
Trunk Injection as a Tool to Deliver Plant Protection Materials—An Overview of Basic Principles and Practical Considerations


Abstract
:1. Introduction
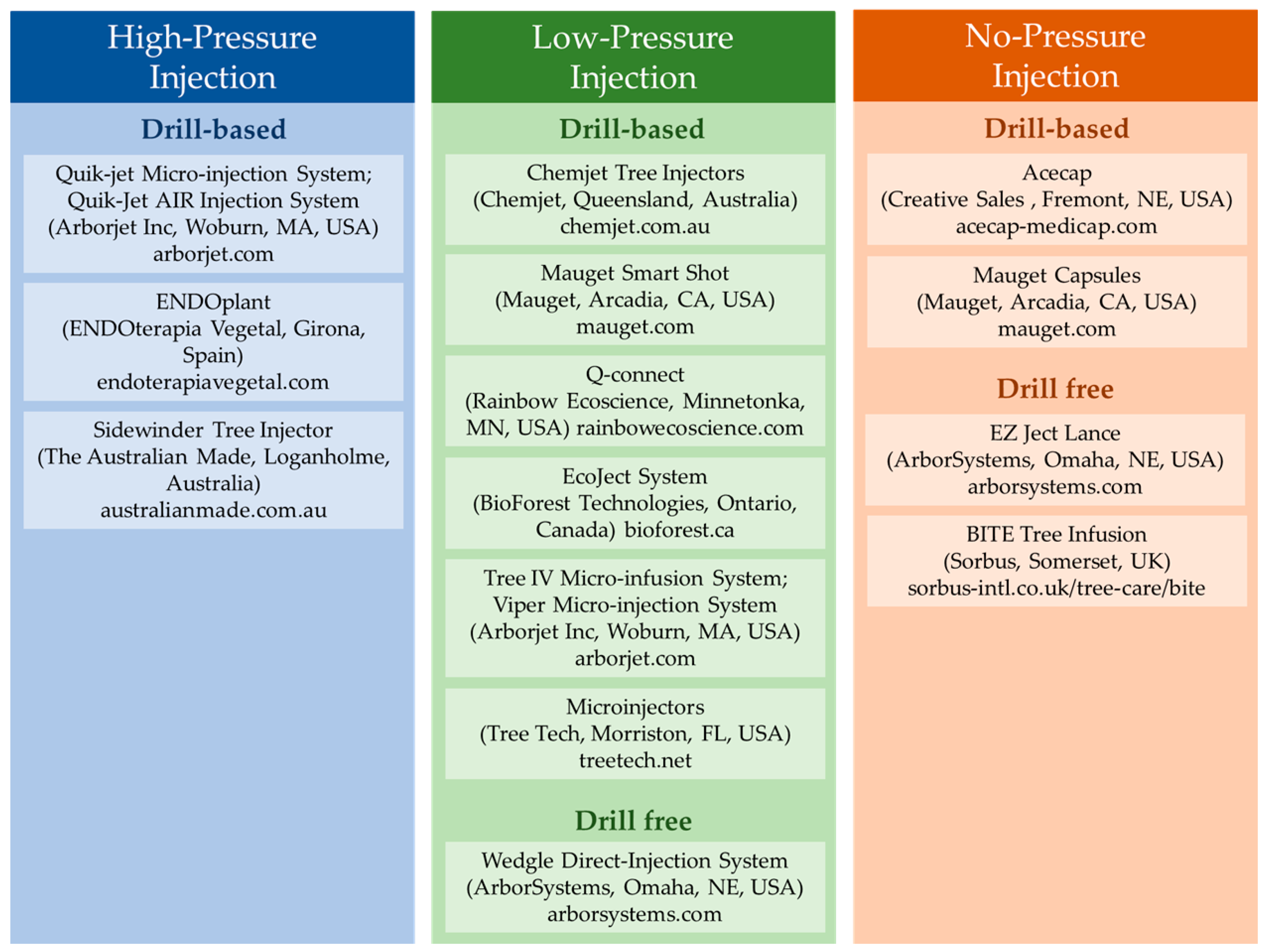
2. Trunk Injection Methods
2.1. High-Pressure Injection
2.2. Low-Pressure Injection
2.3. No-Pressure Injection
3. Physiological Principles of Trunk Injection
3.1. Tree Anatomy
3.2. Cohesion-Tension Theory
3.3. CODIT
4. Physiological Implications of Trunk Injection
4.1. Wound Size: Drilled-Based vs. No-Drill Injection
4.2. Pressure: High-Pressure vs. Low- or No-Pressure Injection
5. Trunk Injection to Control Pests and Diseases
5.1. Fungal and Oomycete Diseases
5.2. Insect Pests and Nematodes
5.3. Bacterial and Phytoplasma Diseases
6. Other Considerations
6.1. Location of Injection
6.2. Xenobiotic Mobility
6.3. Timing of Injection
6.4. Environmental and Human Health Concerns
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sánchez Zamora, M.A.; Fernández Escobar, R. Injector-Size and the Time of Application Affects Uptake of Tree Trunk-Injected Solutions. Sci. Hortic. 2000, 84, 163–177. [Google Scholar] [CrossRef]
- Wise, J.C.; VanWoerkom, A.H.; Acimovic, S.G.; Sundin, G.W.; Cregg, B.M.; Vandervoort, C.V. Trunk Injection: A Discriminating Delivering System for Horticulture Crop IPM. Entomol. Ornithol. Herpetol. Curr. Res. 2014, 3, 1. [Google Scholar] [CrossRef]
- Roach, W.A. Plant Injection as a Physiological Method. Ann. Bot. 1939, 3, 155–277. [Google Scholar] [CrossRef]
- Perry, T.; Santamour, F.; Stipes, R.; Shear, T.; Shigo, A. Exploring Alternatives To Tree Injection. J. Arboric. 1991, 17, 217–226. [Google Scholar] [CrossRef]
- Osterbauer, N.K.; French, D.W. Propiconazole as a Treatment for Oak Wilt in Quercus rubra and Q. ellipsoidalis. J. Agric. Sci. Technol. A 1992, 18, 221–226. [Google Scholar] [CrossRef]
- Darvas, J.M.; Torien, J.C.; Milne, D.L. Injection of Established Avocado Trees for the Effective Control of Phytophthora Root Rot. Calif. Avocado Soc. 1983, 67, 76–77. [Google Scholar]
- Stennes, M.A.; French, D.W. Distribution and Retention of Thiabendazole Hypophosphite and Carbendazim Phosphate Injected into Mature American Elms. Phytopathology 1987, 77, 707–712. [Google Scholar] [CrossRef]
- Haugen, B.L.; Stennes, M. Fungicide Injection to Control Dutch Elm Disease: Understanding the Options. Plant Dis. Q. 1999, 20, 29–38. [Google Scholar]
- Fernandez-Escobar, R.; Gallego, F.J.; Benlloch, M.; Membrillo, J.; Infante, J.; Perez De Algaba, A. Treatment of Oak Decline Using Pressurized Injection Capsules of Antifungal Materials. Eur. J. For. Pathol. 1999, 29, 29–38. [Google Scholar] [CrossRef]
- Filer, T.H. Pressure Apparatus for Injecting Chemicals Into Trees. Plant Dis. Rep. 1973, 57, 338–341. [Google Scholar]
- Holmes, F.W. Distribution of Dye in Elms after Trunk or Root Injection. J. Arboric. 1982, 8, 250–252. [Google Scholar]
- Kozlowski, T.T.; Winget, C.H. Patterns of Water Movement in Forest Trees. Bot. Gaz. 1963, 124, 301–311. [Google Scholar] [CrossRef]
- Sachs, R.M.; Nyland, G.; Hackett, W.P.; Coffelt, J.; Debie, J.; Giannini, G. Pressurized Injection of Aqueous Solutions into Tree Trunks. Sci. Hortic. 1977, 6, 297–310. [Google Scholar] [CrossRef]
- Shigo, A.L.; Marx, H.G. Compartmentalization of Decay in Trees; USDA Forest Service Agency Information Bulletin; USDA: Washington, DC, USA, 1977; pp. 1–73.
