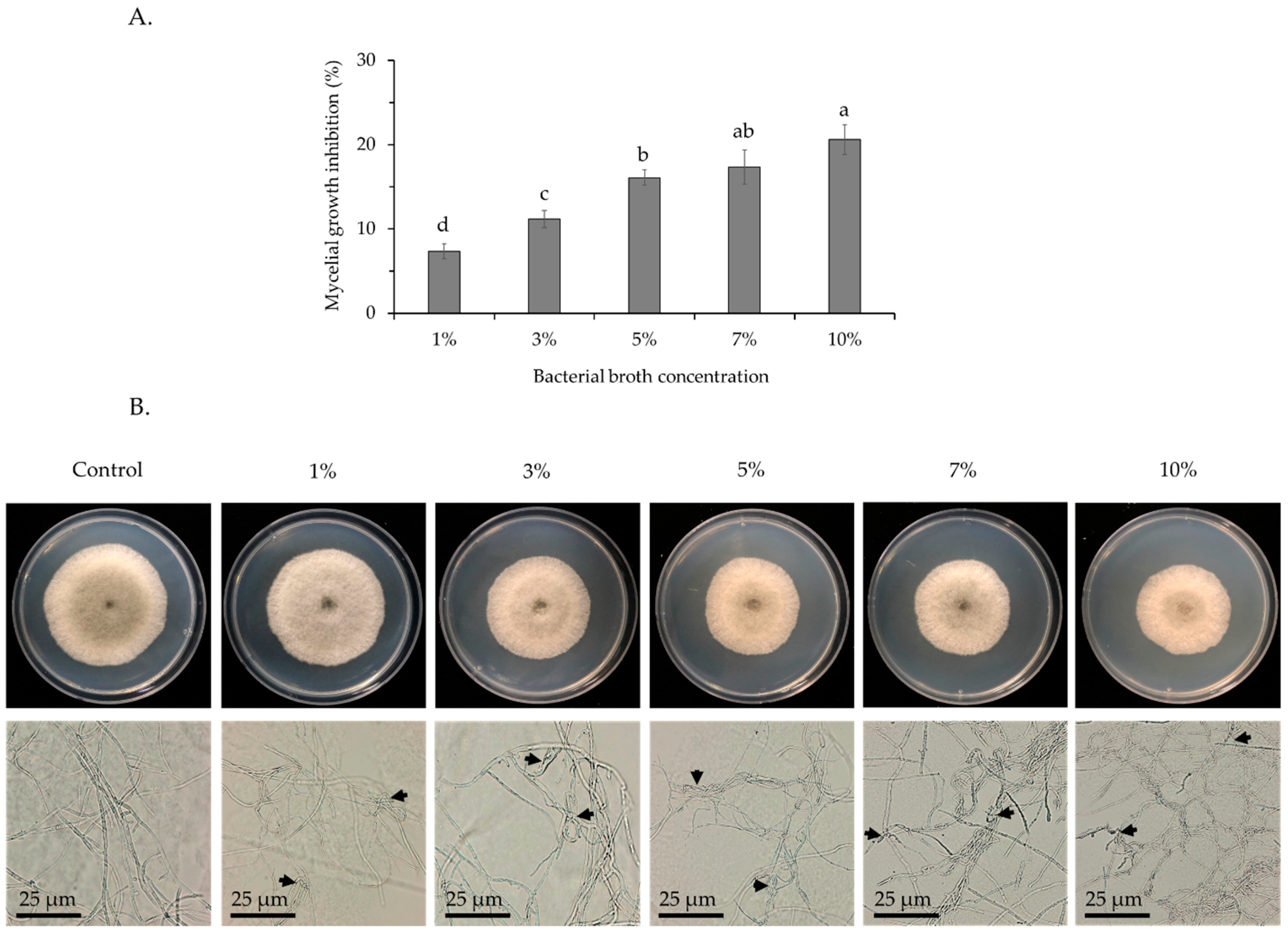
3.1. Antifungal Activity of the VOCs from Bacillus velezensis CE 100 against Spore Germination and Mycelial Growth of Colletotrichum gloeosporioides
3.2. HS-SPME/GC-MS Analysis of VOCs Emitted by Bacillus velezensis CE 100
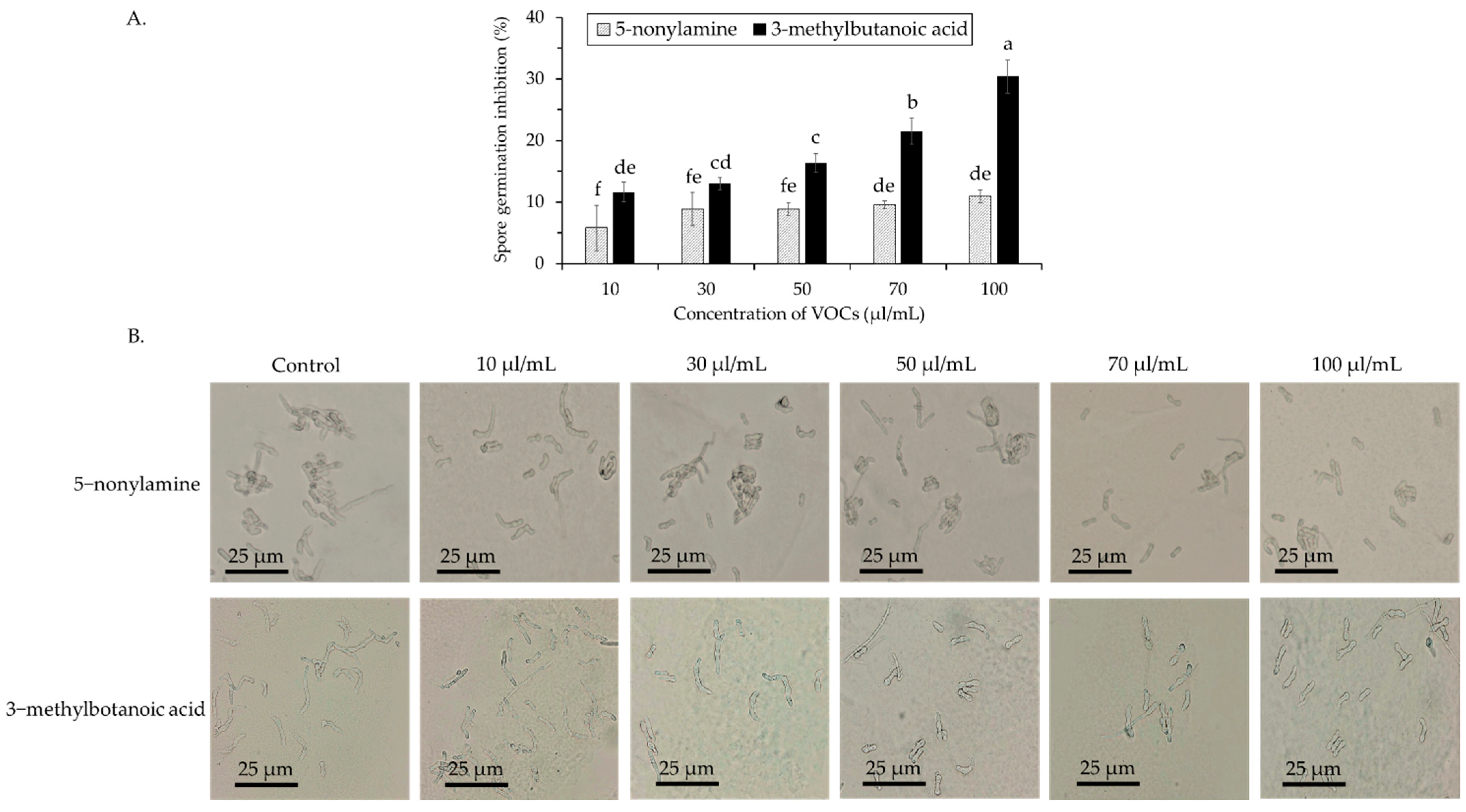
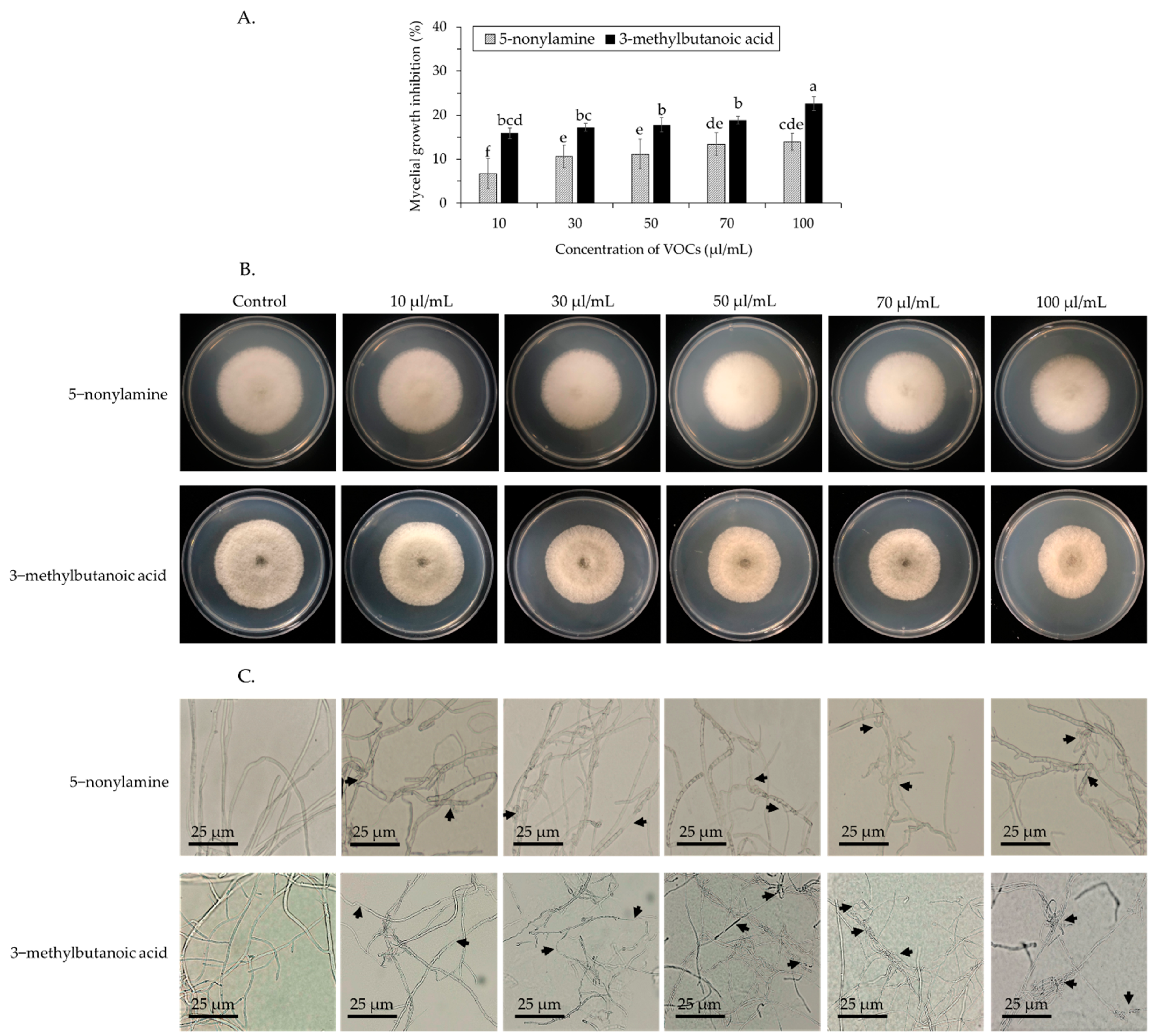
3.3. Antifungal Activity of 5-Nonylamine and 3-Methylbutanoic Acid against Spore Germination and Mycelial Growth of Colletotrichum gloeosporioides
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Devasahayam, H.L. Illustrated Plant Pathology: Basic Concepts, 1st ed.; New India Publishing Agency: New Delhi, India, 2009; pp. 10–11. [Google Scholar]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammondus, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Kulshrestha, S. Colletotrichum Gloeosporioides: An Anthracnose Causing Pathogen of Fruits and Vegetables. Biosci. Biotechnol. Res. Asia 2015, 12, 1233–1246. [Google Scholar] [CrossRef]
- Ojola, P.O.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of Rice Thaumatin-like Protein (Ostlp) Gene in Transgenic Cassava Results in Enhanced Tolerance to Colletotrichum gloeosporioides f. sp. manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-H.; Fan, K.; Li, D.-W.; Han, C.-M.; Qu, Y.-Y.; Qi, Y.-K.; Wu, X.-Q. Identification, Virulence and Fungicide Sensitivity of Colletotrichum gloeosporioides ss Responsible for Walnut Anthracnose Disease in China. Plant Dis. 2020, 104, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Choub, V.; Ajuna, H.B.; Won, S.-J.; Moon, J.-H.; Choi, S.-I.; Maung, C.E.H.; Kim, C.-W.; Ahn, Y.S. Antifungal Activity of Bacillus velezensis CE 100 against Anthracnose Disease (Colletotrichum gloeosporioides) and Growth Promotion of Walnut (Juglans regia L.) Trees. Int. J. Mol. Sci. 2021, 22, 10438. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Won, S.-J.; Moon, J.-H.; Lee, U.; Park, Y.-S.; Maung, C.E.H.; Ajuna, H.B.; Ahn, Y.S. Bacillus licheniformis PR2 Controls Fungal Diseases and Increases Production of Jujube Fruit under Field Conditions. Horticulturae 2021, 7, 49. [Google Scholar] [CrossRef]
