
Temperate Fruit Trees under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Climate Change and Temperate Fruit Productivity
3. Understanding the Role Dormancy
3.1. Dormancy Breaking Chemical Compounds
3.2. Dormancy Breaking Organic Compounds

{kind=link}
{kind=link}
| Compound | Application Time | Applied Concentrations (Effective Concentrations) | Crop (Scientific Name) | Ref. |
|---|---|---|---|---|
| Potassium nitrate (KNO3) | During germination | 0.2, 1, and 3% (0.2%) | Agrimony (Agrimonia eupatoria L.) | [76] |
| Gibberellic acid (GA3) | During germination | 100, 200, and 300 mg L−1 (100 mg L−1) | Agrimony (Agrimonia eupatoria L.) | [76] |
| Hydrogen cyanamide (HC) | During bud cutting | 5% for 4-year-old cuttings | Grapes (Vitis vinifera L. × Vitis labruscana Bailey) | [77] |
| GA3 or KNO3 | During pre-germination | Seeds soaking in GA3 (400 ppm) for 24 h or KNO3 (1 M) for 30 min | Highland papaya (Vasconcelle aquercifolia L.) | [78] |
| GA3 | During seed priming | Seeds primed in GA3 (750 ppm) for 48h | Common poppy (Papaver rhoeas L.) and (P. dubium L.) | [79] |
| KNO3 | During seed priming | Seed treatment with KNO3 (0.5 g L−1) for 24 h | ||
| GA3 or KNO3 | During germination | Dormant seeds treated with GA3 (150 ppm) or KNO3 (1000 ppm) for 24 h. | Wall rocket (Diplot axiserucoides L.) | [80] |
| Garlic extract (GE) or HC | Before winter dormancy | Cuttings immersed in GE (10%) or HC (4%) (v/v) for 10 s | Grape (Vitis vinifera L.) | [68] |
| HC (Dormex) | During bud cutting | 6-year-old cuttings treated with HC (5%) | Grape (Vitis vinifera L. × Vitis labruscana Bailey) | [13,64] |
| GA3 | During germination | Seed soaking in GA3 (10, 100, or 1000 mg L−1) and scoring at 30 d | Ashitaba Angelica keiskei | [81] |
| Zinc sulphate (ZnSO4) | Before winter dormancy | Foliar application (1000, 1500, and 2000 mg L−1) (2000 mg/L) | Kiwifruit (Actinidia deliciosa L.) | [82] |
| HC | 45 days before natural bud break | 2, 4 and 6% (4%) + mineral oil (2%) | Kiwifruit (Actinidia deliciosa L.), cv. Hayward | [83] |
| Erger Biostimulant | At week no. 5, 9, and 13 from bud break | Foliar application (6%) once at three different times in fall | Kiwifruit (Actinidia deliciosa L.) | [84] |
| KNO3 or GA3 | At stratification of seeds | Seed soaking in KNO3 (0.2%) or GA3 (5 mM) | Apple (Malus domestica L.), cv. Ligol | [85] |