- Santamour, F.S. Wound Compartmentalization in Cultivars of Acer, Gleditsia, and Other Genera. J. Environ. Hortic. 1984, 2, 123–125. [Google Scholar] [CrossRef]
- Stack, R.W. Effect of Tree Size, Hole Location and Wetwood Fluxing on Healing of Injection Wounds in American Elms. J. Arboric. 1985, 11, 45–47. [Google Scholar]
- Smith, K.T. Wounding, Compartmentalization, and Treatment Tradeoffs. J. Arboric. 1988, 14, 226–229. [Google Scholar]
- Neely, D. Tree Wound Closure. J. Arboric. 1988, 14, 148–152. [Google Scholar]
- Costinis, A.C. The Wounding Effects of Mauget and Creative Sales Injections. J. Arboric. 1980, 6, 204–208. [Google Scholar]
- Sinclair, W.A.; Larsen, A.O. Wood Characteristics Related to “Injectability” of Trees. J. Arboric. 1980, 7, 6–10. [Google Scholar]
- Chaney, W.R. Anatomy and Physiology Related to Chemical Movement in Trees. J. Arboric. 1985, 12, 85–91. [Google Scholar]
- Doccola, J.J.; Smitley, D.R.; Davis, T.W.; Aiken, J.J.; Wild, P.M. Tree Wound Responses Following Systemic Insecticide Trunk Injection Treatments in Green Ash (Fraxinus pennsylvanica Marsh.) as Determined by Destructive Autopsy. Arboric. Urban For. 2011, 37, 6–12. [Google Scholar] [CrossRef]
- Aćimović, S.G.; Cregg, B.M.; Sundin, G.W.; Wise, J.C. Comparison of Drill- and Needle-Based Tree Injection Technologies in Healing of Trunk Injection Ports on Apple Trees. Urban For. Urban Green. 2016, 19, 151–157. [Google Scholar] [CrossRef]
- Tattar, T.; Tattar, S. Evidence for the Downward Movement of Materials Injected into Trees. Arboric. Urban For. 1999, 25, 325–332. [Google Scholar] [CrossRef]
- Ford, C.R.; Vose, J.M.; Daley, M.; Phillips, N. Use of Water by Eastern Hemlock: Implications for Systemic Insecticide Application. Arboric. Urban For. 2007, 33, 421–427. [Google Scholar] [CrossRef]
- Shang, Q.; Liao, K.; Liu, H.; Zhao, B. Study on Structure of Needle Head and Seal Mechanism of Tree Trunk Injection. In Proceedings of the 2011 International Conference on Transportation, Mechanical, and Electrical Engineering, TMEE 2011, Changchun, China, 16–18 December 2011; pp. 813–816. [Google Scholar]
- Montecchio, L. A Venturi Effect Can Help Cure Our Trees. J. Vis. Exp. 2013, 80, e51199. [Google Scholar] [CrossRef]
- Dalakouras, A.; Jarausch, W.; Buchholz, G.; Bassler, A.; Braun, M.; Manthey, T.; Krczal, G.; Wassenegger, M. Delivery of Hairpin Rnas and Small Rnas into Woody and Herbaceous Plants by Trunk Injection and Petiole Absorption. Front. Plant Sci. 2018, 9, 1253. [Google Scholar] [CrossRef]
- Killiny, N.; Gonzalez-Blanco, P.; Santos-Ortega, Y.; Al-Rimawi, F.; Levy, A.; Hijaz, F.; Albrecht, U.; Batuman, O. Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin. Antibiotics 2019, 8, 262. [Google Scholar] [CrossRef]
- Kuroda, K.; Yamane, K.; Itoh, Y. In Planta Analysis of the Radial Movement of Minerals from inside to Outside in the Trunks of Standing Japanese Cedar (Cryptomeria japonica D. Don) Trees at the Cellular Level. Forests 2021, 12, 251. [Google Scholar] [CrossRef]
- Kuroda, K.; Yamane, K.; Itoh, Y. Cellular Level in Planta Analysis of Radial Movement of Artificially Injected Caesium in Cryptomeria japonica Xylem. Trees Struct. Funct. 2018, 32, 1505–1517. [Google Scholar] [CrossRef]
- Berger, C.; Laurent, F. Trunk Injection of Plant Protection Products to Protect Trees from Pests and Diseases. Crop Prot. 2019, 124, 104831. [Google Scholar] [CrossRef]
- Tsen, E.W.J.; Sitzia, T.; Webber, B.L. To Core, or Not to Core: The Impact of Coring on Tree Health and a Best-Practice Framework for Collecting Dendrochronological Information from Living Trees. Biol. Rev. 2016, 91, 899–924. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, P.A.; Ali, S.; Braun, G.; Bevis, E.; Fillmore, S. Reducing Apple Scab and Frogeye or Black Rot Infections with Salicylic Acid or Its Analogue on Field-Established Apple Trees. Can. J. Plant Pathol. 2019, 41, 345–354. [Google Scholar] [CrossRef]
- Pegg, K.G. Trunk Injection Methodology. Australas. Plant Pathol. 1990, 19, 142–143. [Google Scholar] [CrossRef]
- Aćimović, S.G.; Martin, D.K.H.; Turcotte, R.M.; Meredith, C.L.; Munck, I.A. Choosing an Adequate Pesticide Delivery System for Managing Pathogens with Difficult Biologies: Case Studies on Diplodia corticola, Venturia inaequalis and Erwinia amylovora. In Plant Diseases—Current Threats and Management Trends; IntechOpen: London, UK, 2020. [Google Scholar]
- Hunt, P.; Dabek, A.J.; Schuilin, M. Remission of Symptoms Following Tetracycline Treatment of Lethal Yellowing-Infected Coconut Palms. Phytopathology 1974, 64, 307–312. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Mencuccini, M.; Álvarez, X.; Camacho, J.; Loepfe, L.; Piñol, J. Spatial Distribution and Packing of Xylem Conduits. Am. J. Bot. 2012, 99, 1189–1196. [Google Scholar] [CrossRef]
- Cartenì, F.; Deslauriers, A.; Rossi, S.; Morin, H.; de Micco, V.; Mazzoleni, S.; Giannino, F. The Physiological Mechanisms behind the Earlywood-to-Latewood Transition: A Process-Based Modeling Approach. Front. Plant Sci. 2018, 9, 1053. [Google Scholar] [CrossRef]
- Ellmore, G.S.; Zanne, A.E.; Orians, C.M. Comparative Sectoriality in Temperate Hardwoods: Hydraulics and Xylem Anatomy. Bot. J. Linn. Soc. 2006, 150, 61–71. [Google Scholar] [CrossRef]
- Zanne, A.E.; Sweeney, K.; Sharma, M.; Orians, C.M. Patterns and Consequences of Differential Vascular Sectoriality in 18 Temperate Tree and Shrub Species. Funct. Ecol. 2006, 20, 200–206. [Google Scholar] [CrossRef]
- Zanne, A.E.; Westoby, M.; Falster, D.S.; Ackerly, D.D.; Loarie, S.R.; Arnold, S.E.J.; Coomes, D.A. Angiosperm Wood Structure: Global Patterns in Vessel Anatomy and Their Relation to Wood Density and Potential Conductivity. Am. J. Bot. 2010, 97, 207–215. [Google Scholar] [CrossRef]
- Nadezhdina, N. Integration of Water Transport Pathways in a Maple Tree: Responses of Sap Flow to Branch Severing. Ann. For. Sci. 2010, 67, 107. [Google Scholar] [CrossRef]
- Thorn, A.M.; Orians, C.M. Modeling the Influence of Differential Sectoriality on the Photosynthetic Responses of Understory Saplings to Patchy Light and Water Availability. Trees Struct. Funct. 2011, 25, 833. [Google Scholar] [CrossRef]
