
Hazelnut-Associated Bacteria and Their Implications in Crop Management




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
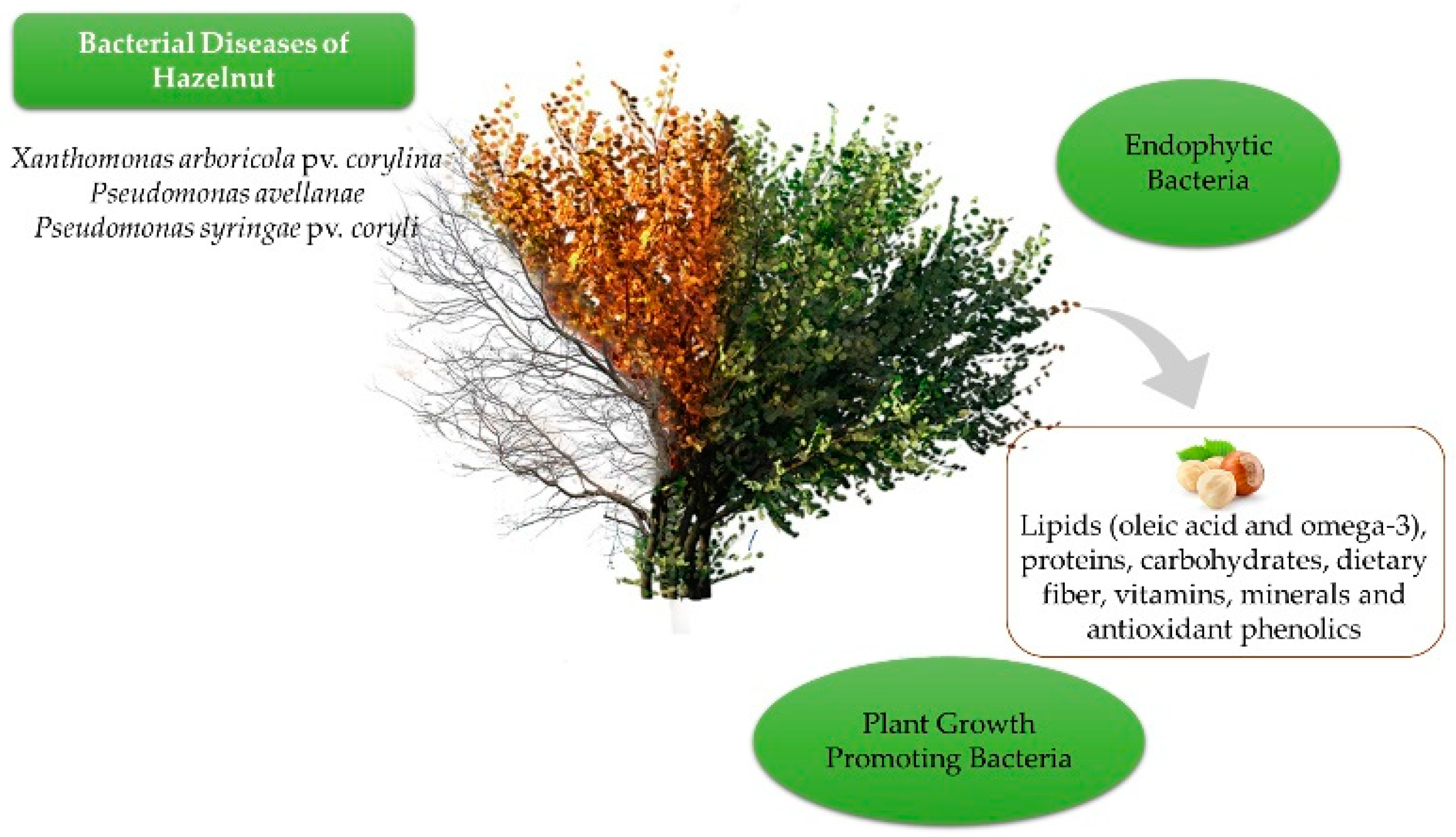
2. Bacterial Diseases of Hazelnut
2.1. Bacterial Blight
2.2. Bacterial Canker
2.3. Other Bacterial Pathogens
3. Management of Bacterial Diseases of Hazelnut
4. Other Bacteria Associated with Hazelnut
4.1. Plant Growth Promoters
4.2. Endophytes
4.3. Epiphytes and Root Associates
5. Antibacterial Properties of Hazelnut
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. World Food and Agriculture-Statistical Yearbook; FAO: Rome, Italy, 2020; p. 366. [Google Scholar] [CrossRef]
- Silvestri, C.; Bacchetta, L.; Bellincontro, A.; Cristofori, V. Advances in cultivar choice, hazelnut orchard management, and nut storage to enhance product quality and safety: An overview. J. Sci. Food Agric. 2021, 101, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Alasalvar, C.; Shahidi, F.; Liyanapathirana, C.M.; Ohshima, T. Turkish tombul hazelnut (Corylus avellana L.). 1. Compositional characteristics. J. Agric. Food Chem. 2003, 51, 3790–3796. [Google Scholar] [CrossRef] [PubMed]
- Köksal, A.İ.; Artik, N.; Şimşek, A.; Güneş, N. Nutrient composition of hazelnut (Corylus avellana L.) varieties cultivated in Turkey. Food Chem. 2006, 99, 509–515. [Google Scholar] [CrossRef]
- Jakopic, J.; Petkovsek, M.M.; Likozar, A.; Solar, A.; Stampar, F.; Veberic, R. HPLC–MS identification of phenols in hazelnut (Corylus avellana L.) kernels. Food Chem. 2011, 124, 1100–1106. [Google Scholar] [CrossRef]
- Shahidi, F.; Alasalvar, C.; Liyana-Pathirana, C.M. Antioxidant phytochemicals in hazelnut kernel (Corylus avellana L.) and hazelnut byproducts. J. Agric. Food Chem. 2007, 55, 1212–1220. [Google Scholar] [CrossRef]
- Pelvan, E.; Alasalvar, C.; Uzman, S. Effects of roasting on the antioxidant status and phenolic profiles of commercial Turkish hazelnut varieties (Corylus avellana L.). J. Agric. Food Chem. 2012, 60, 1218–1223. [Google Scholar] [CrossRef]
- Cristofori, V.; Ferramondo, S.; Bertazza, G.; Bignami, C. Nut and kernel traits and chemical composition of hazelnut (Corylus avellana L.) cultivars. J. Sci. Food Agric. 2008, 88, 1091–1098. [Google Scholar] [CrossRef]
- Bacchetta, L.; Aramini, M.; Zini, A.; Di Giammatteo, V.; Spera, D.; Drogoudi, P.; Rovira, M.; Silva, A.P.; Solar, A.; Botta, R. Fatty acids and alpha-tocopherol composition in hazelnut (Corylus avellana L.): A chemometric approach to emphasize the quality of European germplasm. Euphytica 2013, 191, 57–73. [Google Scholar] [CrossRef]
- Lee, J.H.; Lavie, C.J.; O’Keefe, J.H.; Milani, R. Nuts and seeds in cardiovascular health. In Nuts and Seeds in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 75–82. [Google Scholar]
- Contini, M.; Frangipane, M.T.; Massantini, R. Antioxidants in hazelnuts (Corylus avellana L.). In Nuts and Seeds in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 611–625. [Google Scholar]
