
Fruit Colour and Novel Mechanisms of Genetic Regulation of Pigment Production in Tomato Fruits

Plant Lab, Institute of Life Sciences, Scuola Superiore Sant’Anna, 56127 Pisa, Italy
*
Author to whom correspondence should be addressed.
Horticulturae 2021, 7(8), 259; https://doi.org/10.3390/horticulturae7080259
Submission received: 29 July 2021
/
Revised: 17 August 2021
/
Accepted: 19 August 2021
/
Published: 21 August 2021
(This article belongs to the Special Issue Genetic Modification of Flower and Fruit Color Variations in Horticultural Crops)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Fruit colour represents a genetic trait with ecological and nutritional value. Plants mainly use colour to attract animals and favour seed dispersion. Thus, in many species, fruit colour coevolved with frugivories and their preferences. Environmental factors, however, represented other adaptive forces and further diversification was driven by domestication. All these factors cooperated in the evolution of tomato fruit, one of the most important in human nutrition. Tomato phylogenetic history showed two main steps in colour evolution: the change from green-chlorophyll to red-carotenoid pericarp, and the loss of the anthocyanic pigmentation. These events likely occurred with the onset of domestication. Then spontaneous mutations repeatedly occurred in carotenoid and phenylpropanoid pathways, leading to colour variants which often were propagated. Introgression breeding further enriched the panel of pigmentation patterns. In recent decades, the genetic determinants underneath tomato colours were identified. Novel evidence indicates that key regulatory and biosynthetic genes undergo mechanisms of gene expression regulation that are much more complex than what was imagined before: post-transcriptional mechanisms, with RNA splicing among the most common, indeed play crucial roles to fine-tune the expression of this trait in fruits and offer new substrate for the rise of genetic variables, thus providing further evolutionary flexibility to the character.
1. Fruit Colour and Its Evolution in Tomato Plants
1.1. Function and Evolution of Fruit Colour
Colour is one of the main qualitative attributes of fleshy fruits. Its importance is strictly dependent on its primary ecological function, which is attracting seed dispersers. In a typical example of mutualistic interaction, many plants attract animals by offering their coloured fruits as a nutritious reward in return for their seed dispersal activity. A wide range of different coloration, spanning from white to yellow, red, green, purple, and black, can stain fruit peel and flesh, according to different biotic and abiotic factors and to the developmental stage of fruits. Many fruits, especially when characterized by long ripening periods, change colour during their development, often acquiring more striking and/or contrastive hues at the end of the process.
Both the origin and the adaptive meaning of fruit colours, as well as the high variability of this trait, have been objects of study for long time. Different variables have been considered, from the role of the mutualists, to phylogenetic or environmental constraints. One of the most common interpretations is the disperser syndrome hypothesis, which is in favour of a main role of biotic factors (bird and mammal frugivories, in particular) in selecting fruit colours according to specific visual perception abilities [1]. It is quite common that seed-dispersing birds prefer red and black fruits, whereas fruits with more cryptic colours, yellow, green, or brown, are preferred by mammals frugivories [2]. Under this perspective, colour may represent a specific signal indicating precise messages, such as the degree of fruit maturity, to avoid premature fruit removal from the plant, and can facilitate recognition of specific food sources, such as certain carotenoids, as nutritional rewards [1].
Abiotic factors, mainly temperature, altitude, ripening season length, and UV-B radiation, can in turn account for part of the variation existing in nature as far as fruit colours are concerned [3]. The presence of specific classes of pigments can represent an active defence towards adverse environmental conditions or pathogens. Anthocyanins, for example, can offer photoprotection against intense solar radiation and can also absorb more solar radiation when temperature is low to increase fruit metabolic and developmental rates [1]. Thus, environmental factors can also represent drivers of diversification of fruit colour.
The most recent studies tend to adopt integrative approaches to examine the different actors in fruit colour diversification and consider dispersers and environment equally important and interacting variables [2,4]. Contrastive colours, primarily linked with anthocyanin pigments, appear in fact to be mainly associated with bird frugivories in warm areas, whereas in cold areas or where UV-B radiation is high, contrastive hues seem to be prevalent in any case, even in the presence of mammals [4]. Interestingly, the hypothesis that fruit colour may also be result of phylogenetic constraints [5,6] has progressively lost importance [7]. Colour as other fruit traits, including size and scent, can be dynamically shaped by frugivories and in many taxa, and for some groups of animals (e.g., olfactory-dependent vertebrates), are the object of coevolution more than a phylogenetic signal [8,9,10]. This could not be truer in fruits which contribute to human nutrition. The domestication process of crop plants, which have been progressively selected for adaptation to cultivation for human needs, represented an anthropogenic-driven pressure of adaptive evolution, especially in terms of fruit size, shape, colour, flavour, and texture [11,12]. However, human selection, while detecting and improving desirable phenotypic traits, largely contributed to reduce the genetic variability of cultivated plants, as occurred with tomato with the near fixation of about 25% of its genome [13]. This could have slowed down the further diversification of specific traits. In these cases, the use of wild germplasm as a source of new alleles, also known as “introgression breeding”, continues to represent an important tool to increase genetic variability in cultivated crops.
1.2. Carotenoid and Flavonoid Pigments as Determinants of Fruit Colour in Solanaceae
The Solanaceae represents one of the most important families of flowering plants, with more than 2.900 different species [14]. It includes fruit crops, such as tomato (Solanum lycopersicum), chili/pepper (Capsicum spp.), and eggplant (Solanum melongena), tuber vegetables, such as potato (Solanum tuberosum), medicinal (Datura stramonium, Mandragora officinarum, Atropa belladonna), and ornamental plants (Petunia spp., Physalis spp.). Some of these species represent models for the study of important physiological processes, particularly tomato and potato, tobacco (Nicotiana tabacum), and petunia (Petunia x hybrida). A great diversity of habitats and ecosystems characterize these plants, which are spread throughout the continents, with the main centre of taxonomic diversity in South America [14]. The fruits of the Solanaceae mostly belong to berry and capsule types and can strongly differ in size, shape, and colour [15]. Some of these traits were commonly selected in different species, particularly with domestication, whereas others appear more species-specific [11].
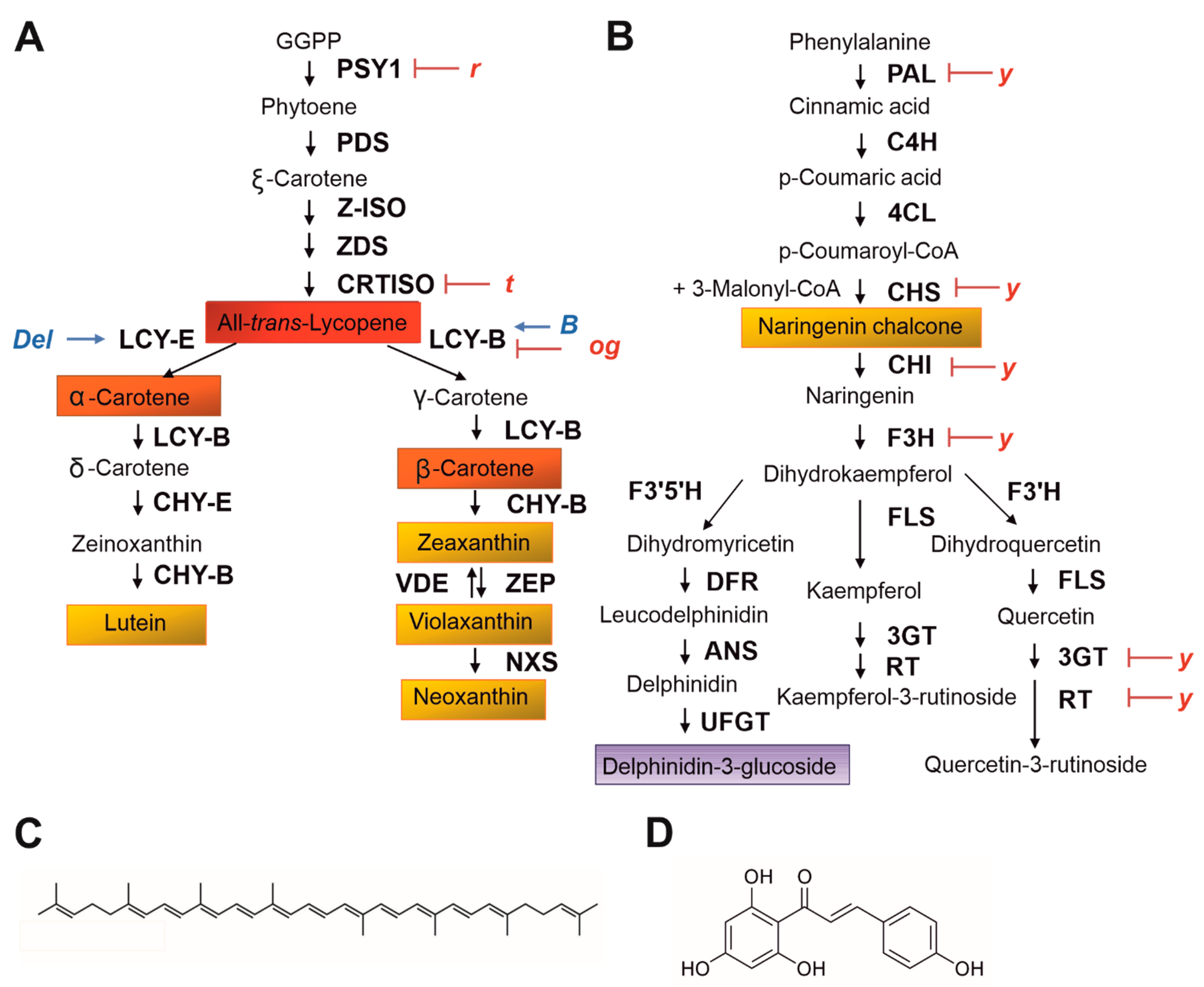
Colour represents a very flexible attribute, varying in Solanaceae from white/yellow to orange, red, purple, and brown: it mainly depends on the quantity and quality of the pigments synthesized in the fruit. These compounds belong to two different classes of secondary metabolites, carotenoids, and flavonoids, resulting from specific biosynthetic pathways, whose enzymatic steps are very conserved among the Angiosperms and well characterized in several Solanaceae (Figure 1) (for comprehensive reviews see [16,17,18,19]). While flavonoids, particularly anthocyanins, are responsible for purple, blue, and red colours, carotenoids are typical of yellow, orange, and red fruits.