- Tanis, S.R.; Cregg, B.M.; Mota-Sanchez, D.; McCullough, D.G.; Poland, T.M. Spatial and Temporal Distribution of Trunk-Injected 14C-Imidacloprid in Fraxinus Trees. Pest Manag. Sci. 2012, 68, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Ewers, F.W.; Ewers, J.M.; Jacobsen, A.L.; López-Portillo, J. Vessel Redundancy: Modeling Safety in Numbers. IAWA J. 2007, 28, 373–388. [Google Scholar] [CrossRef]
- Schenk, H.J.; Espino, S.; Goedhart, C.M.; Nordenstahl, M.; Cabrera, H.I.M.; Jones, C.S. Hydraulic Integration and Shrub Growth Form Linked across Continental Aridity Gradients. Proc. Natl. Acad. Sci. USA 2008, 105, 11248–11253. [Google Scholar] [CrossRef]
- Dixon, H.H.; Joly, J. XII. On the Ascent of Sap. Philos. Trans. R. Soc. Lond. 1895, 186, 563–576. [Google Scholar] [CrossRef]
- Pickard, W.F. The Ascent of Sap in Plants. Prog. Biophys. Mol. Biol. 1981, 37, 181–229. [Google Scholar] [CrossRef]
- Kim, H.K.; Park, J.; Hwang, I. Investigating Water Transport through the Xylem Network in Vascular Plants. J. Exp. Bot. 2014, 65, 1895–1904. [Google Scholar] [CrossRef]
- Zimmermann, U.; Schneider, H.; Wegner, L.H.; Haase, A. Water Ascent in Tall Trees: Does Evolution of Land Plants Rely on a Highly Metastable State? New Phytol. 2004, 162, 575–615. [Google Scholar] [CrossRef]
- Pereira, L.; Jansen, S.; Miranda, M.T.; Pacheco, V.S.; Kaack, L.; Pires, G.S.; Guan, X.; Mayer, J.L.S.; Machado, E.C.; Schenk, H.J.; et al. Dynamic Changes in Gas Solubility of Xylem Sap Reiterate the Enigma of Plant Water Transport under Negative Pressure. bioRxiv 2022. [Google Scholar]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant Xylem Hydraulics: What We Understand, Current Research, and Future Challenges. J. Integr. Plant Biol. 2017, 59, 356–389. [Google Scholar] [CrossRef]
- Jacobsen, A.L.; Pratt, B.R.; Tobin, M.F.; Hacke, U.G.; Ewers, F.W. A Global Analysis of Xylem Vessel Length in Woody Plants. Am. J. Bot. 2012, 99, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Stroock, A.D.; Pagay, V.V.; Zwieniecki, M.A.; Holbrook, N.M. The Physicochemical Hydrodynamics of Vascular Plants. Annu. Rev. Fluid Mech. 2014, 46, 615–642. [Google Scholar] [CrossRef]
- Nagy, N.E.; Norli, H.R.; Fongen, M.; Østby, R.B.; Heldal, I.M.; Davik, J.; Hietala, A.M. Patterns and Roles of Lignan and Terpenoid Accumulation in the Reaction Zone Compartmentalizing Pathogen-Infected Heartwood of Norway Spruce. Planta 2022, 255, 63. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, A.; Planas-Marquès, M.; Capellades, M.; Valls, M.; Coll, N.S. Blocking Intruders: Inducible Physico-Chemical Barriers against Plant Vascular Wilt Pathogens. J. Exp. Bot. 2021, 72, 184–198. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Roddy, A.B.; Wason, J.W.; McElrone, A.J. Functional Status of Xylem Through Time. Annu. Rev. Plant Biol. 2019, 70, 407–433. [Google Scholar] [CrossRef]
- Pouzoulet, J.; Rolshausen, P.E.; Charbois, R.; Chen, J.; Guillaumie, S.; Ollat, N.; Gambetta, G.A.; Delmas, C.E.L. Behind the Curtain of the Compartmentalization Process: Exploring How Xylem Vessel Diameter Impacts Vascular Pathogen Resistance. Plant Cell Environ. 2020, 43, 2782–2796. [Google Scholar] [CrossRef]
- Morris, H.; Brodersen, C.; Schwarze, F.W.M.R.; Jansen, S. The Parenchyma of Secondary Xylem and Its Critical Role in Tree Defense against Fungal Decay in Relation to the CODIT Model. Front. Plant Sci. 2016, 7, 1665. [Google Scholar] [CrossRef]
- Morris, H.; Hietala, A.M.; Jansen, S.; Ribera, J.; Rosner, S.; Salmeia, K.A.; Schwarze, F.W.M.R. Using the CODIT Model to Explain Secondary Metabolites of Xylem in Defence Systems of Temperate Trees against Decay Fungi. Ann. Bot. 2020, 125, 701–720. [Google Scholar] [CrossRef]
- Pearce, R.B. Reaction Zone Relics and the Dynamics of Fungal Spread in the Xylem of Woody Angiosperms. Physiol. Mol. Plant Pathol. 1991, 39, 41–55. [Google Scholar] [CrossRef]
- Schmitt, U.; Liese, W. Response of Xylem Parenchyma by Suberization in Some Hardwoods after Mechanical Injury. Trees 1993, 8, 23–30. [Google Scholar] [CrossRef]
- Robbertse, P.J.; Duvenhage, J.A. Reaction of Avocado Wood on Injections into the Trunk. South Afr. Avacado Grow. Assoc. Yearb. 1999, 22, 39–47. [Google Scholar]
- Smith, K.T.; Lewis, P.A. Potential Concerns for Tree Wound Response from Stem Injection. In Proceedings of the 3rd Symposium on Hemlock Wooley Adelgid Conference, Asheville, NC, USA, 1–3 February 2005; pp. 173–178. [Google Scholar]
- Biggs, A.R. Suberized Boundary Zones and the Chronology of Wound Response in Tree Bark. Phytopathology 1985, 75, 1191–1195. [Google Scholar] [CrossRef]
- Eyles, A.; Davies, N.W.; Mohammed, C. Novel Detection of Formylated Phloroglucinol Compounds (FPCs) in the Wound Wood of Eucalyptus Globulus and E. Nitens. J. Chem. Ecol. 2003, 29, 881–898. [Google Scholar] [CrossRef] [PubMed]
- Shigo, A.L.; Money, W.E.; Dodds, D.I. Some Internal Effects of Mauget Tree Injections. J. Arboric. 1977, 3, 213–220. [Google Scholar] [CrossRef]
- Shigo, A.L.; Shortle, W.C. Compartmentalization of Discolored Wood in Heartwood of Red Oak. Phytopathology 1979, 69, 710–711. [Google Scholar] [CrossRef]
- Vasconcellos, L.A.B.C.; Castle, W.S. Trunk Xylem Anatomy of Mature Healthy and Blighted Grapefruit Trees on Several Rootstocks. J. Am. Soc. Hortic. Sci. 1994, 119, 185–194. [Google Scholar] [CrossRef]
- Dujesiefken, D.; Liese, W. The CODIT Principle: Implications for Best Practices; Martin One Source: Champaign, IL, USA, 2015. [Google Scholar]
- James, R.; Tisserat, N.; Todd, T. Prevention of Pine Wilt of Scots Pine (Pinus sylvestris) with Systemic Abamectin Injections. Arboric. Urban For. 2006, 32, 195–201. [Google Scholar] [CrossRef]
- Reil, W. Pressure-Injecting Chemicals in to Trees. Calif. Agric. 1979, 33, 16–19. [Google Scholar]
- Martín, J.A.; Sobrino-Plata, J.; Rodríguez-Calcerrada, J.; Collada, C.; Gil, L. Breeding and Scientific Advances in the Fight against Dutch Elm Disease: Will They Allow the Use of Elms in Forest Restoration? New For. 2019, 50, 183–215. [Google Scholar] [CrossRef]
- D’Arcy, C.J. Dutch elm disease. Am. Phytopathol. Soc. 1981, 245, 56–67. [Google Scholar] [CrossRef]
- Marcotrigiano, M. Elms Revisited. Arboric. Urban For. 2017, 43, 217–241. [Google Scholar] [CrossRef]