- Shao, F.; Wilson, I.W.; Qiu, D. The research progress of taxol in Taxus. Curr Pharm Biotechnol 2021, 22, 360–366. [Google Scholar] [CrossRef]
- Gallego, A.; Malik, S.; Yousefzadi, M.; Makhzoum, A.; Tremouillaux-Guiller, J.; Bonfill, M. Taxol from Corylus avellana: Paving the way for a new source of this anti-cancer drug. Plant Cell Tissue Organ Cult. 2017, 129, 1–16. [Google Scholar] [CrossRef]
- Bottone, A.; Cerulli, A.; D’Urso, G.; Masullo, M.; Montoro, P.; Napolitano, A.; Piacente, S. Plant specialized metabolites in hazelnut (Corylus avellana) kernel and byproducts: An update on chemistry, biological activity, and analytical aspects. Planta Med. 2019, 85, 840–855. [Google Scholar] [CrossRef] [Green Version]
- Barss, H.P. A new filbert disease in Oregon. Oregon Agric. Coll. Exp. Sta. Biennial Crop Pest Hort. Rep. 1913, 14, 213–223. [Google Scholar]
- Miller, P.W.; Bollen, W.B.; Simmons, J.E.; Gross, H.N.; Barss, H.P. The pathogen of filbert bacteriosis compared with Phytomonas juglandis, the cause of walnut blight. Phytopathology 1940, 30, 713–733. [Google Scholar]
- Sutic, D. Bacterial spots on leaves of filbert. Zast Biija 1956, 37, 47–53. [Google Scholar]
- Noviello, C. Osservazioni sulle malattie parassitarie del nocciolo, con particolare riferimento alla Campania. Ann. Fac. Sci. Agrar. Univ. Studi Napoli Portici 1968, 3, 11–39. [Google Scholar]
- Alay, K.; Altinyay, N.; Hancioglu, O.; Dundar, F.; Unal, A. Studies on desiccation of hazel nut branches in the Black Sea region. Bitki Koruma Bul. 1973, 13, 202–213. [Google Scholar]
- Luisetti, J.; Jailloux, F.; Germain, E.; Prunier, J.P.; Gardan, L. Caracterisation de Xanthomonas corylina (Miller et al.) Starr et Burkholder responsable de la bacteriose du noisetier recemment observee en France. Compte Rendu Acad. Agric. Fr. 1975, 61, 845–849. [Google Scholar]
- Koval, G.K. Diseases of hazel. Zashchita Rastenii 1978, 8, 44–45. [Google Scholar]
- Locke, T.; Barnes, D. Xanthomonas corylina on cob-nuts and filberts. Plant Pathol. 1979, 28, 53. [Google Scholar] [CrossRef]
- Wimalajeewa, D.L.S.; Washington, W.S. Bacterial blight of hazelnut. Australas. Plant Pathol. 1980, 9, 113–114. [Google Scholar] [CrossRef]
- Guerrero, C.J.; Lobos, A.W. Xanthomonas campestris pv. corylina, causal agent of bacterial blight of hazel in Region IX, Chile. Agric. Técnica 1987, 47, 422–426. [Google Scholar]
- Kazempour, M.N.; Ali, B.; Elahinia, S.A. First report of bacterial blight of hazelnut caused by Xanthomonas arboricola pv. corylina in Iran. J. Plant Pathol. 2006, 88, 341. [Google Scholar]
- Pulawska, J.; Kaluzna, M.; Kolodziejska, A.; Sobiczewski, P. Identification and characterization of Xanthomonas arboricola pv. corylina causing bacterial blight of hazelnut: A new disease in Poland. J. Plant Pathol. 2010, 92, 803–806. [Google Scholar]
- Kałużna, M.; Fischer-Le Saux, M.; Pothier, J.F.; Jacques, M.A.; Obradović, A.; Tavares, F.; Stefani, E. Xanthomonas arboricola pv. juglandis and pv. corylina: Brothers or distant relatives? Genetic clues, epidemiology, and insights for disease management. Mol. Plant Pathol. 2021, 22, 1481–1499. [Google Scholar] [PubMed]