Carotenoids are a class of terpenoid pigments deriving from the isoprenoid biosynthetic pathway, synthesized and accumulated in plastids [16]. They play different roles in plants, including photoprotection from oxidative damage in photosynthetic tissues and attraction of pollinators and seed dispersers when accumulated in chromoplasts of flowers and fruits. Flavonoids are polyphenolic compounds, synthesized through the phenylpropanoid pathway. They exhibit several biological activities in plants, including protection against different abiotic (especially UV radiation, low temperatures, low nitrogen) and biotic (herbivores and pathogens) stresses [18]. Anthocyanins are a special class of water-soluble flavonoids, generally accumulated into vacuoles, which confer colouration to several flowers and fruits.
The genetic diversification of the flavonoid pathway is mainly found at the regulatory level [18], while the evolution of the structural genes, encoding the enzymatic proteins acting along the biosynthetic way, was strongly constrained, particularly for those genes involved in the early reactions, common to multiple pathways. In this case, differences in fruit colour often rely on the presence or absence of specific classes of pigments, in turn depending on the specific activation or repression exerted by different transcription factors (TFs) on the pathway, or part of it. Three main types of factors, R2R3-MYB, bHLH, and WDR, take part in the so-called MBW ternary complexes, which are responsible for the transcriptional activation of several biosynthetic genes of the flavonoid pathway [17,20]. Different developmental and environmental factors can induce the transcription of the R2R3-MYB proteins, thus ultimately controlling the activation of the flavonoid pathway. At the same time, negative feedback loops, under control of both environmental or developmental factors, can be turned on through production of repressor proteins, such as R3-MYBs or hormonal signalling intermediates [20], to precisely fine-tune the process. Most of the phenotypes related to the flavonoid genetic diversification in fruits are caused by mutations affecting positive or negative regulators of the process, such as MYB or bHLH TFs. Contrary to flavonoids, carotenoid fruit colour mainly derives from the quantity of pigments produced or degraded, whose nature can depend on the activity of specific anabolic or catabolic genes or on their defect [17]. Both anthocyanins and carotenoids can be synthesised as photoprotective pigments as a response to high light conditions. Nevertheless, the two biosynthetic machineries are not overlapping. In many species, both flavonoids and carotenoids accumulate in fruit surface and/or flesh. In others, one of the two groups is predominant, and the other pathway can be attenuated or even switched off. However, at the genetic level, both are present, since no major losses in their regulatory or structural genes have been found in the genomes of this family of plants [11,17]. Due to their variety of anthocyanins, pepper and eggplant fruits have long been studied as suitable models for the anthocyanin pathway, whereas tomatoes, containing lower amounts of flavonoids, have been principally considered as a model for the carotenoid pathway.
In tomato, the fruit colour we can observe is a combination of different pigments accumulated in the epidermis, the sub-epidermal layer, and the fruit pericarp (flesh). Whereas in the pericarp of young fruits the green colour is predominant and due to the presence of chlorophylls, being the photosynthetic apparatus present and functional, a significant increase in the content of certain carotenoids occurs during ripening, while the thylakoid membranes in the chloroplasts break down and the plastids are converted into chromoplasts [21]. Consequently, fruit colour progressively changes from green to yellow, orange, and red, when different carotenoids and flavonoids are synthesized, and lycopene, the main carotenoid in red mature fruits, is finally accumulated (Figure 1A,C). The change in colour progressively occurs between the mature green stage, when all the plastids are still chloroplasts, and the fully ripe stage, when only developed chromoplasts are present [21]. This colour change is intimately interconnected with developmental signals, among which ethylene production plays a key role. Tomato is a climacteric fruit, and a production of high levels of ethylene is observed at the onset of ripening. Many of the carotenoid biosynthetic genes are regulated by this hormone [22] and thus participate in the general transcriptional regulation of the fruit ripeness, which is essentially driven by ethylene. Important mediators in this process are some TFs belonging to the MADS-box gene family [17], which, in plants, is involved in the regulation of many different developmental processes. The master regulator of the ripening cascade in tomato fruit is the Ripening Inhibitor (RIN) gene, encoding a MADS-box TF which can interact with the promoter of several genes involved in ethylene synthesis and transduction, cell wall modifications, and carotenoid biosynthesis, thus synchronising the ripeness cascade with the initiation of carotenoid production [23]. Other MADS-box TFs are involved in the regulation of the tomato fruit ripening [17], with possible influence on carotenoid production: some of them will be described in the following paragraphs.
Differently from carotenoids, mostly accumulated in tomato fruit pericarp, flavonoids (Figure 1B) are mainly synthesized in the single-cell layer epidermis, which is normally yellow due to the accumulation of naringenin chalcone (Figure 1D) from the breaker stage onward. This flavonoid likely plays an important role against fruit desiccation and offers photoprotection to the underlying photosynthetic tissues [24]. Anthocyanins are generally not present in tomato fruits, but when synthesized belong to the delphinidin-type (Figure 1B), and are also accumulated in the epidermis and in the sub-epidermal layer.
1.3. Phylogenetic Reconstruction and Human Selection of Fruit Colour in Tomato
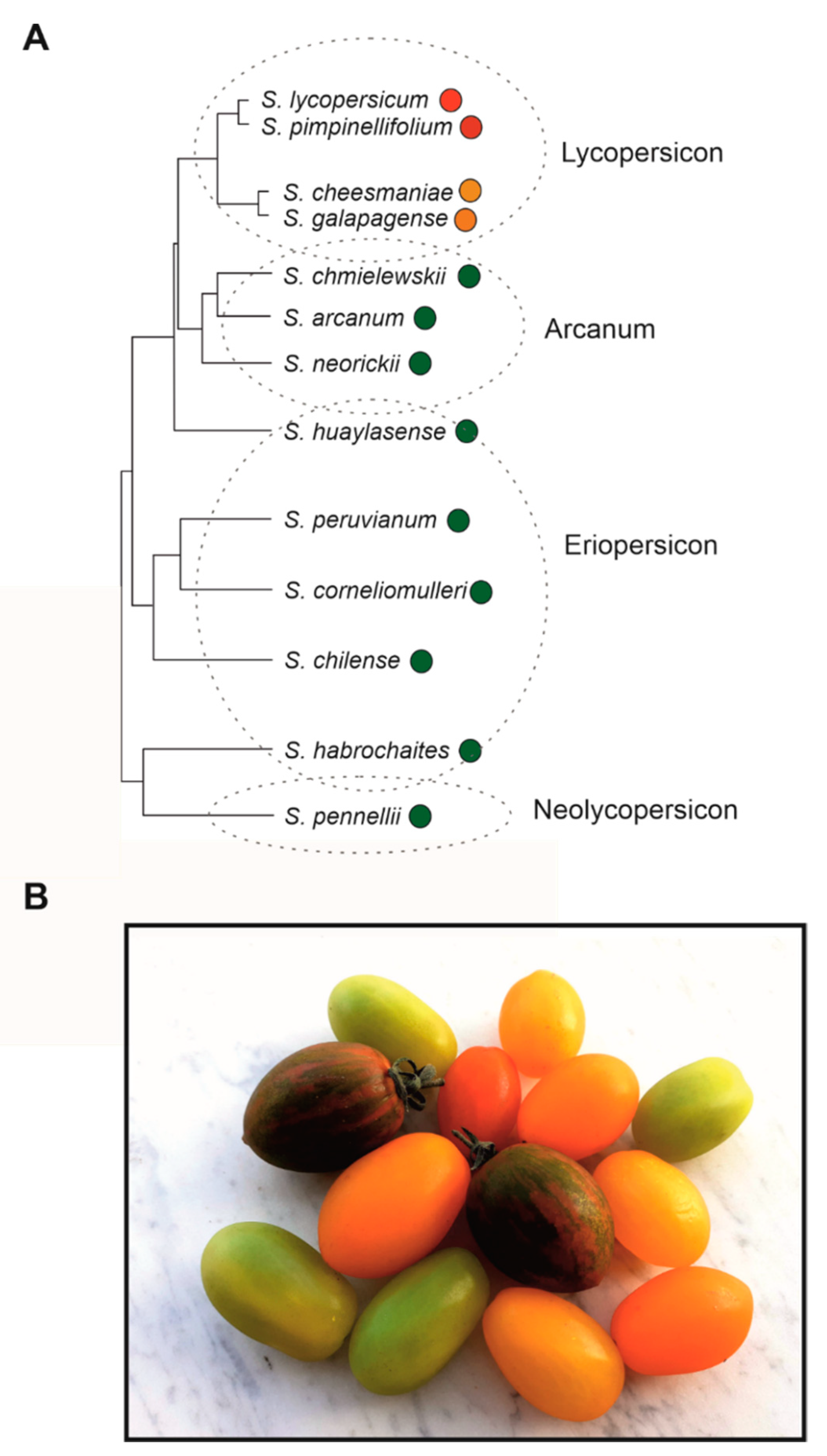
Cultivated tomato varieties belong to the species S. lycopersicum var. lycopersicum (SLL), which originated in South America and was first domesticated in Mexico and Perú (likely in separate phases, [25]), before being introduced in Europe in the 16th century and finally, from the 19th century onwards, an object of important breeding programs in Europe and America [11]. The first described tomato fruits introduced in Europe showed some variability in colour, shape, and size, and yellow and red were described as two possible colours [26]. SLL belongs to the so-called “tomato clade”, a group of thirteen Solanum relative species, further divided in four subgroups [27,28] (Figure 2A). SLL is included in the section Lycopersicon, a restricted group of tomato close species, all self-compatible, to which Solanum pimpinellifolium (SP) also belongs, considered the closest relative of SLL, which produces red fruits (Figure 2A). Red fruits are also produced by the var. cerasiforme of S. lycopersicum (SLC), regarded as an intermediate between SP and SLL and the direct ancestor of the cultivated varieties due to its close genetic relationship with them [13,25]. Solanum galapagense and Solanum cheesmaniae are both endemic to the Galapagos Islands, and produce small orange-yellowish fruits [29,30]. They also belong to the section Lycopersicon (Figure 2A). In this subgroup of the tomato clade, red and orange carotenoids represent the main fruit pigments. The orange colour shown by the Galapagos Islands species, later introgressed in SLL, is due to a high accumulation of β-carotene, de-novo produced from trans-lycopene thanks to a high expression of the dominant BETA (B) allele of a chromoplast specific lycopene β-cyclase (LCY-B) gene, which in wild type tomato is generally poorly transcribed [31,32] (Figure 1A). On the contrary, the old-gold (og) recessive allele of LCY-B induces higher levels of trans-lycopene production and leads to dark red fruits [31] (Figure 1A). Other two different “orange” alleles identified in SLL are the Delta (Del) dominant allele, introgressed from the wild species Solanum pennellii, which induces increased expression of the lycopene δ-cyclase (LCY-E), resulting in increased transformation of lycopene in δ-carotene [31,32], and the tangerine (t) allele, a mutated allele of the carotene isomerase (CRTISO) gene, which leads to the main accumulation of cys-lycopene and other carotenoids, at the expense of the red trans-lycopene [32,33,34] (Figure 1A). Yellow-coloured tomato fruits are due to recessive alleles of the yellow flesh (r) locus, which correspond to mutations in the phytoene synthase 1 (PSY1) gene, encoding the first committed enzyme in the carotenoid biosynthetic pathway [32,35] (Figure 1A). In this case, the yellow colour is due to the flavonols present in the fruit peel and to other carotenoids, such as lutein, which are normally present in green tissues. The brown colour of some tomato fruits, exhibited especially by some heirloom tomatoes, is instead dependent on the superimposition of the red lycopene colouration with the green colour of the chlorophylls, which, usually degraded during tomato ripening, are here retained because of the inhibition of their degradation.