- Brazee, N.J.; Marra, R.E. Incidence of Internal Decay in American Elms (Ulmus americana) under Regular Fungicide Injection to Manage Dutch Elm Disease. Arboric. Urban For. 2020, 46, 1–11. [Google Scholar] [CrossRef]
- Postma, J.; Goossen-van de Geijn, H. Twenty-Four Years of Dutch Trig® Application to Control Dutch Elm Disease. BioControl 2016, 61, 305–312. [Google Scholar] [CrossRef]
- Stipes, R.J. The Management of Dutch Elm Disease. In The Elms; Springer: Boston, MA, USA, 2000; pp. 157–172. [Google Scholar]
- Schutte, G.C.; Botha, T.; Bezuidenhout, J.J.; Kotze, J.M. Distribution of Phosphite in Avocado Trees after Trunk Injection with Phosphorous Acid and Its Possible Response to Phytophthora cinnamomi. South Afr. Avocado Grow. Assoc. Yearb. 1988, 12, 32–34. [Google Scholar]
- Ramírez-Gil, J.G.; Castañeda-Sánchez, D.A.; Morales-Osorio, J.G. Production of Avocado Trees Infected with Phytophthora cinnamomi under Different Management Regimes. Plant Pathol. 2017, 66, 623–632. [Google Scholar] [CrossRef]
- Dann, E.K.; Le, D.P. Effects of Silicon Amendment on Soilborne and Fruit Diseases of Avocado. Plants 2017, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.H.; Montas, W.; Evans, E.A.; Olszack, R. How to Make a Simple and Inexpensive Passive and Pressurized Infusion System for Systemically Applied Pest Control Substances to Fruit Trees. Proc. Fla. State Hortic. Soc. 2014, 127, 6–9. [Google Scholar]
- De Beer, Z.W.; Procter, M.; Wingfield, M.J.; Marincowitz, S.; Duong, T.A. Generic Boundaries in the Ophiostomatales Reconsidered and Revised. In Studies in Mycology; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2022; pp. 57–120. [Google Scholar] [CrossRef]
- Fraedrich, S.W.; Harrington, T.C.; Rabaglia, R.J.; Ulyshen, M.D.; Mayfield, A.E.; Hanula, J.L.; Eickwort, J.M.; Miller, D.R. A Fungal Symbiont of the Redbay Ambrosia Beetle Causes a Lethal Wilt in Redbay and Other Lauraceae in the Southeastern United States. Plant Dis. 2008, 92, 215–224. [Google Scholar] [CrossRef]
- Harrington, T.C.; Fraedrich, S.W.; Achayeva, D.N. Raffaelea Lauricola, a New Ambrosia Beetle Symbiont and Pathogen on the Lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Carrillo, D.; Duncan, R.E.; Ploetz, J.N.; Campbell, A.F.; Ploetz, R.C.; Peña, J.E. Lateral Transfer of a Phytopathogenic Symbiont among Native and Exotic Ambrosia Beetles. Plant Pathol. 2014, 63, 54–62. [Google Scholar] [CrossRef]
- Cruz, L.F.; Menocal, O.; Kendra, P.E.; Carrillo, D. Phoretic and Internal Transport of Raffaelea lauricola by Different Species of Ambrosia Beetle Associated with Avocado Trees. Symbiosis 2021, 84, 151–161. [Google Scholar] [CrossRef]
- Dann, E.K.; Ploetz, R.C.; Coates, L.M.; Pegg, K.G. Foliar, Fruit and Soilborne Diseases. In The Avocado: Botany, Production and Uses; CABI: Wallingford, UK, 2013; pp. 380–422. [Google Scholar]
- Crane, J.H.; Wasielweski, J.; Carrillo, D.; Gazis, R.; Schaffer, B.; Ballen, F.; Evans, E. Recommendations for the Detection and Mitigation of Laurel Wilt Disease in Avocado and Related Tree Species in the Home Landscape: HS1358, 2/2020; EDIS, University of Florida: Gainesville, FL, USA, 2020. [Google Scholar] [CrossRef]
- Crane, J.H.; Carrillo, D.; Evans, E.A.; Gazis, R.; Schaffer, B.; Ballen, F.; Wasielweski, J. Current Recommendations for Control of Laurel Wilt and Its Ambrosia Beetle Vectors in Florida’s Commercial Avocado Groves. Proc. Fla. State Hortic. Soc. 2020, 133, 4–6. [Google Scholar]
- Crane, J.H.; Ploetz, R.C.; White, T.; Krogstad, G.C.; Prosser, T.; Konkol, J.; Wideman, R. Efficacy of Three Microinfused Fungicides to Control Laurel Wilt on Avocado in Martin and Brevard Counties. Proc. Fla. State Hortic. Soc. 2015, 128, 58–60. [Google Scholar]
- Dal Maso, E.; Cocking, J.; Montecchio, L. An Enhanced Trunk Injection Formulation of Potassium Phosphite against Chestnut Ink Disease. Arboric. J. 2017, 39, 124–141. [Google Scholar] [CrossRef]
- Dal Maso, E.; Linaldeddu, B.T.; Fanchin, G.; Faccoli, M.; Montecchio, L. The Potential for Pesticide Trunk Injections for Control of Thousand Cankers Disease of Walnut. Phytopathol. Mediterr. 2019, 58, 73–79. [Google Scholar] [CrossRef]
- González, M.; Romero, M.Á.; Serrano, M.S.; Sánchez, M.E. Fosetyl-Aluminium Injection Controls Root Rot Disease Affecting Quercus Suber in Southern Spain. Eur. J. Plant Pathol. 2020, 156, 101–109. [Google Scholar] [CrossRef]
- Romero, M.A.; González, M.; Serrano, M.S.; Sánchez, M.E. Trunk Injection of Fosetyl-Aluminium Controls the Root Disease Caused by Phytophthora cinnamomi on Quercus ilex Woodlands. Ann. Appl. Biol. 2019, 174, 313–318. [Google Scholar] [CrossRef]
- Fischer, J.; Beckers, S.J.; Yiamsawas, D.; Thines, E.; Landfester, K.; Wurm, F.R. Targeted Drug Delivery in Plants: Enzyme-Responsive Lignin Nanocarriers for the Curative Treatment of the Worldwide Grapevine Trunk Disease Esca. Adv. Sci. 2019, 6, 1802315. [Google Scholar] [CrossRef]
- Düker, A.; Kubiak, R. Stem Injection of Triazoles for the Protection of Vitis vinifera L. (‘Riesling’) against Powdery Mildew (Uncinula necator). Vitis J. Grapevine Res. 2011, 50, 73–79. [Google Scholar]
- Masikane, S.L.; Novela, P.; Mohale, P.; McLeod, A. Effect of Phosphonate Application Timing and -Strategy on Phosphite Fruit and Root Residues of Avocado. Crop Prot. 2020, 128, 105008. [Google Scholar] [CrossRef]
- McLeod, A.; Masikane, S.L.; Novela, P.; Ma, J.; Mohale, P.; Nyoni, M.; Stander, M.; Wessels, J.P.B.; Pieterse, P. Quantification of Root Phosphite Concentrations for Evaluating the Potential of Foliar Phosphonate Sprays for the Management of Avocado Root Rot. Crop Prot. 2018, 103, 87–97. [Google Scholar] [CrossRef]
- Mule, R.; Fodale, A.S.; Tucci, A. Control of Olive Verticillium Wilt by Trunk Injection with Different Doses of Fosetyl Al and Benomyl. Acta Hortic. 2002, 586, 761–764. [Google Scholar] [CrossRef]
- Tsror, L.; Bar-On, N.; Tugendhaft, Y.; Lavee, S. Attempts to Control Verticillium Wilt on Olives in Israel. In Proceedings of the Olivebioteq, Marsala, Italy, 5–10 November 2006; pp. 235–238. [Google Scholar]
- Tattar, T.A.; Dotson, J.A.; Ruizzo, M.S.; Steward, V.B. Translocation of Imidacloprid in Three Tree Species When Trunk- and Soil-Injected. J. Arboric. 1998, 24, 54–56. [Google Scholar] [CrossRef]