- Webber, J.B.; Putnam, M.; Serdani, M.; Pscheidt, J.W.; Wiman, N.G.; Stockwell, V.O. Characterization of isolates of Xanthomonas arboricola pv. corylina, the causal agent of bacterial blight, from Oregon hazelnut orchards. J. Plant Pathol. 2020, 102, 799–812. [Google Scholar]
- Webber, J.B.; Wada, S.; Stockwell, V.O.; Wiman, N.G. Susceptibility of some Corylus avellana L. cultivars to Xanthomonas arboricola pv. corylina. Front. Plant Sci. 2021, 12, 800339. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH); Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Jacques, M.A.; Miret, J.A.J.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; et al. Commodity risk assessment of Corylus avellana and Corylus colurna plants from Serbia. EFSA J. 2021, 19, e06571. [Google Scholar]
- Lamichhane, J.R.; Varvaro, L. Xanthomonas arboricola disease of hazelnut: Current status and future perspectives for its management. Plant Pathol. 2014, 63, 243–254. [Google Scholar] [CrossRef]
- Scortichini, M.; Rossi, M.P.; Marchesi, U. Genetic, phenotypic and pathogenic diversity of Xanthomonas arboricola pv. corylina strains question the representative nature of the type strain. Plant Pathol. 2002, 51, 374–381. [Google Scholar]
- Fischer-Le Saux, M.; Bonneau, S.; Essakhi, S.; Manceau, C.; Jacques, M.A. Aggressive emerging pathovars of Xanthomonas arboricola represent widespread epidemic clones distinct from poorly pathogenic strains, as revealed by multilocus sequence typing. Appl. Environ. Microbiol. 2015, 81, 4651–4668. [Google Scholar] [CrossRef] [Green Version]
- Caballero, J.I.; Zerillo, M.M.; Snelling, J.; Boucher, C.; Tisserat, N. Genome sequence of Xanthomonas arboricola pv. corylina, isolated from Turkish filbert in Colorado. Genome Announ. 2013, 1, e00246-13. [Google Scholar]
- Pothier, J.F.; Kałużna, M.; Prokić, A.; Obradović, A.; Rezzonico, F. Complete genome and plasmid sequence data of three strains of Xanthomonas arboricola pv. corylina, the bacterium responsible for bacterial blight of hazelnut. Phytopathology 2022, 112, 956–960. [Google Scholar]
- Psallidas, P.G.; Panagopoulos, C.G. A bacterial canker of Corylus avellana in Greece. J. Phytopathol. 1979, 94, 103–111. [Google Scholar] [CrossRef]
- Psallidas, P.G. Pseudomonas syringae pv. avellanae pathovar nov., the bacterium causing canker disease on Corylus avellana. Plant Pathol. 1993, 42, 358–363. [Google Scholar]
- Scortichini, M.; Tropiano, F.G. Severe outbreak of Pseudomonas syringae pv. avellanae on hazelnut in Italy. J. Phytopathol. 1994, 140, 65–70. [Google Scholar]
- Brzezinski, M.G. Le chancre des arbres, ses causes et ses symptoms. Bull. Intern. Acad. Sci. Cracovie 1903, 7, 139–140. [Google Scholar]
- Thornberry, H.H.; Anderson, H.W. Some bacterial diseases of plants in Illinois. Phytopathology 1937, 27, 946–949. [Google Scholar]