| HC | In late August | Foliar application (12.5 mM) | Grapevines (V. vinifera L. x V. labruscana Bailey), cv. Kyoho | [86] |
| HC, KNO3, mineral oil, thiourea, or Ca(NO3)2 | In December | Foliar application of HC (4%), KNO3 (8%), mineral oil (6%), thiourea (2%), or Ca(NO3)2 (6%) | Apple (Malus sylvestris Mill), cv. Anna | [64] |
3.3. Some Aspects of the Mechanism of Dormancy Breaking
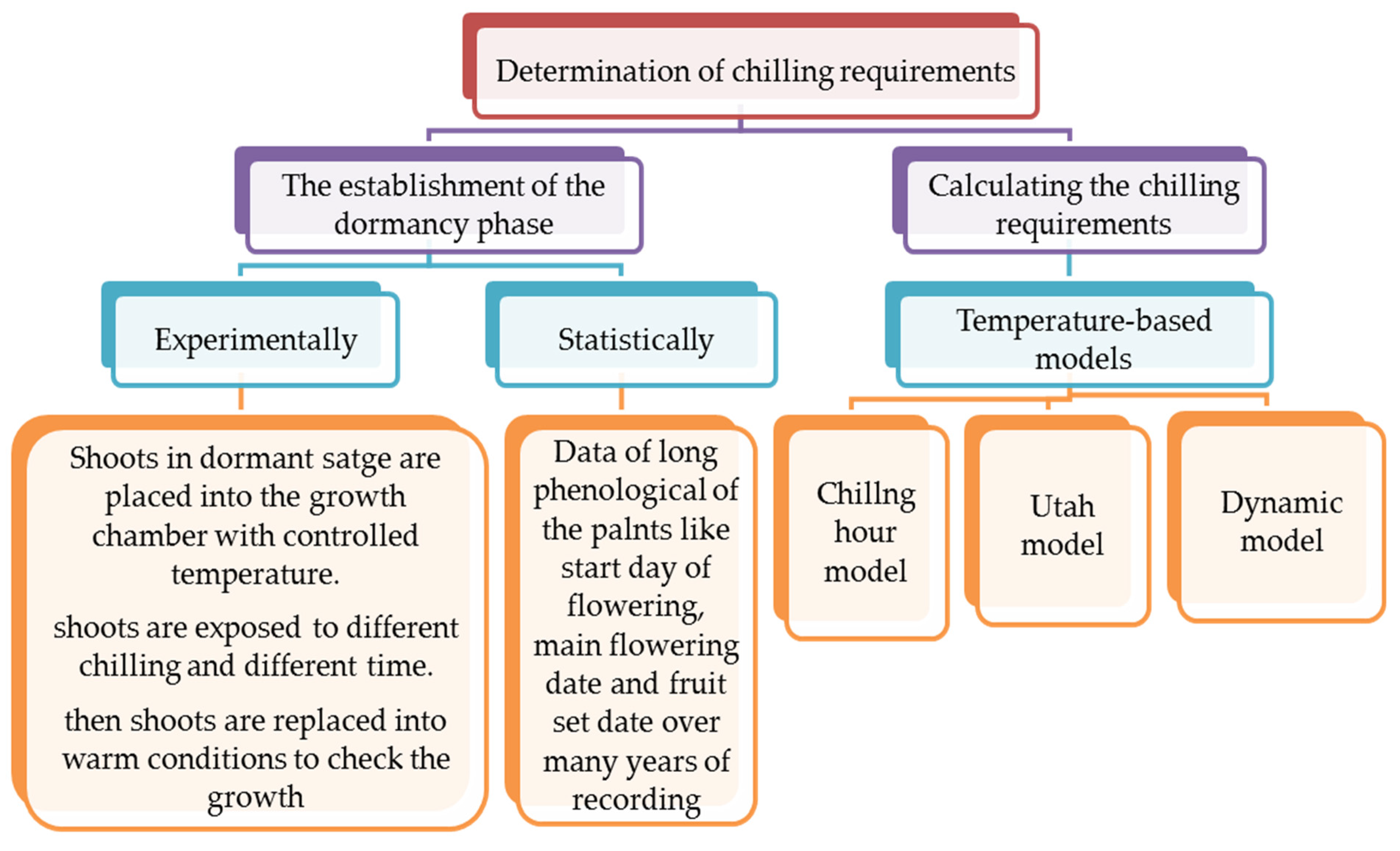
4. Chilling Requirements
4.1. Chilling Requirements Versus Climate Change and Warm Winter Conditions
4.2. Models to Calculate Chilling Requirements
| Model Name | Description | Ref. |
|---|---|---|
| Chilling hours | Simple and widely used. Counts the number of hours below 7.2 °C. | [127] |
| Utah | Uses temperatures that trees are exposed to, expressed as chilling units (CU): 1 CU for 2.5–9.1 °C, 0.5 CU for 9.2–12.4 °C, 0 CU for <1.4 or 12.5–15.9 °C, −0.5 CU for 16–18 °C, and -1 CU for >18 °C. | [128] |
| Landsberg | Uses CU based on dividing the daily temperature by a base temperature of the crop. If the base temperature is 5 °C and the daily temperature is 5 °C, then the chilling unit is 1. | [131] |
| Low Chill | Developed for the low chilling requirements of the ‘Sungold’ nectarine, it can be used for low-chilling-requirement cultivars in subtropics. Uses CU of 1 for 1.8–8 °C, 0 for <−1 and 14–17 °C, and −1 for >19.5 °C. | [132] |
| North Carolina | Uses CU of 1 for 1.6–7.2 °C, 0 for <−1.1 °C and 16.5–19 °C, and −2 for >23.3 °C. | [133] |
| Dynamic | Proposes an intermediate product formed in buds due to cold temperature that can be reversed by warm temperature. If this product reaches a certain concentration at cold temperature, the chill portions are permanently fixed, and they are considered unaffected by warm temperature. One chill portion (CP) is an exposure to 6 °C for 28 hours. | [134] |
| Positive Chill Units (PCU) | A modified version of the Utah model to avoid negative values at warm temperature. CU = 0 is used for >15.9 °C. | [135] |
| ‘Sweetheart’ cherry and ‘Gala’ apple | Uses CU = 1 for −2.1–7 °C (cherry) and −2.1–5.5 °C (apple), −1 for >18 °C (cherry and apple), and 0 for 13–16 °C (cherry and apple). | [136] |
| Jones I | Was used to study the chilling requirements of 20 blackcurrant genotypes. The model depends on a decreasing exponential curve by fitting a curve with the response to chilling temperature. The decreasing exponential = 1 CU for −2.7 °C, 0.5 CU for 2.0 °C, 0.25 CU for 6.6 °C, and 0.1 CU for 12.8 °C. | [137] |