- Cowles, R.S.; Montgomery, M.E.; Cheah, C.A.S.-J. Activity and Residues of Imidacloprid Applied to Soil and Tree Trunks to Control Hemlock Woolly Adelgid (Hemiptera: Adelgidae) in Forests. J. Econ. Entomol. 2006, 99, 1258–1267. [Google Scholar] [CrossRef]
- Mota-Sanchez, D.; Cregg, B.M.; McCullough, D.G.; Poland, T.M.; Hollingworth, R.M. Distribution of Trunk-Injected 14C-Imidacloprid in Ash Trees and Effects on Emerald Ash Borer (Coleoptera: Buprestidae) Adults. Crop Prot. 2009, 28, 655–661. [Google Scholar] [CrossRef]
- Doccola, J.J.; Wild, P.M. Tree Injection as an Alternative Method of Insecticide Application. In Insecticides—Basic and Other Applications; Soloneski, S., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Smitley, D.R.; Doccola, J.J.; Cox, D.L. Multiple-Year Protection of Ash Trees from Emerald Ash Borer with a Single Trunk Injection of Emamectin Benzoate, and Single-Year Protection with an Imidacloprid Basal Drench. Arboric. Urban For. 2010, 36, 206–211. [Google Scholar] [CrossRef]
- McCullough, D.G.; Poland, T.M.; Tluczek, A.R.; Anulewicz, A.; Wieferich, J.; Siegert, N.W. Emerald Ash Borer (Coleoptera: Buprestidae) Densities over a 6-Yr Period on Untreated Trees and Trees Treated with Systemic Insecticides at 1-, 2-, and 3-Yr Intervals in a Central Michigan Forest. J. Econ. Entomol. 2019, 112, 201–212. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer Invasion of North America: History, Biology, Ecology, Impacts, and Management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef]
- Lee, S.C.; Lee, H.R.; Kim, D.S.; Kwon, J.H.; Huh, M.J.; Park, I.K. Emamectin Benzoate 9.7% SL as a New Formulation for a Trunk-Injections against Pine Wood Nematode, Bursaphelenchus xylophilus. J. For. Res. 2020, 31, 1399–1403. [Google Scholar] [CrossRef]
- Sousa, E.; Naves, P.; Vieira, M. Prevention of Pine Wilt Disease Induced by Bursaphelenchus xylophilus and Monochamus galloprovincialis by Trunk Injection of Emamectin Benzoate. Phytoparasitica 2012, 41, 143–148. [Google Scholar] [CrossRef]
- Yang, Z.; ZhengYun, L.; DingChao, Y.; FeiFei, H.; LinPing, Z.; Dong, L. Screening and Prevention Effect of New and Effective Pesticides against Pine Wilt Nematode Bursaphelenchus xylophilus. J. Plant Prot. 2017, 44, 856–862. [Google Scholar]
- Cha, D.J.; Kim, J.; Kim, D.S. Nematicidal Activities of Three Naphthoquinones against the Pine Wood Nematode, Bursaphelenchus xylophilus. Molecules 2019, 24, 3634. [Google Scholar] [CrossRef] [PubMed]
- Coslor, C.C.; Sundin, G.W.; Wise, J.C. The Efficacy of Trunk Injections of Emamectin Benzoate and Phosphorous Acid for Control of Obliquebanded Leafroller and Apple Scab on Semi-Dwarf Apple. Crop Prot. 2019, 118, 44–49. [Google Scholar] [CrossRef]
- Coslor, C.C.; Vandervoort, C.; Wise, J.C. Insecticide Dose and Seasonal Timing of Trunk Injection in Apples Influence Efficacy and Residues in Nectar and Plant Parts. Pest Manag. Sci. 2019, 75, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, R.M.; Lagalante, A.; Jones, J.; Cook, F.; Elliott, T.; Billings, A.A.; Park, Y.L. Spatial and Temporal Distribution of Imidacloprid within the Crown of Eastern Hemlock. J. Insect Sci. 2017, 17, 1–7. [Google Scholar] [CrossRef]
- Coleman, T.W.; Smith, S.L.; Jones, M.I.; Graves, A.D.; Strom, B.L. Efficacy of Systemic Insecticides for Control of the Invasive Goldspotted Oak Borer (Coleoptera: Buprestidae) in California. J. Econ. Entomol. 2017, 110, 2129–2139. [Google Scholar] [CrossRef]
- Bhandari, B.P.; Cheng, Z. Trunk Injection of Systemic Insecticides to Control Stem and Leaf Gall Wasps, Josephiella Species (Hymenoptera: Agaonidae), on Chinese Banyan (Rosales: Moraceae) in Hawaii. Fla. Entomol. 2016, 99, 172–177. [Google Scholar] [CrossRef]
- Wang, J.H.; Che, S.C.; Qiu, L.F.; Li, G.; Shao, J.L.; Zhong, L.; Zhang, G.F.; Xu, H. Efficacy of Emamectin Benzoate Trunk Injection against the Asian Long-Horned Beetle [Anoplophora glabripennis (Coleoptera: Cerambycidae)]. J. Econ. Entomol. 2020, 113, 340–347. [Google Scholar] [CrossRef]
- Bhandari, B.P.; Cheng, Z. Lobate Lac Scale, Paratachardina Pseudolobata (Hemiptera: Keriidae), in Hawaii’s Urban Landscape: Hosts and Management. Int. J. Trop. Insect Sci. 2018, 38, 71–76. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, J.; Li, Y.; Li, J.; Shi, X.H. Evaluation of the Effectiveness of Insecticide Trunk Injections for Control of Latoia Lepida (Cramer) in the Sweet Olive Tree Osmanthus fragrans. PeerJ 2016, 4, e2480. [Google Scholar] [CrossRef]
- Ali, A.D.; Caldwell, D.L. Management of Staining and Galling Associated with Oxhorn Bucida Trees in Florida. Fla. Entomol. 2017, 100, 602–606. [Google Scholar] [CrossRef]
- Jagiełło, R.; Walczak, U.; Iszkuło, G.; Karolewski, P.; Baraniak, E.; Giertych, M.J. Impact of Cameraria Ohridella on Aesculus hippocastanum Growth and Long-Term Effects of Trunk Injection with Pesticides. Int. J. Pest Manag. 2019, 65, 33–43. [Google Scholar] [CrossRef]
- Gubka, A.; Zubrik, M.; Rell, S.; Gareau, N.; Goble, T.; Nikolov, C.; Galko, J.; Vakula, J.; Kunca, A.; Dejonge, R. The Effectiveness of the Neem Product TreeAzin® in Controlling Cameraria ohridella (Lepidoptera: Gracillariidae: Lithocolletinae). Eur. J. Entomol. 2020, 117, 463–473. [Google Scholar] [CrossRef]
- Grosman, D.M.; Eskalen, A.; Brownie, C. Evaluation of Emamectin Benzoate and Propiconazole for Management of a New Invasive Shot Hole Borer (Euwallacea Nr. fornicatus, Coleoptera: Curculionidae) and Symbiotic Fungi in California Sycamores. J. Econ. Entomol. 2019, 112, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E.; Kabashima, J.; Eskalen, A.; Dimson, M.; Mayorquin, J.S.; Carrillo, J.D.; Hanlon, C.C.; Paine, T.D. Evaluations of Insecticides and Fungicides for Reducing Attack Rates of a New Invasive Ambrosia Beetle (Euwallacea Sp., Coleoptera: Curculionidae: Scolytinae) in Infested Landscape Trees in California. J. Econ. Entomol. 2017, 110, 1611–1618. [Google Scholar] [CrossRef]
- Byrne, F.J.; Almanzor, J.; Tellez, I.; Eskalen, A.; Grosman, D.M.; Morse, J.G. Evaluation of Trunk-Injected Emamectin Benzoate as a Potential Management Strategy for Kuroshio Shot Hole Borer in Avocado Trees. Crop Prot. 2020, 132, 105136. [Google Scholar] [CrossRef]
- Wheeler, C.E.; Vandervoort, C.; Wise, J.C. Organic Control of Pear Psylla in Pear with Trunk Injection. Insects 2020, 11, 650. [Google Scholar] [CrossRef]