- Janse, J.D.; Rossi, P.; Angelucci, L.; Scortichini, M.; Derks, J.H.J.; Akkermans, A.D.L.; De Vrijer, R.; Psallidas, P.G. Reclassification of Pseudomonas syringae pv. avellanae as Pseudomonas avellanae (spec. nov, the bacterium causing canker of hazelnut (Corylus avellana L.). Syst. Appl. Microbiol. 1996, 19, 589–595. [Google Scholar]
- Marcelletti, S.; Scortichini, M. Definition of plant-pathogenic Pseudomonas genomospecies of the P. syringae complex through multiple comparative approaches. Phytopathology 2014, 104, 1274–1282. [Google Scholar] [CrossRef]
- Scortichini, M.; Marchesi, U.; Dettori, M.T.; Angelucci, L.; Rossi, M.P.; Morone, C. Genetic and pathogenic diversity of Pseudomonas avellanae strains isolated from Corylus avellana trees in north-west of Italy, and comparison with strains from other regions. Eur. J. Plant Pathol. 2000, 106, 147–154. [Google Scholar] [CrossRef]
- Scortichini, M.; Marchesi, U.; Rossi, M.P.; Di Prospero, P. Bacteria associated with hazelnut (Corylus avellana L.) decline are of two groups: Pseudomonas avellanae and strains resembling P. syringae pv. syringae. Appl. Environ. Microbiol. 2002, 68, 476–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaluzna, M.; Ferrante, P.; Sobiczewski, P.; Scortichini, M. Characterization and genetic diversity of Pseudomonas syringae from stone fruits and hazelnut using repetitive-PCR and MLST. J. Plant Pathol. 2010, 92, 781–787. [Google Scholar]
- Scortichini, M.; Rossi, M.P.; Loreti, S.; Bosco, A.; Fiori, M.; Jackson, R.W.; Stead, D.E.; Aspin, A.; Marchesi, U.; Zini, M.; et al. Pseudomonas syringae pv. coryli, the causal agent of bacterial twig dieback of Corylus avellana. Phytopathology 2005, 95, 1316–1324. [Google Scholar] [PubMed] [Green Version]
- Scortichini, M.; Marcelletti, S.; Ferrante, P.; Firrao, G. A genomic redefinition of Pseudomonas avellanae species. PLoS ONE 2013, 8, e75794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, O.; Monteil, C.L.; Bartoli, C.; Chandeysson, C.; Guilbaud, C.; Sands, D.C.; Morris, C.E. A user’s guide to a data base of the diversity of Pseudomonas syringae and its application to classifying strains in this phylogenetic complex. PLoS ONE 2014, 9, e105547. [Google Scholar] [CrossRef]
- Scortichini, M.; Ferrante, P.; Cozzolino, L.; Zoina, A. Emended description of Pseudomonas syringae pv. avellanae, causal agent of European hazelnut (Corylus avellana L.) bacterial canker and decline. Eur. J. Plant Pathol. 2016, 144, 213–215. [Google Scholar]
- Marcelletti, S.; Scortichini, M. Comparative genomic analyses of multiple Pseudomonas strains infecting Corylus avellana trees reveal the occurrence of two genetic clusters with both common and distinctive virulence and fitness traits. PLoS ONE 2015, 10, e0131112. [Google Scholar] [CrossRef]
- Turco, S.; Zuppante, L.; Drais, M.I.; Mazzaglia, A. Dressing like a pathogen: Comparative analysis of different Pseudomonas genomospecies wearing different features to infect Corylus avellana. J. Phytopathol. 2022, 170, 504–516. [Google Scholar] [CrossRef]