- Cannon, P.; Damm, U.; Johnston, P.; Weir, B. Colletotrichum—Current Status and Future Directions. Studies Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Ojiambo, P.; Louws, F. Initial Inoculum and Spatial Dispersal of Colletotrichum gloeosporioides, the causal agent of Strawberry Anthracnose Crown Rot. Plant Dis. 2015, 99, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Ishii, H.; Watanabe, H.; Yamaoka, Y.; Schnabel, G. Sensitivity to Fungicides in Isolates of Colletotrichum gloeosporioides and C. acutatum Species Complexes and Efficacy against Anthracnose Diseases. Pestici. Biochem. Physiol. 2022, 182, 105049. [Google Scholar] [CrossRef]
- Sanders, G.M.; Korsten, L.; Wehner, F. Survey of Fungicide Sensitivity in Colletotrichum gloeosporioides from Different Avocado and Mango Production Areas in South Africa. Eur. J. Plant Pathol. 2000, 106, 745–752. [Google Scholar] [CrossRef]
- Dowling, M.; Peres, N.; Villani, S.; Schnabel, G. Managing Colletotrichum on Fruit Crops: A “Complex” Challenge. Plant Dis. 2020, 104, 2301–2316. [Google Scholar] [CrossRef]
- López Zapata, S.P.; Castaño Zapata, J. In Vitro Effect of Four Fungicides on Colletotrichum gloeosporioides Causing Anthracnosis on The Red Globe Grape Variety. Rev. Acad. Colomb. Cienc. Exactas Fis. Nat. 2020, 44, 747–758. [Google Scholar] [CrossRef]
- Wightwick, A.; Walters, R.; Allinson, G.; Reichman, S.; Menzies, N. Environmental Risks of Fungicides Used in Horticultural Production Systems. Fungicides 2010, 1, 273–304. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of Action and Possible Impact on Nontarget Microorganisms. ISRN Ecol. 2011, 2011, 130289. [Google Scholar] [CrossRef] [Green Version]
- Bozdogan, A.M. Assessment of Total Risk on Non-Target Organisms in Fungicide Application for Agricultural Sustainability. Sustainability 2014, 6, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, N.; Kumar, V.; Datta, S.; Wani, A.B.; Singh, D.; Singh, K.; Singh, J. Toxicity, Monitoring and Biodegradation of the Fungicide Carbendazim. Environ. Chem. Lett. 2016, 14, 317–329. [Google Scholar] [CrossRef]