- Monteys, V.I.S.; Ribes, A.C.; Savin, I. The Invasive Longhorn Beetle Xylotrechus chinensis, Pest of Mulberries, in Europe: Study on Its Local Spread and Efficacy of Abamectin Control. PLoS ONE 2021, 16, e0245527. [Google Scholar] [CrossRef]
- Mokhtaryan, A.; Sheikhigarjan, A.; Arbab, A.; Mohammadipour, A.; Ardestanirostami, H. The Efficiency of Systemic Insecticides and Complete Fertilizer by Trunk Injection Method against Leopard Moth in Infested Walnut Trees. J. Basic Appl. Zool. 2021, 82, 55. [Google Scholar] [CrossRef]
- Kiss, M.; Hachoumi, I.; Nagy, V.; Ladányi, M.; Gutermuth, Á.; Szabó, Á.; Sörös, C. Preliminary Results about the Efficacy of Abamectin Trunk Injection against the Walnut Husk Fly (Rhagoletis completa). J. Plant Dis. Prot. 2021, 128, 333–338. [Google Scholar] [CrossRef]
- Fu, B.; Qiu, H.; Li, Q.; Tang, L.; Zeng, D.; Liu, K.; Gao, Y. Flower Injection of Imidacloprid and Spirotetramat: A Novel Tool for the Management of Banana Thrips Thrips hawaiiensis. J. Pest Sci. 2020, 93, 1073–1084. [Google Scholar] [CrossRef]
- Chihaoui-Meridja, S.; Harbi, A.; Abbes, K.; Chaabane, H.; la Pergola, A.; Chermiti, B.; Suma, P. Systematicity, Persistence and Efficacy of Selected Insecticides Used in Endotherapy to Control the Red Palm Weevil Rhynchophorus ferrugineus (Olivier, 1790) on Phoenix canariensis. Phytoparasitica 2020, 48, 75–85. [Google Scholar] [CrossRef]
- Mashal, M.M.; Obeidat, B.F. The Efficacy Assessment of Emamectin Benzoate Using Micro Injection System to Control Red Palm Weevil. Heliyon 2019, 5, e01833. [Google Scholar] [CrossRef] [PubMed]
- Kou, H.; Sun, Y.; Dong, Z.; Zhang, Z. Comparison between Sustained Effects of Spray and Injection Thiamethoxam on Apple Aphids and Non-Target Insects in Apple Orchard. Ecotoxicol. Environ. Saf. 2021, 207, 111307. [Google Scholar] [CrossRef]
- Hu, J.; Wang, N. Evaluation of the Spatiotemporal Dynamics of Oxytetracycline and Its Control Effect against Citrus Huanglongbing via Trunk Injection. Phytopathology 2016, 106, 1495–1503. [Google Scholar] [CrossRef]
- Amanifar, N.; Taghavi, M.; Salehi, M. Xylella Fastidiosa from Almond in Iran: Overwinter Recovery and Effects of Antibiotics. Phytopathol. Mediterr. 2016, 55, 337–345. [Google Scholar] [CrossRef]
- Shin, K.; Ascunce, M.S.; Narouei-Khandan, H.A.; Sun, X.; Jones, D.; Kolawole, O.O.; Goss, E.M.; van Bruggen, A.H.C. Effects and Side Effects of Penicillin Injection in Huanglongbing Affected Grapefruit Trees. Crop Prot. 2016, 90, 106–116. [Google Scholar] [CrossRef]
- Norelli, J.L.; Jones, A.L.; Aldwinckle, H.S. Fire Blight Management in the Twenty-First Century: Using New Technologies That Enhance Host Resistance in Apple. Plant Dis. 2003, 87, 756–765. [Google Scholar] [CrossRef]
- Sundin, G.W.; Werner, N.A.; Yoder, K.S.; Aldwinckle, H.S. Field Evaluation of Biological Control of Fire Blight in the Eastern United States. Plant Dis. 2009, 93, 386–394. [Google Scholar] [CrossRef]
- McManus, P.S.; Jones, A.L. Epidemiology and Genetic Analysis of Streptomycin-Resistant Erwinia amylovora from Michigan and Evaluation of Oxytetracycline for Control. Phytopathology 1994, 84, 627–633. [Google Scholar] [CrossRef]
- Loper, J.E.; Henkels, M.D. Evaluation of Streptomycin, Oxytetracycline, and Copper Resistance of Erwinia amylovora Isolated from Pear Orchards in Washington State. Plant Dis. 1991, 75, 287–290. [Google Scholar] [CrossRef]
- Christiano, R.S.C.; Reilly, C.C.; Miller, W.P.; Scherm, H. Oxytetracycline Dynamics on Peach Leaves in Relation to Temperature, Sunlight, and Simulated Rain. Plant Dis. 2010, 94, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Aćimović, S.G.; Mcghee, G.C.; Sundin, G.W.; Wise, J.C. Evaluation of Trunk-Injected Bactericides and Prohexadione-Calcium for Environmentally Friendly Control of Fire Blight (Erwinia amylovora) in Apples. In Proceedings of the 7th Congress on Plant Protection, Zlatibor, Serbia, 24–28 November 2014; Marčić, D.M., Glavendekić, P.N., Eds.; Plant Protection Society of Serbia: Serbia, Belgrade, 2015; pp. 129–134. [Google Scholar]
- Aćimović, S.G.; Zeng, Q.; McGhee, G.C.; Sundin, G.W.; Wise, J.C. Control of Fire Blight (Erwinia amylovora) on Apple Trees with Trunk-Injected Plant Resistance Inducers and Antibiotics and Assessment of Induction of Pathogenesis-Related Protein Genes. Front. Plant Sci. 2015, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Bahder, B.W.; Helmick, E.E.; Chakrabarti, S.; Osorio, S.; Soto, N.; Chouvenc, T.; Harrison, N.A. Disease Progression of a Lethal Decline Caused by the 16SrIV-D Phytoplasma in Florida Palms. Plant Pathol. 2018, 67, 1821–1828. [Google Scholar] [CrossRef]
- Gurr, G.M.; Johnson, A.C.; Ash, G.J.; Wilson, B.A.L.; Ero, M.M.; Pilotti, C.A.; Dewhurst, C.F.; You, M.S. Coconut Lethal Yellowing Diseases: A Phytoplasma Threat to Palms of Global Economic and Social Significance. Front. Plant Sci. 2016, 7, 1521. [Google Scholar] [CrossRef]
- Elliott, M.L. Emerging Palm Diseases in Florida. Horttechnology 2009, 19, 717–718. [Google Scholar] [CrossRef]
- McCoy, R.E.; Norris, R.C.; Vieyra, G.; Delgado, S. Lethal Yellowing Disease of Coconut Palms. FAO Plant Prot. Bull. 1982, 30, 79–80. [Google Scholar]
- Li, M.; Nangong, Z. Precision Trunk Injection Technology for Treatment of Huanglongbing (HLB)-Affected Citrus Trees—A Review. J. Plant Dis. Prot. 2022, 129, 15–34. [Google Scholar] [CrossRef]
- Gottwald, T.R.; da Graça, J.V.; Bassanezi, R.B. Citrus Huanglongbing: The Pathogen and Its Impact. Plant Health Prog. 2007, 8, 31. [Google Scholar] [CrossRef]
- Da Graça, J.V.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G.; Zhao, H. Huanglongbing: An Overview of a Complex Pathosystem Ravaging the World’s Citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar] [CrossRef]
- Bassanezi, R.B.; Primiano, I.V.; Vescove, H.V. Effect of Enhanced Nutritional Programs and Exogenous Auxin Spraying on Huanglongbing Severity, Fruit Drop, Yield and Economic Profitability of Orange Orchards. Crop Prot. 2021, 145, 105609. [Google Scholar] [CrossRef]
- Graca, J.V. Citrus Greening Disease. Annu. Rev. Phytopathol. 1991, 29, 109–136. [Google Scholar] [CrossRef]