- Maleki-Zadeh, H.R.; Falahi Charkhabi, N.; Khodaygan, P.; Rahimian, H. Bacterial leaf spot and die-back of hazelnut caused by a new pathovar of Pseudomonas amygdali. Eur. J. Plant Pathol. 2022, 163, 293–303. [Google Scholar] [CrossRef]
- Bucini, D.; Balestra, G.M.; Pucci, C.; Paparatti, B.; Speranza, S.; Proietti Zolla, C.; Varvaro, L. Bio-ethology of Anisandrus dispar F. and its possible involvement in dieback (Moria) diseases of hazelnut (Corylus avellana L.) plants in central Italy. Acta Hort. 2004, 686, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, J.C.; Pérez, S.F.; Ferrada, E.Q.; Cona, L.Q.; Bensch, E.T. Phytopathogens of hazelnut (Corylus avellana L.) in southern Chile. Acta Hort. 2014, 1052, 269–274. [Google Scholar] [CrossRef]
- Krol, E.; Machowicz-Stefaniak, Z.; Zalewska, E. Bacteria damaging the fruit of hazel (Corylus avellana L.) cultivated in South-East Poland. Acta Sci. Pol. Hortorum Cultus 2004, 3, 75–84. [Google Scholar]
- Marcone, C.; Ragozzino, A.; Seemüller, E. Association of phytoplasmas with the decline of European hazel in southern Italy. Plant Pathol. 1996, 45, 857–863. [Google Scholar] [CrossRef]
- Postman, J.D.; Johnson, K.B.; Jomantiene, R.; Maas, J.L.; Davis, R.E. The Oregon hazelnut stunt syndrome and phytoplasma associations. Acta Hort. 2001, 556, 407–409. [Google Scholar] [CrossRef]
- Cieślińska, M.; Kowalik, B. Detection and molecular characterization of ‘Candidatus Phytoplasma asteris’ in European hazel (Corylus avellana) in Poland. J. Phytopathol. 2011, 159, 585–588. [Google Scholar] [CrossRef]
- Hodgetts, J.; Flint, L.J.; Davey, C.; Forde, S.; Jackson, L.; Harju, V.; Skelton, A.; Fox, A. Identification of’ Candidatus Phytoplasma fragariae’(16Sr XII-E) infecting Corylus avellana (hazel) in the United Kingdom. New Dis. Rep. 2015, 32, 3. [Google Scholar] [CrossRef] [Green Version]
- Mehle, N.; Jakoš, N.; Mešl, M.; Miklavc, J.; Matko, B.; Rot, M.; Ferlež Rus, A.; Brus, R.; Dermastia, M. Phytoplasmas associated with declining of hazelnut (Corylus avellana) in Slovenia. Eur. J. Plant Pathol. 2019, 155, 1117–1132. [Google Scholar] [CrossRef] [Green Version]
- Gentili, A.; Donati, L.; Bertin, S.; Manglli, A.; Ferretti, L. First report of ‘Candidatus Phytoplasma fragariae’ infecting hazelnut in Italy. Plant Dis. 2022, 106, 2254. [Google Scholar] [CrossRef]
- Lessio, F.; Picciau, L.; Gonella, E.; Tota, F.; Mandrioli, M.; Alma, A. The mosaic leafhopper Orientus ishidae: Host plants, spatial distribution, infectivity, and transmission of 16SrV phytoplasmas to vines. Bull. Insectol. 2016, 69, 277–289. [Google Scholar]
- Mehle, N.; Dermastia, M. Towards the evaluation of potential insect vectors of phytoplasmas infecting hazelnut plants in Slovenia. Phytopath. Mollicutes 2019, 9, 49–50. [Google Scholar] [CrossRef]
- Vuono, G.; Balestra, G.M.; Varvaro, L. Control of dieback (“Moria”) of Corylus avellana in central Italy using copper compounds. J. Plant Pathol. 2006, 88, 215–218. [Google Scholar]
- Scortichini, M.; Liguori, R. Integrated management of bacterial decline of hazelnut, by using Bion as an activator of systemic acquired resistance (SAR). In Pseudomonas Syringae and Related Pathogens; Springer: Dordrecht, The Netherlands, 2003; pp. 483–487. [Google Scholar]