Figure 2.
(A) Phylogenetic tree of Solanum species belonging to the tomato clade, divided in the four subgroups identified by [27,28]. Redrawn with simplifications from [29], with the addition of the fruit colour, represented by coloured circles close to the species names. (B) Photo of a group of ripe Solanum lycopersicum fruits exhibiting possible colour variants in the carotenoid/chlorophyll pathway: red, orange, yellow, green, and “green stripe”-like colourations are shown.
Figure 2.
(A) Phylogenetic tree of Solanum species belonging to the tomato clade, divided in the four subgroups identified by [27,28]. Redrawn with simplifications from [29], with the addition of the fruit colour, represented by coloured circles close to the species names. (B) Photo of a group of ripe Solanum lycopersicum fruits exhibiting possible colour variants in the carotenoid/chlorophyll pathway: red, orange, yellow, green, and “green stripe”-like colourations are shown.

The phenotype is called green flesh (gf) and is due to different types of mutations identified in a tomato homolog of the staygreen (SGR) gene of rice, which encodes a chloroplast protein regulating the degradation of chlorophyll during senescence [36,37]. All these different genotypes thus originated a wide panel of different carotenoid colourations of tomato fruits (Figure 2B). Other peculiar tomato fruit colours originate from variants in the flavonoid pathway, such as the pink colour associated with the locus y: this is caused by a mutation in the master regulatory factor SlMYB12, with the downregulation of the expression of several structural genes of the flavonoid pathway and the absence of the yellow-coloured flavonoid naringenin chalcone in the fruit peel, which remains transparent [13,38]. By combining the loci y and r in the same genotype an almost white tomato fruit can be produced, characterized by a very pale-yellow flesh [11]. When the SGR gene is also mutated in a y x r background, tomato fruits remain green, since they do not contain carotenoids and flavonoids and cannot degrade chlorophylls during ripening [39].
Naringenin chalcone (Figure 1D) represents the main flavonoid synthesized in tomato fruits, together with lower amounts of the flavonols quercetin-3-rutinoside (rutin) and kaempferol-3-rutinoside (Figure 1B). Other trace glycosides of naringenin chalcone, quercetin, and kaempferol have been identified [38], but the phenylpropanoid biosynthetic pathway is blocked downstream and does not allow the production of other important classes of polyphenols, including anthocyanins. Nevertheless, wild related Solanum species can produce anthocyanins in their fruit peel (Figure 3A), and this trait was repeatedly introgressed into SLL through interspecific crosses [40]. One of the most studied anthocyanin-enriched phenotypes was Anthocyanin fruit (Aft), derived from interspecific cross with Solanum chilense [40,41,42] (Figure 3B). A similar phenotype, named Aubergine (Abg) [42], was introgressed from Solanum lycopersicoides. An even more marked anthocyanin pigmentation was obtained in the Aft or Abg tomato lines with the inclusion of the atroviolacea (atv) locus, introgressed from S. cheesmaniae [40,42] (Figure 3B,C). All these anthocyanin phenotypes are controlled by genes inherited from wild Solanum species, whose fruits in nature are commonly green, but are also able to accumulate purple anthocyanins in specific environmental conditions [30] (Figure 3A). Excluding the restricted group of Solanum closest relatives above described, the green fruit phenotype is found in all the other subgroups of the tomato clade, from Solanum chmielewskii to S. pennellii (Figure 2A). These are all species characteristic of the Andean regions of South America and the geographic distribution, particularly the altitude of their typical habitats, may explain the ability they developed and maintained to produce, when necessary, secondary metabolites allowing photoprotection and/or protection of the fruit from low temperatures.
In addition to genes involved in carotenoid and flavonoid synthesis and in chlorophyll degradation, other genes have been shown to influence tomato fruit colour. Mutations in negative regulators of the upstream light-signalling pathways which regulate both carotenoid and flavonoid biosynthesis, such as the high pigment (hp) mutations [43,44], increase the amount of all the pigments normally produced under light stimulation (carotenoids, flavonoids, and chlorophylls). hp tomatoes thus show a darker colour of both peel and flesh. Other regulators of fruit colour are some TFs involved in fruit ripening: besides the already mentioned MADS-box TF RIN [23,45], it is worth to remember the SBP-box TF Colourless Non-Ripening (CNR) [46], and the NAC TF Non-Ripening (NOR) [47], as well as some components of the ethylene signalling pathway, such as Never Ripe (NR) [48] and Ethylene Insensitive2 (EIN2) [49,50,51].
Going backwards in the phylogeny of SLL, it appears that the red colour, typical of the fruits produced by SP, likely represented the original colour of the first domesticated tomato plants and it is still the most typical of the modern varieties (Figure 2A). Red colour is also shown by the fruits of the several cultivated landraces of SLC, which in South America still experience occasional gene flows with the wild relative SP [13,52]. Spontaneous mutations in the carotenoid pathways may have occurred on several occasions during tomato cultivation and some of them, such as the yellow flesh, even in the original areas of tomato domestication, since this colour was already present in some tomato genotypes described in Europe in the 16th century [53]. Other colours, such as the orange or brown, which are typical of heirloom accessions worldwide spread, may have appeared in similar ways.
Genomic analyses into the history of tomato domestication identified the principal traits under selection in fruit size and taste: fruit mass, in particular, was the key character selected by men [13]. However, studying tomato domestication and breeding, structural variants also affecting the expression of genes involved in lycopene metabolism were recently detected. They would indicate reduction of lycopene from SP to SLC and subsequent settlement in both the heirloom and modern populations of the big-fruited SLL tomato, suggesting that the trait could have been under selection during domestication but not in the more recent phases of tomato improvement and modern breeding [54]. A reduction in the level of β-carotene was also highlighted in the transition of SP to SLC in the South American areas of domestication, together with tendencies toward larger fruit size, thicker pericarps, higher number of locules, lower content of citric acid, and of soluble solids, all considered typical traits of tomato domestication syndrome [25].
The green colour of fruits (Figure 2A), together with allogamy and self-incompatibility, thus represents a primitive trait in the tomato clade [27]. A recent study identified functional PSY1 enzyme, with similar catalytic activity, in both green-fruited and red-fruited Solanum species [55]. Therefore, it seems that a major difference in PSY1 expression, probably due to important changes in PSY1 promoter/5′-UTR regions, with the loss of regulatory elements involved in abiotic stress response and the acquisition of new ethylene-responsive elements, was important in the association of carotenogenesis with ripening, thus contributing to pericarp colour variation from green to red during tomato domestication [55]. Concomitantly with this, red-fruited tomato species lost the ability to synthesize anthocyanins, possibly because of differential climatic adaptations or the same domestication process.
In addition to the effects of spontaneous mutations, diversity in fruit colour existing in SLL varieties may be ascribed to new alleles introgressed from wild tomato genomes into modern cultivars. Changes in fruit colours may have also occurred as a secondary consequence of breeding strategies aimed at improving other traits. More recently, thanks to the increasing awareness of the beneficial effects on human health of certain classes of secondary metabolites, colour has become a major determinant of the economic value of tomato: therefore, enrichment of its fruits with specific groups of pigments, such as anthocyanins and other flavonoids, has become a novel goal of genetic engineering or breeding. These approaches led to the production of high-flavonol [56] and high-anthocyanin [40] tomatoes (Figure 3C), further increasing the variability of the tomato surface pigmentation.
2. Novel Genetic Mechanisms in the Regulation of Pigment Biosynthesis in Tomato Fruits
2.1. Alternative RNA Splicing and Epigenetic Modifications May Modulate Carotenoid-Derived Colour in Tomato Fruits
Spontaneous mutations affecting tomato fruit carotenogenesis have been described since the beginning of the last century, and many of them were identified in their genomic determinants in the last decades. As previously described, yellow and orange represent possible variants of the more common red colour in tomato fruits. These variants originated several times, either spontaneously or through introgression breeding, because of mutations in one or more genes involved in the carotenoid pathway (Figure 1A). Recently, a series of new genotypes characterized by yellow fruit colour, still elicited by different post-transcriptional mechanisms of gene regulation, were described. These mechanisms involve a series of processes which act at RNA processing, stability, or translation levels. Alternative RNA splicing indeed emerged as an important additional way to fine-tune protein production, produce new protein isoforms, and create novel mechanisms for mutations to affect expression of genes involved in pigment biosynthetic pathways, thus providing further evolutionary flexibility to the colour trait.