| Jones II | Winter chill data of 0–7.2 °C (denoted by 0–7.2 CU) calculated by the summation of total chill hours from October 1st in a relevant year when the mean temperature was 0–7.2 °C. Similarly, <7.2 °C (denoted by 7.2 CU) for mean temperature <7.2 °C, and exponential units (denoted by “exp”), where exp is the summation of hours from 1 October for the function exp = 0.6702 [exp (−0.148 × Ta)], where Ta is the hourly measured mean temperature. | [138] |
| Luedeling: ’chillR’ | “chillR” contains functions for processing temperature records into chilling models (chilling hours, Utah, and dynamic models) and heat units (growing degree hours). | [139] |
5. Conclusions
- New and more effective dormancy breaking organic compounds have to be discovered;
- New climate-smart cultivars have to be bred in order to solve the increasing difficulties for dormancy and chilling requirements [146], i.e., a longer period from the fulfillment of chilling requirements to bud break. This would help to escape spring cold events in some years in Mediterranean areas;
- Dormancy and chilling forecasting models have to be improved further in order to successfully manage temperate fruit tree production worldwide but especially in the tropical and subtropical regions.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Boadu, F.O. Climate change. In Agricultural Law and Economics in Sub-Saharan Africa Cases and Comments; Boadu, F.O., Ed.; Elsevier Inc.: Dordrecht, The Netherlands, 2016; pp. 555–571. [Google Scholar] [CrossRef]
- Miranda, C.; Urrestarazu, J.; Santesteban, L.G. Fruclimadapt: An R package for climate adaptation assessment of temperate fruit species. Comp. Electr. Agric. 2021, 180, 105879. [Google Scholar] [CrossRef]
- Ahmed, N.; Lone, F.A.; Hussain, K.; Kanth, R.H.; Mahdi, S.S. Impact of Climate change on temperate fruit production in Kashmir Valley, North Western Himalayan Region of India—Challenges, opportunities and way forward. In Climate Change and Agriculture in India: Impact and Adaptation; Sheraz Mahdi, S., Ed.; Springer International Publishing AG: Cham, Switzerland, 2019; pp. 251–262. [Google Scholar] [CrossRef]
- Anon. 2020. Earth’s CO2 Home Page. Available online: https://www.co2.earth (accessed on 25 December 2020).