- Schiavi, D.; Ronchetti, R.; Di Lorenzo, V.; Salustri, M.; Petrucci, C.; Vivani, R.; Giovagnoli, S.; Camaioni, E.; Balestra, G.M. Circular hazelnut protection by lignocellulosic waste valorization for nanopesticides development. Appl. Sci. 2022, 12, 2604. [Google Scholar] [CrossRef]
- Pisetta, M.; Albertin, I.; Petriccione, M.; Scortichini, M. Effects of hot water treatment to control Xanthomonas arboricola pv. corylina on hazelnut (Corylus avellana L.) propagative material. Sci. Hort. 2016, 211, 187–193. [Google Scholar]
- Prokić, A.; Ivanović, M.; Gasic, K.; Kuzmanovic, N.; Zlatković, N.; Obradovic, A. Studying Xanthomonas arboricola pv. corylina strains from Serbia for streptomycin and kasugamycin resistance and copper sulfate sensitivity in vitro. In Proceedings of the 12th International Congress of Plant Pathology: Plant Health in a Global Economy, Boston, MA, USA, 29 July–3 August 2018; Available online: https://apsnet.confex.com/apsnet/ICPP2018/meetingapp.cgi/Paper/11264 (accessed on 15 October 2022).
- European Food Safety Authority (EFSA); Arena, M.; Auteri, D.; Barmaz, S.; Bellisai, G.; Brancato, A.; Brocca, D.; Bura, L.; Byers, H.; Chiusolo, A.; et al. Peer review of the pesticide risk assessment of the active substance copper compounds copper(I), copper(II) variants namely copper hydroxide, copper oxychloride, tribasic copper sulfate, copper(I) oxide, Bordeaux mixture. EFSA J. 2018, 16, e05152. [Google Scholar] [PubMed] [Green Version]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria: Role in soil improvement, abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef]
- Rostamikia, Y.; Kouchaksaraei, M.T.; Asgharzadeh, A.; Rahmani, A. The effect of plant growth-promoting rhizobacteria on growth and physiological characteristics of Corylus avellana seedlings. Ecopersia 2016, 4, 1471–1479. [Google Scholar] [CrossRef]
- Rostamikia, Y.; Kouchaksaraei, M.T.; Asgharzadeh, A.; Rahmani, A. Effect of growth promoting rhizobacteria on growth and nutrient elements of common hazelnut (Corylus avellana L.) seedlings in Ardabil Fandoqlou nursery. Iran J. For.Poplar Res. 2017, 25, 116–126. [Google Scholar]
- Luziatelli, F.; Ficca, A.G.; Bonini, P.; Muleo, R.; Gatti, L.; Meneghini, M.; Tronati, M.; Melini, F.; Ruzzi, M. A genetic and metabolomic perspective on the production of indole-3-acetic acid by Pantoea agglomerans and use of their metabolites as biostimulants in plant nurseries. Front. Microbiol. 2020, 11, 1475. [Google Scholar] [CrossRef]
- Nicoletti, R.; Di Vaio, C.; Cirillo, C. Endophytic fungi of olive tree. Microorganisms 2020, 8, 1321. [Google Scholar] [CrossRef]
- Nicoletti, R.; Beccaro, G.L.; Sekara, A.; Cirillo, C.; Di Vaio, C. Endophytic fungi and ecological fitness of chestnuts. Plants 2021, 10, 542. [Google Scholar] [CrossRef]