- Aubert, B.; Bové, J.M. Effect of Penicillin or Tetracycline Injections of Citrus Trees Affected by Greening Disease Under Field Conditions in Reunion Island. Int. Organ. Citrus Virol. Conf. Proc. 1980, 8, 103–108. [Google Scholar] [CrossRef]
- McVay, J.; Sun, X.; Jones, D.; Urbina, H.; Aldeek, F.; Cook, J.M.; Jeyaprakash, A.; Hodges, G.; Smith, T. Limited Persistence of Residues and Metabolites in Fruit and Juice Following Penicillin Trunk Infusion in Citrus Affected by Huanglongbing. Crop Prot. 2019, 125, 104753. [Google Scholar] [CrossRef]
- Ascunce, M.S.; Shin, K.; Huguet-Tapia, J.C.; Poudel, R.; Garrett, K.A.; van Bruggen, A.H.C.; Goss, E.M. Penicillin Trunk Injection Affects Bacterial Community Structure in Citrus Trees. Microb. Ecol. 2019, 78, 457–469. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, C.; Powell, C.A.; Avery, P.B.; Wang, J.; Huang, Y.; Duan, Y. Field Evaluation of Integrated Management for Mitigating Citrus Huanglongbing in Florida. Front. Plant Sci. 2019, 9, 1890. [Google Scholar] [CrossRef]
- Li, J.; Kolbasov, V.G.; Lee, D.; Pang, Z.; Huang, Y.; Collins, N.; Wang, N. Residue Dynamics of Streptomycin in Citrus Delivered by Foliar Spray and Trunk Injection and Effect on “Candidatus Liberibacter asiaticus” Titer. Phytopathology 2021, 111, 1095–1103. [Google Scholar] [CrossRef]
- Li, J.; Pang, Z.; Duan, S.; Lee, D.; Kolbasov, V.G.; Wang, N. The in Planta Effective Concentration of Oxytetracycline against ‘Candidatus Liberibacter asiaticus’ for Suppression of Citrus Huanglongbing. Phytopathology 2019, 109, 2046–2054. [Google Scholar] [CrossRef]
- Hu, J.; Jiang, J.; Wang, N. Control of Citrus Huanglongbing via Trunk Injection of Plant Defense Activators and Antibiotics. Phytopathology 2018, 108, 186–195. [Google Scholar] [CrossRef]
- Albrecht, U.; Archer, L. Principles and risks of trunk injection for delivery of crop protection materials. Citrus Ind. Mag. 2021, 102, 14–17. [Google Scholar]
- Rumbos, I.C. Control of Plum Chlorotic Leaf Roll Disease by Trunk Pressure Injection of Oxytetracycline-HCI. J. Plant Dis. Prot. 1985, 92, 581–587. [Google Scholar]
- Keil, H.L. Apricot Trunk Injection with Oxytetracycline to Control Bacterial Spot. Proc. Am. Phytopathol. Soc. 1977, 4, 216. [Google Scholar]
- Yang, C.; Powell, C.A.; Duan, Y.; Shatters, R.G.; Lin, Y.; Zhang, M. Mitigating Citrus Huanglongbing via Effective Application of Antimicrobial Compounds and Thermotherapy. Crop Prot. 2016, 84, 150–158. [Google Scholar] [CrossRef]
- Harrison, N.A.; Elliott, M.L. Lethal Yellowing (LY) of Palm. PP-222/PP146 01/2006; EDIS, University of Florida: Gainesville, FL, USA, 2019. [Google Scholar] [CrossRef]
- Cooley, D.R.; Tattar, T.A.; Schieffer, J.T. Treatment of X-Disease of Peaches Using Oxytetracycline Microinjection Capsules. HortScience 1992, 27, 235–237. [Google Scholar] [CrossRef]
- Rosenberger, D.A.; Jones, A.L. Symptom Remission in X-Diseased Peach Trees as Affected by Date, Method, and Rate of Application of Oxytetracycline-HCl. Phytopathology 1977, 77, 277–282. [Google Scholar] [CrossRef]
- Reil, W.O.; Beutel, J.A.; Hemstreet, C.L.; Seyman, W.S. Trunk Injection Corrects Iron and Zinc Deficiency in Pear Trees. Calif. Agric. 1978, 32, 22–23. [Google Scholar]
- McIntyre, J.L.; Schneider, H.; Lacy, G.H.; Dodds, J.A. Pear Decline in Connecticut and Response of Diseased Trees to Oxytetracycline Infusion. Phytopathology 1979, 69, 955–958. [Google Scholar] [CrossRef]
- Lacy, G.H.; McLntyre, J.L.; Walton, G.S.; Dodds, J.A. Rapid Method for and Effects of Infusing Trees with Concentrated Oxytetracycline-HCL Solutions for Pear Decline Control. Can. J. Plant Pathol. 1980, 2, 96–101. [Google Scholar] [CrossRef]
- Du Plessis, H.J. Control of Bacterial Spot on Plums by Sprays of Calcium Oxytetracycline and Copper and Infusion with N-Pyrrolidinomethyl Tetracycline. Phytophylactica 1987, 19, 227–229. [Google Scholar] [CrossRef]
- Puttamuk, T.; Zhang, S.; Duan, Y.; Jantasorn, A.; Thaveechai, N. Effect of Chemical Treatments on “Candidatus Liberibacter asiaticus” Infected Pomelo (Citrus maxima). Crop Prot. 2014, 65, 114–121. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Konkol, J.L.; Pérez-Martínez, J.M.; Fernandez, R. Management of Laurel Wilt of Avocado, Caused by Raffaelea lauricola. Eur. J. Plant Pathol. 2017, 149, 133–143. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi Efficiency, Systemic Properties, and Novel Delivery Methods for Pest Insect Control: What We Know so Far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; von Aderkas, P.; Zaharia, L.I. Effects of Stem-Injected Gibberellins and 6-Benzylaminopurine on Phytohormone Profiles and Cone Yield in Two Lodgepole Pine Genotypes. Trees Struct. Funct. 2018, 32, 765–775. [Google Scholar] [CrossRef]
- Stephano-Hornedo, J.L.; Torres-Gutiérrez, O.; Toledano-Magaña, Y.; Gradilla-Martínez, I.; Pestryakov, A.; Sánchez-González, A.; García-Ramos, J.C.; Bogdanchikova, N. ArgovitTM Silver Nanoparticles to Fight Huanglongbing Disease in Mexican Limes (Citrus aurantifolia Swingle). RSC Adv. 2020, 10, 6146–6155. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Ashworth, V.E.T.M.; Geitner, N.K.; Wiesner, M.R.; Ginnan, N.; Rolshausen, P.; Roper, C.; Jassby, D. Delivery, Fate, and Mobility of Silver Nanoparticles in Citrus Trees. ACS Nano 2020, 14, 2966–2981. [Google Scholar] [CrossRef]
- Beckers, S.J.; Staal, A.H.J.; Rosenauer, C.; Srinivas, M.; Landfester, K.; Wurm, F.R. Targeted Drug Delivery for Sustainable Crop Protection: Transport and Stability of Polymeric Nanocarriers in Plants. Adv. Sci. 2021, 8, 2100067. [Google Scholar] [CrossRef]
- Gardner, C.L.; da Silva, D.R.; Pagliai, F.A.; Pan, L.; Padgett-Pagliai, K.A.; Blaustein, R.A.; Merli, M.L.; Zhang, D.; Pereira, C.; Teplitski, M.; et al. Assessment of Unconventional Antimicrobial Compounds for the Control of ‘Candidatus Liberibacter asiaticus’, the Causative Agent of Citrus Greening Disease. Sci. Rep. 2020, 10, 5395. [Google Scholar] [CrossRef]
- Werrie, P.Y.; Burgeon, C.; le Goff, G.J.; Hance, T.; Fauconnier, M.L. Biopesticide Trunk Injection into Apple Trees: A Proof of Concept for the Systemic Movement of Mint and Cinnamon Essential Oils. Front. Plant Sci. 2021, 12, 650132. [Google Scholar] [CrossRef]