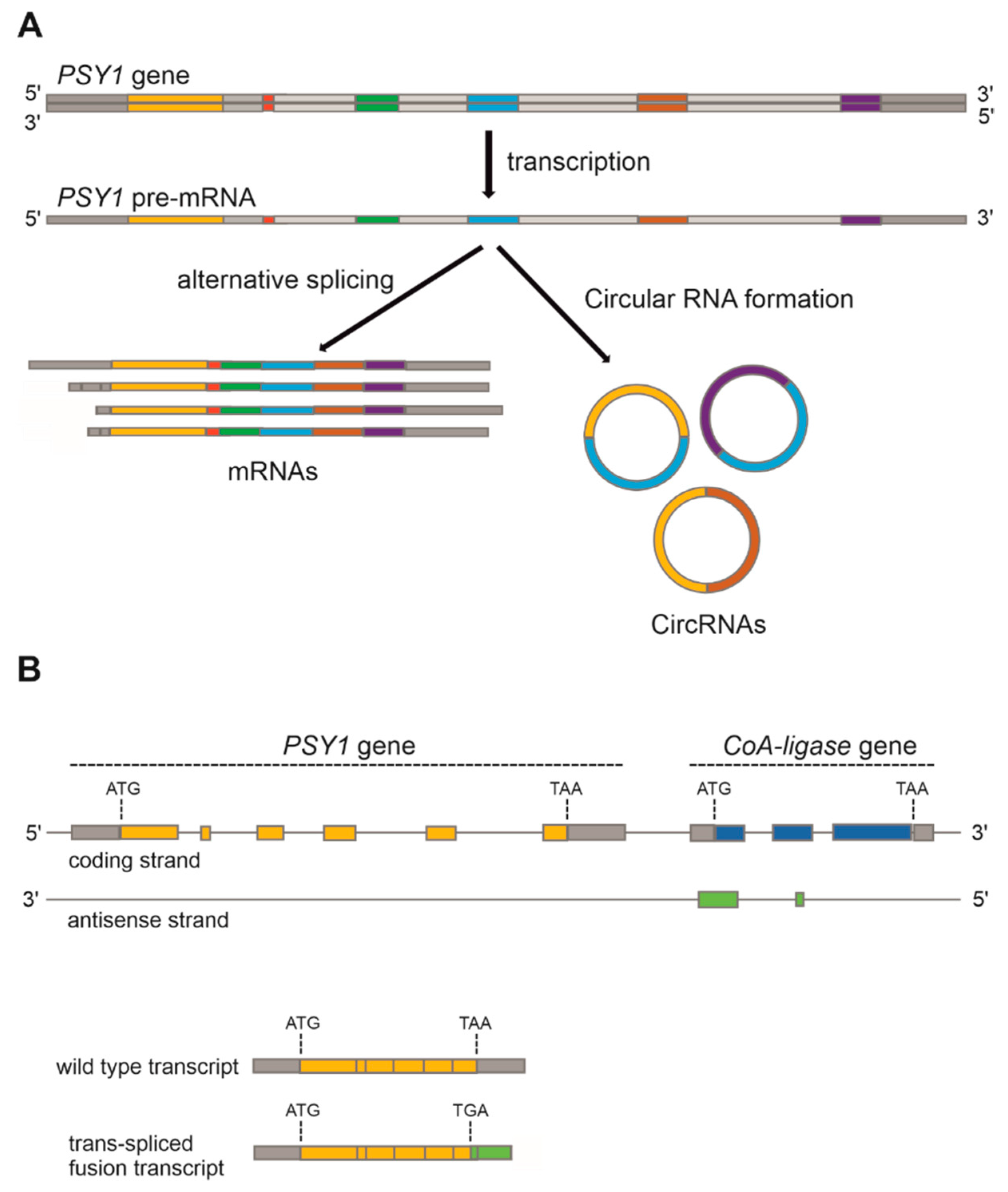
Alternative splicing (AS) is the process of splicing the exons of the primary transcript of a gene in different arrangements: this allows the production of structurally and functionally distinct mRNAs which can lead to protein variants or may reduce the abundance of the canonical mRNA splicing form, thus leading to a reduction or a suppression of the biosynthetic pathway in which the relative protein is involved [57]. This mechanism and its possible effects on fruit colours have been recently described in several species, including Citrus sinensis [58], Triticum aestivum [59], Arabidopsis thaliana [60], and tomato [61]. Most of the studies carried out in SLL focused on the AS of the PSY1 gene, which encodes the only phytoene synthase of tomato expressed in fruits. This enzyme catalyses the first committed step in the carotenoid biosynthesis (Figure 1A) and is therefore crucial for carotenogenesis: its suppression is sufficient to block the entire pathway, resulting in yellow fruits. Recently, in a genomic survey of AS patterns in tomato seedlings, flowers, and young developing fruits, AS of PSY1, leading to alternative splice variants (ASVs) during fruit early development, has been identified [61] (Figure 4A); however, the functional meaning has still to be elucidated. As was found in other plants, for example in wheat endosperm [59], it is possible that the production of ASVs reduces the mRNA abundance of the canonical one, contributing to a decrease of carotenoid biosynthesis.
Another emerging category of gene expression regulatory mechanisms is the one mediated by Circular RNAs (CircRNAs). These are covalently closed loop, single-stranded transcripts that are ubiquitously expressed in all eukaryotes and are increasingly thought to be involved in regulation of transcription, modulation of AS, and binding of microRNAs and proteins [62]. In another recent survey carried out in tomato fruits, several CircRNAs transcribed from pigment biosynthetic genes were discovered, some of them appearing to be regulated by ripening [63]. In particular, CircRNAs from PSY1, Phytoene Desaturase (PDS), and 15-cis-zeta-carotene isomerase (Z-ISO) genes, all involved in carotenoid biosynthesis (Figure 1A), resulted to be up-regulated during fruit ripening, exhibiting expression profiles similar to those of their parent genes. From PSY1, three classes of Circular RNAs were identified, indicating that alternative circularization events may occur on this gene (Figure 4A). The overexpression of one of these variants showed correlation with a reduced transcription of PSY1 in yellow fruited transgenic plants, showing that the continuous high expression of PSY1 CircRNAs may inhibit their parent mRNA accumulation. The production of CircRNAs from carotenoid biosynthetic genes could therefore represent an additional mechanism to modulate the expression levels of their most abundant mRNAs, fine-tuning the enzymes catalysing specific biosynthetic steps.
A particular mechanism of RNA splicing which has been identified in tomato thanks to the analysis of a specific fruit phenotype is “trans-splicing”, which produces chimeric transcripts by joining mRNAs derived from two different genes located on the same or different chromosomes [64]. This mechanism of alternative RNA splicing still scarcely described in plants, requires, like “cis-splicing”, canonical splice sites (GU-AG) between the 5’ splice donor of the pre-mRNA of the first gene and the 3’ splice acceptor of the pre-mRNA of the second gene. This process was sometimes explained as a by-product of cis-splicing, because trans- and cis-splicing are generally found to occur in parallel [65]. However, trans-splicing might be another form of AS with regulatory functions. In tomato, a yellow fruit phenotype was found to be caused by loss-of-function of PSY1 induced by the contemporary production of hybrid transcripts through the trans-splicing fusion of the transcript of PSY1 with a portion of the adjacent gene encoding the Acyl-CoA synthetase (ACoS), transcribed on the neighbour opposite DNA strand (Figure 4B). This peculiar AS event was first described in two yellow fruited SLC accessions [66,67] and then also in a cherry SLL genotype [68]. In all these lines, a SNP in the fourth intron of PSY1 and a simple sequence repeat (SSR) in the intergenic region between the two adjacent genes appeared associated with the trans-splicing process [66]; however, the only intronic SNP resulted in not being sufficient to induce it [69]. Interestingly, the sequence fused to PSY1 transcript resulted almost identical to a previously identified long non-coding RNA (lncRNA) which was named ACoS-AS1. By analysing several yellow fruited tomato lines, three possible alternative PSY1-ACoS-AS1 trans-spliced transcripts were isolated, indicating the existence of possible variants of the trans-splicing event [69]. The transcription of a wild type ACoS-AS1 sequence resulted essential to produce trans-splicing between PSY1 and ACoS-AS1 transcripts. Finally, the sub-cellular localization analysis showed that the chimeric protein PSY1-ACoS-AS1 could not enter plastids where PSY1 plays its enzymatic activity [69]. This, coupled with the reduced expression of the canonical PSY1 transcript, may explain the yellow phenotype of this group of mutants.
The nature and the abundance of pigments in tomato fruits may also be affected by epigenetic mechanisms modifying patterns of gene expression. Several different mechanisms of epigenetic regulation exist in plants [70]; some involve cytosine methylation and histone modification of DNA, which in turn can affect TFs’ binding affinity or recruitment of chromatin remodelling proteins with consequent alteration of gene expression. An epigenetic change in a genomic region encompassing the CNR locus, for example, is responsible of a general inhibition of tomato fruit ripening with the concurrent development of a colourless fruit pericarp, because of incapacity to synthesise carotenoid precursors [46]. The SQUAMOSA promoter binding protein-like (SPL) gene, belonging to the SBP-box gene family of TFs, was identified at the CNR locus and the mutated phenotype resulted from a naturally occurring and heritable hypermethylation of the promoter region, which reduces SPL gene expression with the consequent inhibition of a subset of downstream processes involved in normal fruit development and ripening [46]. Another exemplary case study is the “green stripe” fruit colour. The GREEN STRIPE tomato is a natural mutation characterized by nonuniform pigmentation of the fruit peel, with irregular longitudinal green stripes (GS) alternated to light green stripes (LGS) in the epidermis of unripe fruits. This phenotype is controlled by the green stripe (gs) locus, described as a single recessive gene [71] linked to chromosome 7 [72], and is due to a longer retention of chlorophylls in GS during ripening (Figure 2B). In fully ripe fruit, the GS may assume a paler red colour than the other parts of the epidermis, or a yellow colour. In a recent study [73], this phenotype was found to be associated with the methylation level of the promoter of the Tomato Agamous-Like1 (TAGL1) gene, which encodes a MADS-box TF known to affect chloroplast development. The promoter methylation was found to be associated to a specific SNP in the second intron of the same gene. High degrees of methylation of TAGL1 promoter led to downregulation of TAGL1 expression, associated with GS phenotype. Conversely, low degrees of methylation increase the expression of TAGL1, leading to LGS phenotype. TAGL1 expression can in turn affect the transcription of different genes: in particular, it downregulates some genes associated with chloroplast accumulation in immature fruit, and upregulates the expression of PSY1, thus acting as a positive regulator of carotenoid biosynthesis in ripe fruit [74]. The mechanism underlying TAGL1 promoter methylation is as ever-complex. In plants, de novo DNA methylation to specific genomic DNA sequences occurs via a peculiar pathway called “RNA-directed DNA methylation” or RdDM, which involves non-coding RNA molecules such as short interfering RNAs, scaffold RNAs (lncRNAs), and a range of proteins [75]. It was hypothesized that the second intron of TAGL1 may be transcribed into lncRNAs, as already observed in Arabidopsis thaliana, where from the second intron of Agamous gene (which is the homologous of tomato TAGL1) three functional lncRNAs are produced [76]. Because of the SNP, the lncRNAs transcribed from the second intron of TAGL1 may be altered in their sequence, resulting in a different ability to take part to the RdDM pathway, and finally influencing the degree of TAGL1 promoter methylation. In addition to this mechanism of epigenetic regulation, AS of TAGL1 was also observed, with production of alternative RNA transcripts in GS and LGS at different levels: this may also contribute to differentiate TAGL1 expression in the different coloured areas of the fruit peel.
Carotenoid production, as already explained, is a process which is tightly regulated in the broader context of tomato fruit ripening: ethylene perception, for example, is necessary to induce expression of several carotenoid biosynthetic genes in fruit pericarp [22]. Thus, hormonal and transcriptional regulators play a central role in the complex dynamics of fruit maturation, including proper colour development, and an increasing number of studies indicate how epigenetic mechanisms, particularly those controlling the DNA methylation status, represent a key factor interconnected with both hormones and TFs modulation [77]. A general targeted DNA demethylation indeed appears to be an important dynamic component in the control of tomato fruit ripening; the epigenetic mutations in ripening TFs, such as those reported in CNR-SPL or TAGL1, can then further modulate the epigenome’s influence over ripening [77].