- Vandana, U.K.; Rajkumari, J.; Singha, L.P.; Satish, L.; Alavilli, H.; Sudheer, P.D.; Chauhan, S.; Ratnala, R.; Satturu, V.; Mazumder, P.B.; et al. The endophytic microbiome as a hotspot of synergistic interactions, with prospects of plant growth promotion. Biology 2021, 10, 101. [Google Scholar] [CrossRef]
- Pathak, P.; Rai, V.K.; Can, H.; Singh, S.K.; Kumar, D.; Bhardwaj, N.; Roychowdhury, R.; Basilio de Azevedo, L.C.; Kaushalendra, K.; Verma, H.; et al. Plant-endophyte interaction during biotic stress management. Plants 2022, 11, 2203. [Google Scholar] [CrossRef]
- Scortichini, M.; Loreti, S. Occurrence of an endophytic, potentially pathogenic strain of Pseudomonas syringae in symptomless wild trees of Corylus avellana L. J. Plant Pathol. 2007, 89, 431–434. [Google Scholar]
- Loreti, S.; Gallelli, A.; De Simone, D.; Bosco, A. Detection of Pseudomonas avellanae and the bacterial microflora of hazelnut affected by ‘moria’ in central Italy. J. Plant Pathol. 2009, 91, 365–373. [Google Scholar]
- Reed, B.M.; Mentzer, J.; Tanprasert, P.; Yu, X. Internal bacterial contamination of micropropagated hazelnut: Identification and antibiotic treatment. Plant Cell Tissue Organ Cult. 1998, 52, 67–70. [Google Scholar] [CrossRef]
- Liu, H.; Brettell, L.E.; Qiu, Z.; Singh, B.K. Microbiome-mediated stress resistance in plants. Trends Plant Sci. 2020, 25, 733–743. [Google Scholar] [CrossRef]
- Foo, J.L.; Ling, H.; Lee, Y.S.; Chang, M.W. Microbiome engineering: Current applications and its future. Biotechnol. J. 2017, 12, 1600099. [Google Scholar] [CrossRef]
- Hand, C.R.; Wada, N.; Stockwell, V.; Reed, B.M. Node position influences viability and contamination in hazelnut shoot explants. In Vitro Cell. Dev. Biol. Plant 2016, 52, 580–589. [Google Scholar] [CrossRef]
- Zalewska, E. Effect of phyllosphere microorganisms on the growth of Monilia coryli. Phytopathol. Pol. 1999, 18, 57–67. [Google Scholar]
- Krol, E.; Machowicz-Stefaniak, Z.; Zalewska, E. The possibilities of limiting some pathogens of hazel by antagonistic bacteria. Acta Sci. Pol. Hortorum Cultus 2003, 2, 51–57. [Google Scholar]
- Manirajan, B.A.; Suarez, C.; Ratering, S.; Rusch, V.; Geissler-Plaum, R.; Cardinale, M.; Schnell, S. Spirosoma pollinicola sp. nov., isolated from pollen of common hazel (Corylus avellana L.). Int. J. Syst. Evol. Microbiol. 2018, 68, 3248–3254. [Google Scholar] [CrossRef] [PubMed]
- Deveau, A.; Antony-Babu, S.; Le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Temporal changes of bacterial communities in the Tuber melanosporum ectomycorrhizosphere during ascocarp development. Mycorrhiza 2016, 26, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, X.; Yang, M.; Yan, L.; Kang, Z.; Xiao, Y.; Tang, P.; Ye, L.; Zhang, B.; Zou, J.; et al. Tuber borchii shapes the ectomycorrhizosphere microbial communities of Corylus avellana. Mycobiology 2019, 47, 180–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Zou, J.; Liu, C.; Xiao, Y.; Zhang, X.; Yan, L.; Ye, L.; Tang, P.; Li, X. Chinese white truffles shape the ectomycorrhizal microbial communities of Corylus avellana. Ann. Microbiol. 2019, 69, 553–565. [Google Scholar] [CrossRef]