- Chung, W.-H.; Chung, W.-C.; Peng, M.-T.; Yang, H.-R.; Huang, J.-W. Specific Detection of Benzimidazole Resistance in Colletotrichum gloeosporioides from Fruit Crops by PCR-RFLP. New Biotechnol. 2010, 27, 17–24. [Google Scholar] [CrossRef]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A. Mechanisms Influencing the Evolution of Resistance to Qo Inhibitor Fungicides. Pest. Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef]
- Hahn, M. The Rising Threat of Fungicide Resistance in Plant Pathogenic Fungi: Botrytis as A Case Study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Cao, G.; Jiang, S.; Han, S.; Yang, C.; Wan, X.; Zhang, F.; Chen, L.; Xiao, J.; Zhu, P. Identification of the Anthracnose Fungus of Walnut (Juglans spp.) and Resistance Evaluation through Physiological Responses of Resistant vs. Susceptible Hosts. Plant Pathol. 2021, 70, 1219–1229. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- Magnusson, E.; Cranfield, J. Consumer Demand for Pesticide-Free Food Products in Canada: A Probit Analysis. Can. J. Agric. Econ. 2005, 53, 67–81. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, J. Consumer Preference and Demand for ORGANIC FOOD: EVIDENCE from a Vermont Survey. Proceedings of The American Agricultural Economics Association Annual Meeting, Montreal, QC, Canada, 27–30 July 2003; pp. 1–24. [Google Scholar] [CrossRef]
- Kong, W.-L.; Li, P.-S.; Wu, X.-Q.; Wu, T.-Y.; Sun, X.-R. Forest Tree Associated Bacterial Diffusible and Volatile Organic Compounds against Various Phytopathogenic Fungi. Microorganisms 2020, 8, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asari, S.; Matzén, S.; Petersen, M.A.; Bejai, S.; Meijer, J. Multiple effects of Bacillus amyloliquefaciens Volatile Compounds: Plant Growth Promotion and Growth Inhibition of Phytopathogens. FEMS Microbiol. Ecol. 2016, 92, fiw070. [Google Scholar] [CrossRef] [Green Version]