- Pio, R.; de Souza, F.B.M.; Kalcsits, L.; Bisi, R.B.; Farias, D.H. Advances in the production of temperate fruits in the tropics. Acta Sci. 2019, 41, e39549. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.; Pérez-López, D.; Centeno, A.; Ruiz-Ramos, M. Viability of temperate fruit tree varieties in Spain under climate change according to chilling accumulation. Agric. Syst. 2021, 186, 102961. [Google Scholar] [CrossRef]
- Yahia, E.M. Classification of horticultural commodities. In Postharvest Technology of Perishable Horticultural Commodities; Yahia, E.M., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2019; pp. 71–97. [Google Scholar] [CrossRef]
- FAO Faostat: Statistical Database. Food and Agriculture Organization of the United Nation, Rome, Italy. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 2 May 2019).
- Rai, R.; Joshi, S.; Roy, S.; Singh, O.; Samir, M.; Chandra, A. Implications of changing climate on productivity of temperate fruit crops with special reference to apple. J. Hortic. 2015, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E. A conceptual framework for winter dormancy in deciduous trees. Agronomy 2020, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Khalil-Ur-Rehman, M.; Wang, W.; Dong, Y.; Faheem, M.; Xu, Y.; Gao, Z.; Tao, J. Comparative transcriptomic and proteomic analysis to deeply investigate the role of hydrogen cyanamide in grape bud dormancy. Int. J. Mol. Sci. 2019, 20, 3528. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.A.; Díaz-Vivancos, P.; Martínez-Sanchez, G.; Alburquerque, N.; Martínez, D.; Barba-Espín, G.; Acosta-Motos, J.R.; Carrera, E.; García-Brunton, J. Physiological and biochemical characterization of bud dormancy: Evolution of carbohydrate and antioxidant metabolisms and hormonal profile in a low chill peach variety. Sci. Hortic. 2021, 281, 109957. [Google Scholar] [CrossRef]
- Vegis, A. Dormancy in higher plants. Annu. Rev. Plant Physiol. 1964, 15, 185–224. [Google Scholar] [CrossRef]
- Chuine, I.; Régnière, J. Process-based models of phenology for plants and animals. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 159–182. [Google Scholar] [CrossRef]
- Zhao, H.; Fu, Y.H.; Wang, X.; Zhang, Y.; Liu, Y.; Janssens, I.A. Diverging models introduce large uncertainty in future climate warming impact on spring phenology of temperate deciduous trees. Sci. Total Environ. 2021, 757, 143903. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endodormancy, paradormancy, and ecodormancy—Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar]
- Bielenberg, D.G.; Wang, Y.E.; Li, Z.; Zhebentyayeva, T.; Fan, S.; Reighard, G.L.; Scorza, R.; Abbott, A.G. Sequencing and annotation of the evergrowing locus in peach [Prunus persica (L.) Batsch] reveals a cluster of six MADS-box transcription factors as candidate genes for regulation of terminal bud formation. Tree Genet. Genom. 2008, 4, 495–507. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome profiles reveal the crucial roles of hormone and sugar in the bud dormancy of Prunus mume. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fenili, C.L.; Petri, J.L.; Sezerino, A.A.; De Martin, M.S.; Gabardo, G.C.; Daniel, E.S. Bluprins® as alternative bud break promoter for ‘Maxi Gala’ and ‘Fuji Suprema’ apple trees. J. Exp. Agric. Internat. 2018, 26, 1–13. [Google Scholar] [CrossRef]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud dormancy in perennial fruit tree species: A pivotal role for oxidative cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- AR4 Climate Change. The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 1–450. [Google Scholar]
- Luedeling, E.; Girvetz, E.H.; Brown, P.H. Climate change affects winter chill for temperate fruit and nu tress. PLoS ONE 2011, 6, e20155. [Google Scholar] [CrossRef]