- Pershern, A.S.; Breene, W.M.; Lulai, E.C. Analysis of factors influencing lipid oxidation in hazelnuts (Corylus spp.). J. Food Proc. Preserv. 1995, 19, 9–26. [Google Scholar] [CrossRef]
- Leichtfried, D.; Krist, S.; Puchinger, L.; Messner, K.; Buchbauer, G. Investigations of the natural microflora of poppy seeds (Papaver somniferum) and hazelnut kernels (Corylus avellana) including microbiological decomposition. Eur. Food Res. Technol. 2004, 219, 282–285. [Google Scholar] [CrossRef]
- Kit, W.S.; Priya, M.; Chin, J.H.; Mariam, A.; Akowuah, G.A. Antimicrobial and antiradical activities of Corylus cornuta (marsh., betulacea) kernel extracts. Orient. Pharm. Exp. Med. 2016, 16, 45–51. [Google Scholar] [CrossRef]
- Oliveira, I.; Sousa, A.; Morais, J.S.; Ferreira, I.C.; Bento, A.; Estevinho, L.; Pereira, J.A. Chemical composition, and antioxidant and antimicrobial activities of three hazelnut (Corylus avellana L.) cultivars. Food Chem. Toxicol. 2008, 46, 1801–1807. [Google Scholar] [CrossRef]
- Oliveira, I.; Sousa, A.; Valentão, P.; Andrade, P.B.; Ferreira, I.C.; Ferreres, F.; Bento, A.; Seabra, R.; Estevinho, L.; Pereira, J.A. Hazel (Corylus avellana L.) leaves as source of antimicrobial and antioxidative compounds. Food Chem. 2007, 105, 1018–1025. [Google Scholar] [CrossRef]
- Nikolaieva, N.; Kačániová, M.; González, J.C.; Grygorieva, O.; Nôžková, J. Determination of microbiological contamination, antibacterial and antioxidant activities of natural plant hazelnut (Corylus avellana L.) pollen. J. Environ. Sci. Health B 2019, 54, 525–532. [Google Scholar] [CrossRef]
- Amarowicz, R.; Dykes, G.A.; Pegg, R.B. Antibacterial activity of tannin constituents from Phaseolus vulgaris, Fagoypyrum esculentum, Corylus avellana and Juglans nigra. Fitoterapia 2008, 79, 217–219. [Google Scholar] [CrossRef]
- Cerulli, A.; Lauro, G.; Masullo, M.; Cantone, V.; Olas, B.; Kontek, B.; Nazzaro, F.; Bifulco, G.; Piacente, S. Cyclic diarylheptanoids from Corylus avellana green leafy covers: Determination of their absolute configurations and evaluation of their antioxidant and antimicrobial activities. J. Nat. Prod. 2017, 80, 1703–1713. [Google Scholar] [CrossRef]
- Scortichini, M. Sustainable management of diseases in horticulture: Conventional and new options. Horticulturae 2022, 8, 517. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicoletti, R.; Petriccione, M.; Curci, M.; Scortichini, M. Hazelnut-Associated Bacteria and Their Implications in Crop Management. Horticulturae 2022, 8, 1195. https://doi.org/10.3390/horticulturae8121195
Nicoletti R, Petriccione M, Curci M, Scortichini M. Hazelnut-Associated Bacteria and Their Implications in Crop Management. Horticulturae. 2022; 8(12):1195. https://doi.org/10.3390/horticulturae8121195
Chicago/Turabian StyleNicoletti, Rosario, Milena Petriccione, Martina Curci, and Marco Scortichini. 2022. "Hazelnut-Associated Bacteria and Their Implications in Crop Management" Horticulturae 8, no. 12: 1195. https://doi.org/10.3390/horticulturae8121195