- Choub, V.; Maung, C.E.H.; Won, S.-J.; Moon, J.-H.; Kim, K.Y.; Han, Y.S.; Cho, J.-Y.; Ahn, Y.S. Antifungal Activity of Cyclic Tetrapeptide from Bacillus velezensis CE 100 against Plant Pathogen Colletotrichum gloeosporioides. Pathogens 2021, 10, 209. [Google Scholar] [CrossRef]
- Calvo, H.; Mendiara, I.; Arias, E.; Gracia, A.P.; Blanco, D.; Venturini, M.E. Antifungal Activity of the Volatile Organic Compounds Produced by Bacillus velezensis Strains against Postharvest Fungal Pathogens. Postharvest Biol. Technol. 2020, 166, 111208. [Google Scholar] [CrossRef]
- Li, X.-Y.; Mao, Z.-C.; Wu, Y.-X.; Ho, H.-H.; He, Y.-Q. Comprehensive Volatile Organic Compounds Profiling of Bacillus species with Biocontrol Properties by Headspace Solid-Phase Microextraction with Gas Chromatography-Mass Spectrometry. Biol. Sci. Technol. 2015, 25, 132–143. [Google Scholar] [CrossRef]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The Antimicrobial Volatile Power of The Rhizospheric Isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Li, P.; Wu, S.; Zhou, M.; Zhi, R.; Gao, H. Volatile Organic Compounds (VOCs) from Bacillus subtilis CF-3 Reduce Anthracnose and Elicit Active Defense Responses in Harvested Litchi Fruits. AMB Express 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.; Hung, R.; Lee, S.; Padhi, S. 18 Fungal and Bacterial Volatile Organic Compounds: An Overview and Their Role as Ecological Signaling Agents. Fungal Assoc. 2012, 373–393. [Google Scholar] [CrossRef]
- Wheatley, R. The Consequences of Volatile Organic Compound Mediated Bacterial and Fungal Interactions. Antonie Van Leeuwenhoek 2002, 81, 357–364. [Google Scholar] [CrossRef]
- Syed-Ab-Rahman, S.F.; Carvalhais, L.C.; Chua, E.T.; Chung, F.Y.; Moyle, P.M.; Eltanahy, E.G.; Schenk, P.M. Soil Bacterial Diffusible and Volatile Organic Compounds Inhibit Phytophthora capsici and Promote Plant Growth. Sci. Total Environ. 2019, 692, 267–280. [Google Scholar] [CrossRef]