2.2. Splicing Mutations and Alternative Transcripts Can Affect Anthocyanin Synthesis in Tomato Fruit Peel
As occurred in carotenoid biosynthesis, AS and epigenetic modifications of genes involved in the regulation of anthocyanin pathway can influence the nature and the accumulation of these pigments as well. Thanks to the increasingly common transcriptome analyses, the AS landscape and dynamics correlated with both flower and fruit anthocyanin colouration are more and more extensively characterized in plants. What clearly appears is an important role of these post-transcriptional gene regulatory mechanisms in the modulation of flavonoid metabolism and anthocyanin biosynthesis. This has been recently described in several species, including tea plant (Camellia sinensis) [78], Brassica napus [79], and chrysanthemum (Chrysanthemum morifolium) [80], where either MYB or bHLH regulatory genes were found to undergo AS to fine-tune anthocyanin production in specific tissues.
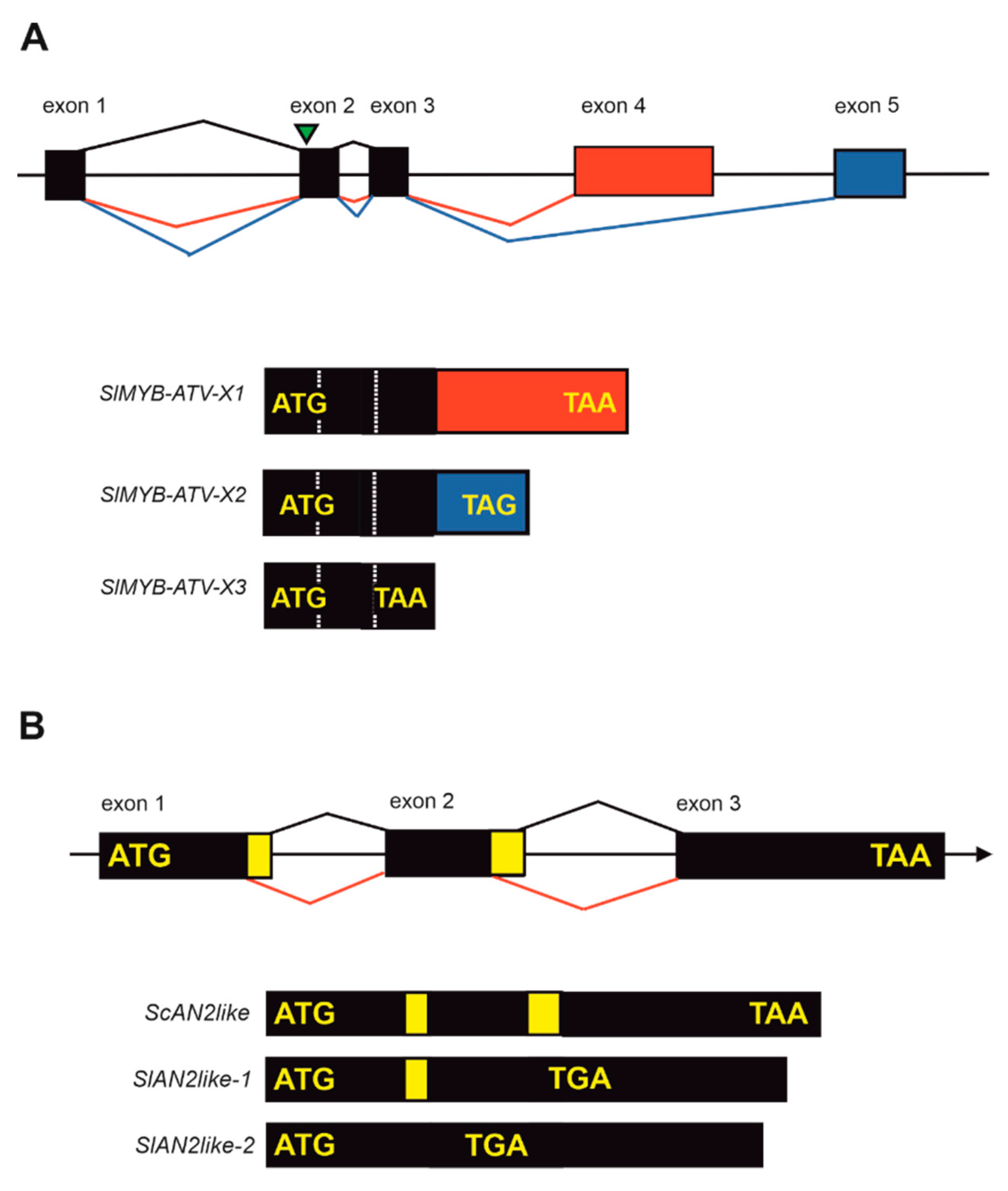
In tomato fruits, similar mechanisms are still scarcely described. However, some interesting cues are beginning to emerge as the new flavonol- and anthocyanin-enriched fruit phenotypes are studied at the genetic level. Two different MYB factors, introgressed from wild tomato relatives and involved with opposite roles in the ectopic anthocyanin pigmentation of the fruit peel, have recently been shown to be subject to post-transcriptional regulation. The first one was the atv locus, introgressed from S. cheesmaniae: it can increase anthocyanin pigmentation in vegetative tissues of tomato under high light conditions, and, when expressed with one of the loci Aft or Abg, strongly enhances the fruit peel anthocyanin pigmentation, leading to the so-called “purple” tomatoes [40,81] (Figure 3B,C). Genetic and molecular analyses identified the gene SlMYB-ATV, encoding an R3-MYB factor, as responsible for the phenotype [82,83]. The SlMYB-ATV protein negatively interferes with the activation of the anthocyanin biosynthetic pathway mediated by the endogenous MBW complexes, acting as a competitive inhibitor of the R2R3-MYB activating TFs, through direct binding with the bHLH factors [83]. Interestingly, either the wild type or the mutated SlMYB-ATV alleles undergo AS and produce three different transcripts [82] (Figure 5A), whose functional meaning is still to be elucidated. When transcribed from the wild type gene, these transcripts are translated in polypeptides characterized by different lengths, but all displaying the R3-bHLH binding domain and thus putatively able to interfere with the MBW complexes. While two of them are shorter and do not include long C-terminal domains after the R3 motif, the third one includes a longer sequence downstream of the MYB domain. Interestingly, this transcript, which is also the most different from the predicted mature mRNA produced by canonical splicing, appeared two orders of magnitude more expressed than the other two in fruit peel, and therefore can be considered as the most relevant in this organ [82]. In the atv genotype, a 4-nucleotide insertion in the second exon, which is common to all the three transcripts, produces a premature stop codon which results in a truncated protein missing the R3-bHLH binding domain [82,83]. It will be interesting to understand if the three ASVs of SlMYB-ATV may produce alternative forms of the repressor protein which, in wild type plants, act in different tissues or under specific environmental signals. It is also possible that the SlMYB-ATV mRNA most expressed in fruits, which includes the long C-terminal domain, may exert a stronger repression on the anthocyanin pathway, possibly involving direct binding with the promoter region of some structural or regulatory genes.
The other MYB protein whose alternatively spliced transcripts were isolated and characterized is the R2R3-MYB TF identified as the master positive regulator of the anthocyanin pigmentation in tomato fruit peel. For a long time, it was believed that the default state of cultivated tomato was the lack of anthocyanin pigmentation in the fruits. This was supposed to be caused by mutations affecting the expression of structural biosynthetic genes, such as the one encoding the chalcone isomerase (CHI) enzyme (Figure 1B), whose substrate, naringenin chalcone, is indeed the main flavonoid intermediate compound accumulating in the peel [84] (Figure 1D). Through the introgression of delimited chromosomic regions from wild tomato species into the S. lycopersicum genome, such as in the Aft accession (Figure 3B), it was possible to induce a purple-spotted anthocyanin pigmentation in tomatoes’ skin [41,42]. Recently, the locus responsible for such a phenotype was identified [85,86,87]: it corresponds to an R2R3-MYB encoding gene, named SlAN2like, which is not functional in cultivated tomato due to a splicing mutation affecting an AC splice site in the second intron [85]. A second possible splicing mutation, leading to an even shorter transcript, was also found in wild type fruits [86]. Due to these mutations, the gene in SLL is alternatively spliced, producing aberrant transcripts with premature stop codons (Figure 5B) which likely induce a nonsense mediated decay, or, if translated, lead to truncated and non-functional polypeptides, lacking the R3 domain with the bHLH-interaction aminoacidic signature [85,86]. On the contrary, in Aft tomato the AN2like allele, introgressed from S. chilense (Figure 3A), is functional: it undergoes canonical splicing and produces an active R2R3-MYB TF (Figure 5B) which, in turn, can trigger the anthocyanin biosynthetic pathway by taking part to the MBW complexes [85,86,87]. This recent evidence revealed that in SLL the inability to synthesize anthocyanins in fruit peel was due to mutations in regulatory rather than in structural genes. As described in a previous paragraph, since red fruits are also characteristic of the closest tomato relatives SP and SLC, it is conceivable that the splicing mutation in the AN2like gene occurred immediately before or in the first stages of tomato domestication [86,88]. Further in-depth analyses of the chromosome 10 region, which includes AN2like as well as other three functional R2R3-MYB genes [86,89], in SLL and in other species of the Solanum clade, including both close relatives of SLL and more distant wild species, may help to piece together the evolution of this important genomic region, whose structure and function are major determinants of anthocyanin synthesis in tomato fruits.
3. Conclusions
The fruit colour is a complex phenotypic trait that can be modelled by different factors: environmental elements, phylogenetic constraints and, more than others, coevolution with seeds dispersers’ preferences. Tomato fruits experienced a profound change in most of their attributes as domestication progressed. From the small, red, and round fruits produced by SP plants, edible but still considered wild, major changes occurred in size, taste, and scent in SLC and SLL fruits, as a consequence of the selection carried out by humans. From this point of view, colour appears as a quite stable trait, since most of the modern tomato varieties still bear red fruits. Actually, the big change in fruit colour seems to have occurred earlier, from the green-fruited to the red-fruited species of the tomato clade, then before domestication or at the very early history of that. Another important step in the evolution of this trait was the loss of the capacity to synthesize anthocyanins in the fruit peel, which presumably occurred again in the passage from the green-fruited to the red-fruited species. Recent studies indicate that an important step in the acquisition of the red colour was the activation of the carotenogenesis in tomato fruits thanks to the progressive increase of the expression of PSY1, the key biosynthetic gene of the carotenoid pathway in fruits, through the modification of important regulatory elements of its promoter region. The loss of anthocyanins was instead due to knock out mutations in the gene encoding the key regulatory TF, activating their biosynthetic pathway in fruit peel.