- Scortichini, M.; Loreti, S.; Pucci, N.; Scala, V.; Tatulli, G.; Verweire, D.; Oehl, M.; Widmer, U.; Codina, J.M.; Hertl, P.; et al. Progress towards Sustainable Control of Xylella fastidiosa subsp. pauca in Olive Groves of Salento (Apulia, Italy). Pathogens 2021, 10, 668. [Google Scholar] [CrossRef]
- Hesami, A.; Jafari, N.; Shahriari, M.H.; Zolfi, M. Yield and Physico-Chemical Composition of Date-Palm (Phoenix dactylifera) as Affected by Nitrogen and Zinc Application. Commun. Soil Sci. Plant Anal. 2017, 48, 1943–1954. [Google Scholar] [CrossRef]
- Rosmana, A.; Taufik, M.; Asman, A.; Jayanti, N.J.; Hakkar, A.A. Dynamic of Vascular Streak Dieback Disease Incidence on Susceptible Cacao Treated with Composted Plant Residues and Trichoderma asperellum in Field. Agronomy 2019, 9, 650. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Ager, K.L. Biological Control of Citrus Huanglongbing with Eb92-1, a Benign Strain of Xylella fastidiosa. Plant Dis. 2021, 105, 2914–2918. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.; Czarnocka, K.; Cochard, B.; Oszako, T.; Lefort, F. Biocontrol Endotherapy with Trichoderma spp. and Bacillus amyloliquefaciens against Phytophthora spp.: A Comparative Study with Phosphite Treatment on Quercus robur and Fagus sylvatica. J. Agric. Sci. Technol. A 2015, 5, 428–439. [Google Scholar] [CrossRef]
- Kondo, E.S. A Method for Introducing Water—Soluble Chemicals into Mature Elms; Information Report O-X-171; Canadian Forest Services Publications: Sault Ste. Marie, ON, Canada, 1972. [Google Scholar]
- Stipes, R.J. Glitches and Gaps in the Science and Technology of Tree Injection. J. Arboric. 1988, 14, 165–172. [Google Scholar]
- Aćimović, S.G.; Vanwoerkom, A.H.; Reeb, P.D.; Vandervoort, C.; Garavaglia, T.; Cregg, B.M.; Wise, J.C. Spatial and Temporal Distribution of Trunk-Injected Imidacloprid in Apple Tree Canopies. Pest Manag. Sci. 2014, 70, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.A. Physicochemical Factors Governing the Transport of Xenobiotic Chemicals in Plants: Movement into Roots and Partitioning Between Xylem and Phloem. Acta Hortic. 1989, 239, 43–54. [Google Scholar] [CrossRef]
- Al-Rimawi, F.; Hijaz, F.; Nehela, Y.; Batuman, O.; Killiny, N. Uptake, Translocation, and Stability of Oxytetracycline and Streptomycin in Citrus Plants. Antibiotics 2019, 8, 196. [Google Scholar] [CrossRef] [PubMed]
- Kleier, D.A. Phloem Mobility of Xenobiotics I. Mathematical Model Unifying the Weak Acid and Intermediate Permeability Theories. Plant Physiol. 1988, 86, 803–810. [Google Scholar] [CrossRef]
- Riederer, M. Uptake and Transport of Xenobiotics. In Plant Toxicology; Hock, B., Elstner, E.F., Eds.; Marcel Dekker: New York, NY, USA, 2004; pp. 131–150. [Google Scholar]
- Tyree, M.T.; Peterson, C.A.; Edgington, L.V. A Simple Theory Regarding Ambimobility of Xenobiotics with Special Reference to the Nematicide, Oxamyl. Plant Physiol. 1979, 63, 367–374. [Google Scholar] [CrossRef]
- McCoy, R.E. Comparative Epidemiology of the Lethal Yellowing, Kaincope, and Cadang-Cadang Diseases of Coconut Palm. Plant Dis. Report. 1976, 60, 498–502. [Google Scholar]
- VanWoerkom, A.H.; Aćimović, S.G.; Sundin, G.W.; Cregg, B.M.; Mota-Sanchez, D.; Vandervoort, C.; Wise, J.C. Trunk Injection: An Alternative Technique for Pesticide Delivery in Apples. Crop Prot. 2014, 65, 173–185. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Duffy, B. Use of Antibiotics in Plant Agriculture. OIE Rev. Sci. Tech. 2012, 31, 199–210. [Google Scholar] [CrossRef] [PubMed]
- McManus, P.S.; Stockwell, V.O.; Sundin, G.W.; Jones, A.L. Antibiotic Use in Plant Agriculture. Annu. Rev. Phytopathol. 2002, 40, 443–465. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Reeder, R. Antibiotic Use on Crops in Low and Middle-Income Countries Based on Recommendations Made by Agricultural Advisors. CABI Agric. Biosci. 2020, 1, 1. [Google Scholar] [CrossRef]


{kind=link}
| Crop Species | Pest/Disease | Antibiotic | Efficacy of Trunk Injection | Reference |
|---|---|---|---|---|
| Almond (Prunus dulcis) | Almond leaf scorch (Xylella fastidiosa) | Oxytetracycline, penicillin | Effective | [137] |
| Apple (Malus domestica) | Fire blight (Erwinia amylovora) | Copper chelate, kasugamycin, oxytetracycline | Effective | [144] |
| Apple (Malus domestica) | Fire blight (Erwinia amylovora) | Streptomycin | Effective | [145] |
| Apricot (Prunus armeniaca) | Apricot leaf roll disease | Oxytetracycline | Effective for one season | [163] |
| Apricot (Prunus armeniaca) | Bacterial spot (Xanthomonas pruni) | Oxytetracycline | Effective | [164] |
| Grapefruit (Citrus paradisi) | Huanglongbing (Ca. Liberibacter asiaticus) | Penicillin | Effective | [138] |
| Grapefruit (Citrus paradisi) | Huanglongbing (Ca. Liberibacter asiaticus) | Ampicillin | Effective | [165] |
| Palm (Aracaceae sp.) | Lethal Bronzing (Phytoplasma palmae) | Oxytetracycline | Effective as preventative measure only | [166] |
| Peach (Prunus persica) | Peach X-disease (Mycoplasma sp.) | Oxytetracycline | Effective | [167,168] |
| Pear (Pyrus calleryana) | Pear decline | Oxytetracycline | Effective | [169,170,171] |
| Plum (Prunus domestica) | Bacterial spot (Xanthomonas campestris) | Tetracycline | Effective | [172] |
| Pomelo (Citrus maxima) | Huanglongbing (Ca. Liberibacter asiaticus) | Streptomycin + penicillin + ampicillin | Effective | [173] |
| Sweet orange (Citrus sinensis) | Huanglongbing (Ca. Liberibacter asiaticus) | Oxytetracycline | Effective | [136,161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Archer, L.; Crane, J.H.; Albrecht, U. Trunk Injection as a Tool to Deliver Plant Protection Materials—An Overview of Basic Principles and Practical Considerations. Horticulturae 2022, 8, 552. https://doi.org/10.3390/horticulturae8060552
Archer L, Crane JH, Albrecht U. Trunk Injection as a Tool to Deliver Plant Protection Materials—An Overview of Basic Principles and Practical Considerations. Horticulturae. 2022; 8(6):552. https://doi.org/10.3390/horticulturae8060552
Chicago/Turabian StyleArcher, Leigh, Jonathan H. Crane, and Ute Albrecht. 2022. "Trunk Injection as a Tool to Deliver Plant Protection Materials—An Overview of Basic Principles and Practical Considerations" Horticulturae 8, no. 6: 552. https://doi.org/10.3390/horticulturae8060552