- Martínez-Cámara, R.; Montejano-Ramírez, V.; Moreno-Hagelsieb, G.; Santoyo, G.; Valencia-Cantero, E. The Volatile Organic Compound Dimethylhexadecylamine Affects Bacterial Growth and Swarming Motility of Bacteria. Folia Microbiol. 2020, 65, 523–532. [Google Scholar] [CrossRef]
- Tabbal, S.; El Aroussi, B.; Bouchard, M.; Marchand, G.; Haddad, S. A New Headspace Solid-Phase Microextraction Coupled with Gas Chromatography-Tandem Mass Spectrometry Method for The Simultaneous Quantification of 21 Microbial Volatile Organic Compounds in Urine and Blood. Chemosphere 2022, 296, 133901. [Google Scholar] [CrossRef] [PubMed]
- He, C.-N.; Ye, W.-Q.; Zhu, Y.-Y.; Zhou, W.-W. Antifungal Activity of Volatile Organic Compounds Produced by Bacillus methylotrophicus and Bacillus thuringiensis against Five Common Spoilage Fungi on Loquats. Molecules 2020, 25, 3360. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Korsten, L. Effect of Volatile Compounds Produced by Bacillus Strains on Postharvest Decay in Citrus. Biol. Control 2010, 53, 122–128. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of Endophytic Bacillus velezensis ZSY-1 Strain and Antifungal Activity of Its Volatile Compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Hueck, H.J.; Adema, D.M.; Wiegmann, J.R. Bacteriostatic, Fungistatic, and Algistatic Activity of Fatty Nitrogen Compounds. Appl. Microbiol. 1966, 14, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, Y.; Yuan, Y.; Chu, D.; Cao, J.; Sun, G.; Ai, Y.; Cui, Z.; Zhang, Y.; Wang, F. Identification of Non-Volatile and Volatile Organic Compounds Produced by Bacillus siamensis LZ88 and Their Antifungal Activity against Alternaria alternata. Biol. Control 2022, 169, 104901. [Google Scholar] [CrossRef]
- Choi, T.G.; Maung, C.E.H.; Lee, D.R.; Henry, A.B.; Lee, Y.S.; Kim, K.Y. Role of Bacterial Antagonists of Fungal Pathogens, Bacillus thuringiensis KYC and Bacillus velezensis CE 100 in Control of Root-Knot Nematode, Meloidogyne Incognita and Subsequent Growth Promotion of Tomato. Biocontrol Sci. Technol. 2020, 30, 685–700. [Google Scholar] [CrossRef]