Beyond the evolution of the major determinants of fruit colour in tomato, a common feature that is clearly emerging from the most recent studies is the complexity of the mechanisms that underlie the expression of this trait. Transcriptional, post-transcriptional, and epigenetic mechanisms affecting the expression of an increasing number of regulatory and structural biosynthetic genes appear to cooperate to finally modulate the abundance of the functional proteins operating in the different pathways. If these mechanisms appear multiple and intertwined with each other, they offer an even more complex scenario for possible alterations, further increasing the possibility to modulate the system. As for other important physiological characters, recent studies demonstrate that tomato fruits can represent a model for the genetic dissection of the colour determinants, and as the knowledge of the novel molecular mechanisms controlling gene expression advances, it is plausible that new discoveries will be made in the next years. This will probably allow further modification to tomato colour trait, according to both aesthetic and nutritional criteria, to fully meet human needs.
Author Contributions
S.G. conceived and wrote the manuscript. P.P. provided critical reviewing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This review was funded by the Scuola Superiore Sant’Anna of Pisa, Italy.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
Conflicts of Interest
The authors declare no conflict of interest.
References
- Willson, M.F.; Whelan, C.J. The Evolution of Fruit Color in Fleshy-Fruited Plants. Am. Nat. 1990, 136, 790–809. [Google Scholar] [CrossRef]
- Valenta, K.; Kalbitzer, U.; Razafimandimby, D.; Omeja, P.; Ayasse, M.; Chapman, C.A.; Nevo, O. The evolution of fruit colour: Phylogeny, abiotic factors and the role of mutualists. Sci. Rep. 2018, 8, 14302. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Fritsch, P.W.; Matzke, N.J.; Wang, H.; Kron, K.A.; Li, D.-Z.; Wiens, J.J. Why is fruit colour so variable? Phylogenetic analyses reveal relationships between fruit-colour evolution, biogeography and diversification. Glob. Ecol. Biogeogr. 2019, 28, 891–903. [Google Scholar] [CrossRef]
- Sinnott-Armstrong, M.A.; Donoghue, M.J.; Jetz, W.J. Dispersers and environment drive global variation in fruit colour syndromes. Ecol. Lett. 2021, 24, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.E.; Chapman, C.A. Frugivores and fruit syndromes–Differences in patterns at the genus and species level. Oikos 1993, 66, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Jordano, P. Angiosperm fleshy fruits and seed dispersers: A comparative analysis of adaptation and constraints in plant-animal interactions. Am. Nat. 1995, 145, 163–191. [Google Scholar] [CrossRef] [Green Version]
- Stournaras, K.E.; Lo, E.; Böhning-Gaese, K.; Cazetta, E.; Dehling, D.M.; Schleuning, M.; Caswell Stoddard, M.; Donoghue, M.J.; Prum, R.O.; Schaefer, H.M. How colorful are fruits? Limited color diversity in fleshy fruits on local and global scales. New Phytol. 2013, 198, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Lomascolo, S.B.; Levey, D.; Kimball, R.T.; Bolker, B.M.; Alborn, H.T. Dispersers shape fruit diversity in Ficus (Moraceae). Proc. Natl. Acad. Sci. USA 2010, 107, 14668–14672. [Google Scholar] [CrossRef] [Green Version]
- Galetti, M.; Guevara, R.; Côrtes, M.C.; Fadini, R.; Von Matter, S.; Leite, A.B.; Labecca, F.; Ribeiro, T.; Carvalho, C.S.; Collevatti, R.G.; et al. Functional extinction of birds drives rapid evolutionary changes in seed size. Science 2013, 340, 1086–1090. [Google Scholar] [CrossRef] [Green Version]
- Nevo, O.; Valenta, K.; Kleiner, A.; Razafimandimby, D.; Jeffrey, J.A.J.; Chapman, C.A.; Ayasse, M. The evolution of fruit scent: Phylogenetic and developmental constraints. BMC Evol. Biol. 2020, 20, 138. [Google Scholar] [CrossRef]
- Paran, I.; van der Knaap, E. Genetic and molecular regulation of fruit and plant domestication traits in tomato and pepper. J. Exp. Bot. 2007, 58, 3841–3852. [Google Scholar] [CrossRef] [Green Version]
- Zeller, U.; Göttert, T. The relations between evolution and domestication reconsidered–Implications for systematics, ecology, and nature conservation. Glob. Ecol. Conserv. 2019, 20, e00756. [Google Scholar] [CrossRef]
- Lin, T.; Zhu, G.; Zhang, J.; Xu, X.; Yu, Q.; Zheng, Z.; Zhang, Z.; Lun, Y.; Li, S.; Wang, X.; et al. Genomic analyses provide insights into the history of tomato breeding. Nat. Genet. 2014, 46, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Zhao, J.; He, C. Evolutionary developmental genetics of fruit morphological variation within the Solanaceae. Front. Plant. Sci. 2015, 6, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, C.; Diretto, G.; Giuliano, G. Biosynthesis and engineering of carotenoids and apocarotenoids in plants: State of the art and future prospects. Biotechnol. Genet. Eng. Rev. 2010, 26, 139–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, M.K.; Sharma, R.; Koul, A.; Kaul, S. Development of fruit color in Solanaceae: A story of two biosynthetic pathways. Brief. Funct. Genom. 2015, 14, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Tohge, T.; Perez de Souza, L.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef]
- Li, Z.; Vickrey, T.L.; McNally, M.G.; Sato, S.J.; Clemente, T.E.; Mower, J.P. Assessing Anthocyanin Biosynthesis in Solanaceae as a Model Pathway for Secondary Metabolism. Genes 2019, 10, 559. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Egea, I.; Barsan, C.; Bian, W.; Purgatto, E.; Latché, A.; Chervin, C.; Bouzayen, M.; Pech, J.C. Chromoplast differentiation: Current status and perspectives. Plant Cell Physiol. 2010, 51, 1601–1611. [Google Scholar] [CrossRef] [Green Version]
- Alba, R.; Payton, P.; Fei, Z.; McQuinn, R.; Debbie, P.; Martin, G.B.; Tanksley, S.D.; Giovannoni, J.J. Transcriptome and selected metabolite analyses reveal multiple points of ethylene control during tomato fruit development. Plant Cell 2005, 17, 2954–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, C.; Vrebalov, J.; Tafelmeyer, P.; Giovannoni, J.J. The tomato MADS-box transcription factor RIPENING INHIBITOR interacts with promoters involved in numerous ripening processes in a COLORLESS NONRIPENING-dependent manner. Plant Physiol. 2011, 157, 1568–1579. [Google Scholar] [CrossRef] [Green Version]
- Adato, A.; Mandel, T.; Mintz-Oron, S.; Venger, I.; Levy, D.; Yativ, M.; Domínguez, E.; Wang, Z.; De Vos, R.C.H.; Jetter, R.; et al. Fruit-surface flavonoid accumulation in tomato is controlled by a SlMYB12-regulated transcriptional network. PLoS Genet. 2009, 5, e1000777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razifard, H.; Ramos, A.; Della Valle, A.L.; Bodary, C.; Goetz, E.; Manser, E.J.; Li, X.; Zhang, L.; Visa, S.; Tieman, D.; et al. Genomic Evidence for Complex Domestication History of the Cultivated Tomato in Latin America. Mol. Biol. Evol. 2020, 37, 1118–1132. [Google Scholar] [CrossRef] [PubMed]
- Welty, N. The Early History of Tomato Fruit Morphology. Available online: https://vanderknaaplab.uga.edu/documents/The-early-history-of-tomato-fruit-morphology-characteristics.pdf (accessed on 23 July 2021).