- Moon, J.H.; Won, S.J.; Maung, C.E.H.; Choi, J.H.; Choi, S.I.; Ajuna, H.B.; Ahn, Y.S. Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes the Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings. Microorganisms 2021, 9, 821. [Google Scholar] [CrossRef] [PubMed]
- Won, S.J.; Moon, J.H.; Ajuna, H.B.; Choi, S.I.; Maung, C.E.H.; Lee, S.; Ahn, Y.S. Biological Control of Leaf Blight Disease Caused by Pestalotiopsis maculans and Growth-Promotion of Quercus acutissima Carruth Container Seedlings Using Bacillus velezensis CE 100. Int. J. Mol. Sci. 2021, 22, 11296. [Google Scholar] [CrossRef]
- Méndez-Bravo, A.; Cortazar-Murillo, E.M.; Guevara-Avendaño, E.; Ceballos-Luna, O.; Rodríguez-Haas, B.; Kiel-Martínez, A.L.; Hernández-Cristóbal, O.; Guerrero-Analco, J.A.; Reverchon, F. Plant Growth-Promoting Rhizobacteria Associated with Avocado Display Antagonistic Activity against Phytophthora cinnamomi through Volatile Emissions. PLoS ONE 2018, 13, e0194665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Raza, W.; Shen, Q.; Huang, Q. Antifungal activity of Bacillus amyloliquefaciens NJN-6 Volatile Compounds against Fusarium oxysporum f. sp. cubense. Appl. Environ. Microbiol. 2012, 78, 5942–5944. [Google Scholar] [CrossRef] [Green Version]
- Novinson, T.; Robins, R.K.; Matthews, T.R. Synthesis and Antifungal Properties of Certain 7-Alkylaminopyrazolo [1, 5-a] pyrimidines. J. Med. Chem. 1977, 20, 296–299. [Google Scholar] [CrossRef]
- Ravikumar, T.; Murata, H.; Koepsel, R.R.; Russell, A.J. Surface-Active Antifungal Polyquaternary Amine. Biomacromolecules 2006, 7, 2762–2769. [Google Scholar] [CrossRef] [PubMed]
- Chobe, S.S.; Adole, V.A.; Deshmukh, K.P.; Pawar, T.B.; Jagdale, B.S. Poly (Ethylene Glycol) (PEG-400): A Green Approach towards the Synthesis of Novel Pyrazolo [3, 4-d] Pyrimidine-6-Amines Derivatives and Their Antimicrobial Screening. Arch. Appl. Sci. Res. 2014, 6, 61–66. Available online: http://www.scholarsresearchlibrary.com (accessed on 16 May 2022).
- Zou, C.-S.; Mo, M.-H.; Gu, Y.-Q.; Zhou, J.-P.; Zhang, K.-Q. Possible Contributions of Volatile-Producing Bacteria to Soil Fungistasis. Soil Biol. Biochem. 2007, 39, 2371–2379. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Wang, Y.; Guo, Q.; Lu, X.; Li, S.; Ma, P. Lipopeptides, A Novel Protein, and Volatile Compounds Contribute to The Antifungal Activity of The Biocontrol Agent Bacillus atrophaeus CAB-1. Appl. Microbiol. Biotechnol. 2013, 97, 9525–9534. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Sinha, M.; Singh, H.; Patel, R.S.; Ghosh, S.; Sardana, K.; Ghosh, S.; Sengupta, S. Mechanistic Insight into The Antifungal Effects of A Fatty Acid Derivative against Drug-Resistant Fungal Infections. Front. Microbiol. 2020, 11, 2116. [Google Scholar] [CrossRef]
- Ando, H.; Hatanaka, K.; Ohata, I.; Yamashita-Kitaguchi, Y.; Kurata, A.; Kishimoto, N. Antifungal Activities of Volatile Substances Generated by Yeast Isolated from Iranian Commercial Cheese. Food Control 2012, 26, 472–478. [Google Scholar] [CrossRef]
- Strobel, G.A.; Spang, S.; Kluck, K.; Hess, W.; Sears, J.; Livinghouse, T. Synergism among Volatile Organic Compounds Resulting in Increased Antibiosis in Oidium spp. FEMS Microbiol. Lett. 2008, 283, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Sant, D.; Tupe, S.; Ramana, C.; Deshpande, M. Fungal Cell Membrane—Promising Drug Target for Antifungal Therapy. J. Appl. Microbiol. 2016, 121, 1498–1510. [Google Scholar] [CrossRef] [Green Version]
- Free, S.J. Fungal Cell Wall Organization and Biosynthesis. Adv Genet. 2013, 81, 33–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-W.; Wei, M.; Zhu, B.-Y.; Du, Y.-C.; Feng, L. Antagonistic Activities of Volatiles from Four Strains of Bacillus spp. and Paenibacillus spp. against Soil-Borne Plant Pathogens. Agric. Sci. China 2008, 7, 1104–1114. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Identified Compounds | Molecular Formula | CAS Number | RT (min) | Relative Peak Area (%) | Control/Blank |
|---|---|---|---|---|---|---|
| Amine | 5-Nonylamine | C9H21N | 112−20−9 | 1.70 | 39.5 | Nd |
| Acid | 3-Methylbutanoic acid | C5H10O2 | 140−67−0 | 1.96 | 2.92 | Nd |
| Ester | Cyclotrisiloxane hexamethyl | C6H18O3Si3 | 503−74−2 | 5.45 | 3.21 | * |
| Cyclotetrasiloxane octamethyl | C8H24O4Si4 | 541−05−9 | 8.96 | 16.22 | * | |
| Cyclopentasiloxane decamethyl | C10H30O5Si5 | 556−67−2 | 11.66 | 14.05 | * | |
| Cyclohexasiloxane dodecamethyl | C12H36O6Si6 | 514−02−6 | 14.25 | 4.2 | * | |
| Methyl | Estragole | C10H12O | 140−67−0 | 12.69 | 19.9 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choub, V.; Won, S.-J.; Ajuna, H.B.; Moon, J.-H.; Choi, S.-I.; Lim, H.-I.; Ahn, Y.S. Antifungal Activity of Volatile Organic Compounds from Bacillus velezensis CE 100 against Colletotrichum gloeosporioides. Horticulturae 2022, 8, 557. https://doi.org/10.3390/horticulturae8060557
Choub V, Won S-J, Ajuna HB, Moon J-H, Choi S-I, Lim H-I, Ahn YS. Antifungal Activity of Volatile Organic Compounds from Bacillus velezensis CE 100 against Colletotrichum gloeosporioides. Horticulturae. 2022; 8(6):557. https://doi.org/10.3390/horticulturae8060557
Chicago/Turabian StyleChoub, Vantha, Sang-Jae Won, Henry B. Ajuna, Jae-Hyun Moon, Su-In Choi, Hyo-In Lim, and Young Sang Ahn. 2022. "Antifungal Activity of Volatile Organic Compounds from Bacillus velezensis CE 100 against Colletotrichum gloeosporioides" Horticulturae 8, no. 6: 557. https://doi.org/10.3390/horticulturae8060557