- Peralta, I.E.; Spooner, D.M. Granule-bound starch synthase (GBSSI) gene phylogeny of wild tomatoes (Solanum L. section Lycopersicon [Mill.] Wettst. Subsection Lycopersicon). Am. J. Bot. 2001, 88, 1888–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 100 Tomato Genome Sequencing Consortium. Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J. 2014, 80, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, F.; Wu, F.; Ané, C.; Tanksley, S.; Spooner, D.M. Do potatoes and tomatoes have a single evolutionary history, and what proportion of the genome supports this history? BMC Evol. Biol. 2009, 9, 191. [Google Scholar] [CrossRef] [Green Version]
- Bedinger, P.A.; Chetelat, R.T.; McClure, B.; Moyle, L.C.; Rose, J.K.C.; Stack, S.M.; van der Knaap, E.; Baek, Y.S.; Lopez-Casado, G.; Covey, P.A.; et al. Interspecific reproductive barriers in the tomato clade: Opportunities to decipher mechanisms of reproductive isolation. Sex. Plant Reprod. 2011, 24, 171–187. [Google Scholar] [CrossRef]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to beta-carotene formation in plant chromoplasts discovered by map-based cloning of beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.J.; Park, W.J.; Lee, G.-M.; Oh, C.-S.; Yeam, I.; Won, D.-C.; Kim, C.K.; Lee, J.M. Inferring the Genetic Determinants of Fruit Colors in Tomato by Carotenoid Profiling. Molecules 2017, 22, 764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of beta-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Tanambell, H.; Bishop, K.S.; Quek, S.Y. Tangerine tomatoes: Origin, biochemistry, potential health benefits and future prospects. Crit. Rev. Food Sci. Nutr. 2020, 12, 1–12. [Google Scholar]
- Fray, R.G.; Grierson, D. Identification and genetic analysis of normal and mutant phytoene synthase genes of tomato by sequencing, complementation and co-suppression. Plant Mol. Biol. 1993, 22, 589–602. [Google Scholar] [CrossRef]
- Barry, C.S.; McQuinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-flesh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol. 2008, 147, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.S.; Pandey, P. A survey of cultivated heirloom tomato varieties identifies four new mutant alleles at the green-flesh locus. Mol. Breed. 2009, 24, 269–276. [Google Scholar] [CrossRef]
- Ballester, A.R.; Molthoff, J.; de Vos, R.; Hekkert, B.T.; Orzaez, D.; Fernández-Moreno, J.P.; Tripodi, P.; Grandillo, S.; Martin, C.; Heldens, J.; et al. Biochemical and molecular analysis of pink tomatoes: Deregulated expression of the gene encoding transcription factor SlMYB12 leads to pink tomato fruit color. Plant Physiol. 2010, 152, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-I.; Hwang, I.; Goswami, G.; Jung, H.-J.; Nath, U.K.; Yoo, H.-J.; Lee, J.M.; Nou, I.S. Molecular Insights Reveal Psy1, SGR, and SlMYB12 Genes are Associated with Diverse Fruit Color Pigments in Tomato (Solanum lycopersicum L.). Molecules 2017, 22, 2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzali, S.; Mazzucato, A.; Perata, P. Purple as a tomato: Towards high anthocyanin tomatoes. Trends Plant Sci. 2009, 14, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.M.; Mes, P.; Myers, J.R. Characterization and inheritance of the Anthocyanin fruit (Aft) tomato. J. Hered. 2003, 94, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Mes, P.J.; Boches, P.; Myers, J.R. Characterization of tomatoes expressing anthocyanin in the fruit. J. Am. Soc. Hortic. Sci. 2008, 33, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Mustilli, A.C.; Fenzi, F.; Ciliento, R.; Alfano, F.; Bowler, C. Phenotype of the tomato high pigment-2 mutant is caused by a mutation in the tomato homolog of DEETIOLATED1. Plant Cell 1999, 11, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, M.; Segev, O.; Gilboa, N.; Lalazar, A.; Levin, I. The tomato homolog of the gene encoding UV-damaged DNA binding protein 1 (DDB1) underlined as the gene that causes the high pigment-1 mutant phenotype. Theor. Appl. Genet. 2004, 108, 1574–1581. [Google Scholar] [CrossRef]
- Vrebalov, J.; Ruezinsky, D.; Padmanabhan, V.; White, R.; Medrano, D.; Drake, R.; Schuch, W.; Giovannoni, J. A MADS-box gene necessary for fruit ripening at the tomato Ripening-Inhibitor (Rin) locus. Science 2002, 296, 343–346. [Google Scholar] [CrossRef]
- Manning, K.; Tör, M.; Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Giovannoni, J.J.; Seymour, G.B. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat. Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A new tomato NAC (NAM/ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2014, 55, 119–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanahan, M.B.; Yen, H.C.; Giovannoni, J.J.; Klee, H.J. The never ripe mutation blocks ethylene perception in tomato. Plant Cell 1994, 6, 521–530. [Google Scholar]
- Zhu, H.L.; Zhu, B.Z.; Shao, Y.; Wang, X.G.; Lin, X.J.; Xie, Y.H.; Li, Y.C.; Gao, H.Y.; Luo, Y.B. Tomato fruit development and ripening are altered by the silencing of LeEIN2 gene. J. Integr. Plant Biol. 2006, 48, 1478–1485. [Google Scholar] [CrossRef]
- Hu, Z.; Deng, L.; Chen, X.; Wang, P.; Chen, G. Co-suppression of the EIN2-homology gene LeEIN2 inhibits fruit ripening and reduces ethylene sensitivity in tomato. Russ. J. Plant Physiol. 2010, 57, 554–559. [Google Scholar] [CrossRef]
- Zhao, W.; Gao, L.; Li, Y.; Wang, M.; Zhang, L.; Zhao, L. Yellow-fruited phenotype is caused by 573 bp insertion at 5’ UTR of YFT1 allele in yft1 mutant tomato. Plant Sci. 2020, 300, 110637. [Google Scholar] [CrossRef]
- Ranc, N.; Muños, S.; Santoni, S.; Causse, M. A clarified position for Solanum lycopersicum var. cerasiforme in the evolutionary history of tomatoes (solanaceae). BMC Plant Biol. 2008, 8, 130. [Google Scholar]
- Jenkins, J.A.; Mackinney, G. Inheritance of Carotenoid Differences in the Tomato Hybrid Yellow x Tangerine. Genetics 1953, 38, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, L.; Jiao, C.; Stravoravdis, S.; Hosmani, P.S.; Saha, S.; Zhang, J.; Mainiero, S.; Strickler, S.R.; Catala, C.; et al. Genome of Solanum pimpinellifolium provides insights into structural variants during tomato breeding. Nat. Commun. 2020, 11, 5817. [Google Scholar] [CrossRef] [PubMed]
- Efremov, G.I.; Slugina, M.A.; Shchennikova, A.V.; Kochieva, E.Z. Differential Regulation of Phytoene Synthase PSY1 During Fruit Carotenogenesis in Cultivated and Wild Tomato Species (Solanum section Lycopersicon). Plants 2020, 9, 1169. [Google Scholar] [CrossRef]
- Verhoeyen, M.E.; Bovy, A.; Collins, G.; Muir, S.; Robinson, S.; de Vos, C.H.R.; Colliver, S. Increasing antioxidant levels in tomatoes through modification of the flavonoid biosynthetic pathway. J. Exp. Bot. 2002, 53, 2099–2106. [Google Scholar] [CrossRef] [Green Version]
- Blencowe, B.J. Alternative Splicing: New Insights from Global Analyses. Cell 2006, 126, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, N.G.; Xu, J.; Cheng, Y.J.; Deng, X.X. Lycopene-ε-cyclase pre-mRNA is alternatively spliced in Cara Cara navel range (Citrus sinensis Osbeck). Biotechnol. Lett. 2005, 27, 779–782. [Google Scholar] [CrossRef]
- Howitt, C.A.; Cavanagh, C.R.; Bowerman, A.F.; Cazzonelli, C.; Rampling, L.; Mimica, J.L.; Pogson, B.J. Alternative splicing, activation of cryptic exons and amino acid substitutions in carotenoid biosynthetic genes are associated with lutein accumulation in wheat endosperm. Funct. Integr. Genom. 2009, 9, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, D.; Voß, B.; Maass, D.; Wüst, F.; Schaub, P.; Beyer, P.; Welsch, R. Carotenogenesis is regulated by 5ʹUTR-mediated translation of phytoene synthase splice variants. Plant Physiol. 2016, 172, 2314–2326. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Xiao, H. Identification of alternative splicing events by RNA sequencing in early growth tomato fruits. BMC Genom. 2015, 16, 948. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, S.; Chen, M. Characterization and Function of Circular RNAs in Plants. Front. Mol. Biosci. 2020, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Zhou, Z.; Niu, Y.; Sun, X.; Deng, Z. Identification and Functional Characterization of Tomato CircRNAs Derived from Genes Involved in Fruit Pigment Accumulation. Sci. Rep. 2017, 7, 8594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.N.; Rogers, M.F.; Richardson, D.N.; Hamilton, M.; Ben-Hur, A. Deciphering the plant splicing code: Experimental and computational approaches for predicting alternative splicing and splicing regulatory elements. Front. Plant Sci. 2012, 3, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Guo, G.; Hu, X.; Zhang, Y.; Li, Q.; Li, R.; Zhuang, R.; Lu, Z.; He, Z.; Fang, X.; et al. Deep RNA sequencing at single base-pair resolution reveals high complexity of the rice transcriptome. Genome Res. 2010, 20, 646–654. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.; Gu, Q.; Tian, P.; Xiao, L.; Cao, H.; Yanget, W. A chimeric transcript containing Psy1 and a potential mRNA is associated with yellow flesh color in tomato accession PI 114490. Planta 2014, 240, 1011–1021. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, W.; Li, Y.; Feng, X.; Du, K.; Wang, G.; Zhao, L. Identified trans-splicing of YELLOW-FRUITED TOMATO 2 encoding the PHYTOENE SYNTHASE 1 protein alters fruit color by map-based cloning, functional complementation and RACE. Plant Mol. Biol. 2019, 100, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.S.; Lee, J.M. The Fusion Transcript of Phytoene Synthase 1 Controls Yellow Fruit in Tomato. Hort. Sci. Technol. 2020, 38, 705–716. [Google Scholar]
- Xiao, Y.; Kang, B.; Li, M.; Xiao, L.; Xiao, H.; Shen, H.; Yang, W. Transcription of lncRNA AcoS-AS1 is essential to trans-splicing between SlPsy1 and AcoS-AS1 that causes yellow fruit in tomato. RNA Biol. 2020, 17, 596–607. [Google Scholar] [CrossRef]
- Pikaard, C.S.; Mittelsten Scheid, O. Epigenetic regulation in plants. Cold Spring Harb. Perspect. Biol. 2014, 6, a019315. [Google Scholar] [CrossRef]
- Rick, C.M.; Butler, L. Cytogenetics of the Tomato. In Advances in Genetics; Demerec, M., Ed.; Elsevier: Amsterdam, The Netherlands, 1956; Volume 8, pp. 267–382. [Google Scholar]
- Rick, C.M.; Yoder, J.I. Classical and molecular genetics of tomato: Highlights and perspectives. Annu. Rev. Genet. 1988, 22, 281–300. [Google Scholar] [CrossRef]
- Liu, G.; Li, C.; Yu, H.; Tao, P.; Yuan, L.; Ye, J.; Chen, W.; Wang, Y.; Ge, P.; Zhang, J.; et al. GREEN STRIPE, encoding methylated TOMATO AGAMOUS-LIKE 1, regulates chloroplast development and Chl synthesis in fruit. New Phytol. 2020, 228, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Itkin, M.; Seybold, H.; Breitel, D.; Rogachev, I.; Meir, S.; Aharoni, A. TOMATO AGAMOUS-LIKE 1 is a component of the fruit ripening regulatory network. Plant J. 2009, 60, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.W.; Deng, S.; Xu, H.; Mao, H.Z.; Liu, J.; Niu, Q.W.; Wang, H.; Chua, N.H. A noncoding RNA transcribed from the AGAMOUS (AG) second intron binds to CURLY LEAF and represses AG expression in leaves. New Phytol. 2018, 219, 1480–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, J.; Nguyen, C.; Ampofo, B.; Zhong, S.; Fei, Z. The Epigenome and Transcriptional Dynamics of Fruit Ripening. Annu. Rev. Plant Biol. 2017, 68, 61–84. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, X.; Xu, Q.; Zhao, S.; Tai, Y.; Wei, C. Global dissection of alternative splicing uncovers transcriptional diversity in tissues and associates with the flavonoid pathway in tea plant (Camellia sinensis). BMC Plant Biol. 2018, 18, 266. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Liu, Y.; Yin, S.; Qiu, J.; Jin, Q.; King, G.J.; Wang, J.; Ge, X.; Li, Z. Alternatively Spliced BnaPAP2.A7 Isoforms Play Opposing Roles in Anthocyanin Biosynthesis of Brassica napus L. Front. Plant Sci. 2020, 11, 983. [Google Scholar] [CrossRef]
- Lim, S.-H.; Kim, D.-H.; Jung, J.-A.; Lee, J.-Y. Alternative Splicing of the Basic Helix-Loop-Helix Transcription Factor Gene CmbHLH2 Affects Anthocyanin Biosynthesis in Ray Florets of Chrysanthemum (Chrysanthemum morifolium). Front. Plant Sci. 2021, 12, 669315. [Google Scholar] [CrossRef]
- Povero, G.; Gonzali, S.; Bassolino, L.; Mazzucato, A.; Perata, P. Transcriptional analysis in high-anthocyanin tomatoes reveals synergistic effect of Aft and atv genes. J. Plant Physiol. 2011, 168, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Qiu, Z.; Wang, X.; Van Giang, T.; Liu, X.; Wang, J.; Wang, X.; Gao, J.; Guo, Y.; Du, Y.; et al. A putative R3 MYB repressor is the candidate gene underlying atroviolacium, a locus for anthocyanin pigmentation in tomato fruit. J. Exp. Bot. 2017, 68, 5745–5758. [Google Scholar] [CrossRef]
- Colanero, S.; Perata, P.; Gonzali, S. The atroviolacea Gene Encodes an R3-MYB Protein Repressing Anthocyanin Synthesis in Tomato Plants. Front. Plant Sci. 2018, 9, 830. [Google Scholar] [CrossRef] [Green Version]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; Ric de Vos, C.H.; vanTunen, A.J.; Verhoeyen, M.E. Overexpression of Petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Deng, L.; Du, M.; Zhao, J.; Chen, Q.; Huang, T.; Jiang, H.; Li, C.-B.; Li, C. A transcriptional network promotes anthocyanin biosynthesis in tomato flesh. Mol. Plant 2020, 13, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Colanero, S.; Tagliani, A.; Perata, P.; Gonzali, S. Alternative splicing in the Anthocyanin fruit gene encoding an R2R3 MYB transcription factor affects anthocyanin biosynthesis in tomato fruits. Plant Commun. 2020, 1, 100006. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Chen, N.; Huang, Z.; Li, D.; Zhi, J.; Yu, B.; Liu, X.; Cao, B.; Qiu, Z. Anthocyanin Fruit encodes an R2R3-MYB transcription factor, SlAN2-like, activating the transcription of SlMYBATV to fine-tune anthocyanin content in tomato fruit. New Phytol. 2020, 225, 2048–2063. [Google Scholar] [CrossRef]
- Colanero, S.; Perata, P.; Gonzali, S. What’s behind Purple Tomatoes? Insight into the Mechanisms of Anthocyanin Synthesis in Tomato Fruits. Plant Physiol. 2020, 182, 1841–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiferle, C.; Fantini, E.; Bassolino, L.; Povero, G.; Spelt, C.; Buti, S.; Giuliano, G.; Quattrocchio, F.; Koes, R.; Perata, P.; et al. Tomato R2R3-MYB Proteins SlANT1 and SlAN2: Same Protein Activity, Different Roles. PLoS ONE 2015, 10, e0136365. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(A) The carotenoid biosynthetic pathway in tomato fruit. GGPP, geranylgeranyl diphosphate; PSY1, phytoene synthase1; PDS, phytoene desaturase; Z-ISO, ζ-carotene isomerase; ZDS, ζ-carotene desaturase; CRTISO, carotene isomerase; LCY-B, lycopene β-cyclase; LCY-E, lycopene ε-cyclase; CHY-B, β-carotene hydroxylase; CHY-E, ε-carotene hydroxylase; VDE, violaxanthin deepoxidase; ZEP, zeaxanthin epoxidase; NXS, neoxanthin synthase. The enzymatic steps inhibited (red lines) or induced (blue arrows) in the tomato mutants r (yellow flesh), t (tangerine), Del (Delta), B (Beta), and og (old-gold) are indicated. (B) The flavonoid biosynthetic pathway in tomato fruit. PAL, phenylalanine ammonia lyase; C4H, cinnamic acid 4-hydroxylase; 4CL, 4-coumaric acid: CoA ligase; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavonone 3-hydroxylase; F3’H, flavonoid 3’-hydroxylase; F3’5’H, flavonoid 3’5’-hydroxylase; DFR, dihydroflavonol reductase; FLS, flavonol synthase; 3GT, flavonoid-3-O-glucosyltransferase; RT, flavonoid 3-O-glucoside-rhamnosyltransferase; ANS, anthocyanin synthase; UFGT, flavonoid 3-O-glucosyltransferase. y is a genetic locus associated with the mutation in the master regulatory factor SlMYB12. Some of the steps of the flavonoid pathway downregulated in the y mutant are represented. (C) “all-trans-lycopene” molecular structure. (D) “naringenin chalcone” molecular structure.
Figure 1.
(A) The carotenoid biosynthetic pathway in tomato fruit. GGPP, geranylgeranyl diphosphate; PSY1, phytoene synthase1; PDS, phytoene desaturase; Z-ISO, ζ-carotene isomerase; ZDS, ζ-carotene desaturase; CRTISO, carotene isomerase; LCY-B, lycopene β-cyclase; LCY-E, lycopene ε-cyclase; CHY-B, β-carotene hydroxylase; CHY-E, ε-carotene hydroxylase; VDE, violaxanthin deepoxidase; ZEP, zeaxanthin epoxidase; NXS, neoxanthin synthase. The enzymatic steps inhibited (red lines) or induced (blue arrows) in the tomato mutants r (yellow flesh), t (tangerine), Del (Delta), B (Beta), and og (old-gold) are indicated. (B) The flavonoid biosynthetic pathway in tomato fruit. PAL, phenylalanine ammonia lyase; C4H, cinnamic acid 4-hydroxylase; 4CL, 4-coumaric acid: CoA ligase; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavonone 3-hydroxylase; F3’H, flavonoid 3’-hydroxylase; F3’5’H, flavonoid 3’5’-hydroxylase; DFR, dihydroflavonol reductase; FLS, flavonol synthase; 3GT, flavonoid-3-O-glucosyltransferase; RT, flavonoid 3-O-glucoside-rhamnosyltransferase; ANS, anthocyanin synthase; UFGT, flavonoid 3-O-glucosyltransferase. y is a genetic locus associated with the mutation in the master regulatory factor SlMYB12. Some of the steps of the flavonoid pathway downregulated in the y mutant are represented. (C) “all-trans-lycopene” molecular structure. (D) “naringenin chalcone” molecular structure.

Figure 3.
(A) Photo of Solanum chilense anthocyanin-spotted green ripe fruits from plants spontaneously grown in Chile, Toconao, Quebrada de Jere (−23.14553°, −067.98998°, 2703 m, 12 dec 2012). (B) SLL fruits, cv. MicroTom, showing different levels of anthocyanin pigmentation at the mature green stage of ripening. From left to right: Aft/Aft x ATV/atv, Aft/Aft x atv/atv, and Aft/Aft x atv/atv x hp2/hp2 genotypes. (C) SLL fruits, cv. Ailsa Craig, at mature green with Aft/Aft x atv/atv genotype, and complete or partial anthocyanin pigmentations.
Figure 3.
(A) Photo of Solanum chilense anthocyanin-spotted green ripe fruits from plants spontaneously grown in Chile, Toconao, Quebrada de Jere (−23.14553°, −067.98998°, 2703 m, 12 dec 2012). (B) SLL fruits, cv. MicroTom, showing different levels of anthocyanin pigmentation at the mature green stage of ripening. From left to right: Aft/Aft x ATV/atv, Aft/Aft x atv/atv, and Aft/Aft x atv/atv x hp2/hp2 genotypes. (C) SLL fruits, cv. Ailsa Craig, at mature green with Aft/Aft x atv/atv genotype, and complete or partial anthocyanin pigmentations.

Figure 4.
(A) PSY1 gene alternative RNA splicing mechanisms leading to different splicing variants and Circular RNAs (CircRNAs). The alternative mRNAs sequences correspond to the PSY1 transcript variants 1 ÷ 4 in “NCBI Solanum lycopersicon annotation release 103” (in: https://www.ncbi.nlm.nih.gov/gene/543988, 14 July 2021). The number and the colour of the exons in the CircRNAs are only indicative and do not reflect the real sequences. (B) PSY1 trans-spliced fusion transcripts with the adjacent CoA-ligase gene. Gene structures and intergenic distance are not in one-to-one scale.
Figure 4.
(A) PSY1 gene alternative RNA splicing mechanisms leading to different splicing variants and Circular RNAs (CircRNAs). The alternative mRNAs sequences correspond to the PSY1 transcript variants 1 ÷ 4 in “NCBI Solanum lycopersicon annotation release 103” (in: https://www.ncbi.nlm.nih.gov/gene/543988, 14 July 2021). The number and the colour of the exons in the CircRNAs are only indicative and do not reflect the real sequences. (B) PSY1 trans-spliced fusion transcripts with the adjacent CoA-ligase gene. Gene structures and intergenic distance are not in one-to-one scale.

Figure 5.
(A) Exon-intron structure of SlMYBV-ATV gene with possible alternative splicing variants. The relative three different transcripts which can be spliced from the gene (SlMYB-ATV-X1, SlMYB-ATV-X2, and SlMYB-ATV-X3) are reported with the relative start and stop codons. The first three exons, represented in black, are the bioinformatically predicted ones for the gene. The green triangle indicates the position of the 4-nt insertion identified in the gene in the atv mutant genotype. (B) Exon-intron structure of AN2like gene with the two possible 5′ splicing variants. The transcript produced from the Solanum chilense allele (ScAN2like) identified in the Aft tomato accession is reported as well as the two differently spliced transcripts produced in Solanum lycopersicum (SlAN2like-1 and SlAN2like-2), which both show premature stop codons.
Figure 5.
(A) Exon-intron structure of SlMYBV-ATV gene with possible alternative splicing variants. The relative three different transcripts which can be spliced from the gene (SlMYB-ATV-X1, SlMYB-ATV-X2, and SlMYB-ATV-X3) are reported with the relative start and stop codons. The first three exons, represented in black, are the bioinformatically predicted ones for the gene. The green triangle indicates the position of the 4-nt insertion identified in the gene in the atv mutant genotype. (B) Exon-intron structure of AN2like gene with the two possible 5′ splicing variants. The transcript produced from the Solanum chilense allele (ScAN2like) identified in the Aft tomato accession is reported as well as the two differently spliced transcripts produced in Solanum lycopersicum (SlAN2like-1 and SlAN2like-2), which both show premature stop codons.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gonzali, S.; Perata, P. Fruit Colour and Novel Mechanisms of Genetic Regulation of Pigment Production in Tomato Fruits. Horticulturae 2021, 7, 259. https://doi.org/10.3390/horticulturae7080259
AMA Style
Gonzali S, Perata P. Fruit Colour and Novel Mechanisms of Genetic Regulation of Pigment Production in Tomato Fruits. Horticulturae. 2021; 7(8):259. https://doi.org/10.3390/horticulturae7080259
Chicago/Turabian StyleGonzali, Silvia, and Pierdomenico Perata. 2021. "Fruit Colour and Novel Mechanisms of Genetic Regulation of Pigment Production in Tomato Fruits" Horticulturae 7, no. 8: 259. https://doi.org/10.3390/horticulturae7080259
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.