- Guédon, Y.; Legave, J.M. Analyzing the time-course variation of apple and pear tree dates of flowering stages in the global warming context. Ecol. Model. 2008, 219, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Costa, C.; Stassen, P.J.C.; Mudzunga, J. Chemical rest breaking agents for the South African pome and stone fruit industry. Acta Hortic. 2004, 636, 295–302. [Google Scholar] [CrossRef]
- Erez, A. Means to compensate for insufficient chilling to improve bloom and leafing. Acta Hortic. 1995, 395, 81–95. [Google Scholar] [CrossRef]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and heat requirements of temperate stone fruit trees (Prunus sp.). Agronomy 2020, 10, 409. [Google Scholar] [CrossRef] [Green Version]
- El Yaacoubi, A.; El Jaouhari, N.; Bourioug, M.; El Youssfi, L.; Cherroud, S. Potential vulnerability of Moroccan apple orchard to climate change – induced phenological perturbations: Effects on yields and fruit quality. Int. J. Biometeorol. 2020, 64, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, P.; Bhaskar, R.; Papnai, G.; Joshi, N.; Supyal, V. Impact of climate change on apple phenology and adaptability of Anna variety (low chilling cultivar) in lower hills of Uttarakhand. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 453–460. [Google Scholar] [CrossRef]
- Díez-Palet, I.; Funes, I.; Savé, R.; Biel, C.; Herralde, F.; Miarnau, X.; Vargas, F.; Àvila, G.; Carbó, J.; Aranda, X. Blooming under Mediterranean climate: Estimating cultivar-specific chill and heat requirements of almond and apple trees using a statistical approach. Agronomy 2019, 9, 760. [Google Scholar] [CrossRef] [Green Version]
- Pfleiderer, P.; Menke, I.; Schleussner, C. Increasing risks of apple tree frost damage under climate change. Clim. Chang. 2019, 57, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, H.; Ghalhari, G.F.; Baaghideh, M. Impacts of climate change on apple tree cultivation areas in Iran. Clim. Chang. 2019, 153, 91–103. [Google Scholar] [CrossRef]
- Wolfe, D.W.; De Gaetano, A.T.; Peck, G.M.; Carey, M.; Ziska, L.H.; Lea-Cox, J.; Kemanian, A.R.; Hoffmann, M.P.; Hollinger, D.Y. Unique challenges and opportunities for northeastern US crop production in a changing climate. Clim. Chang. 2018, 146, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Funes, I.; Aranda, X.; Biel, C.; Carbó, J.; Camps, F.; Antonio, J.; Molina de Herralde, F.; Graua, B.; Savé, R. Future climate change impacts on apple flowering date in a Mediterranean subbasin. Agri. Water Manag. 2016, 164, 19–27. [Google Scholar] [CrossRef]
- Chand, H.; Verma, S.C.; Bhardwaj, S.K.; Sharma, S.D.; Mahajan, P.K.; Sharma, R. Effect of changing climatic conditions on chill units accumulation and productivity of apple in mid hill sub humid zone of Western Himalayas, India. Curr. World Environ. 2016, 11, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Antolín, M.C.; Toledo, M.; Pascual, I.; Irigoyen, J.J.; Goicoechea, N. The Exploitation of local Vitis vinifera L. biodiversity as a valuable tool to cope with climate change maintaining berry quality. Plants 2021, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, S.; Fattahi, M.; Bedis, K.; Nasrolahpour-moghadam, S.; Irigoyen, J.J.; Gogorcena, Y. Interactional effects of climate change factors on the water status, photosynthetic rate, and metabolic regulation in peach. Front. Plant Sci. 2020, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Arrizabalaga-Arriazu, M.; Gomès, E.; Morales, F.; Irigoyen, J.J.; Pascual, I.; Hilbert, G. High temperature and elevated carbon dioxide modify berry composition of different clones of grapevine (Vitis vinifera L.) cv. Tempranillo. Front. Plant Sci. 2020, 11, 603687. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vucetic, V.; Dai, Z.; Luedeling, E.; van Leeuwen, C.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of grapevine phenology to water availability, temperature and CO2 concentration. Front. Environ. Sci. 2016, 4, 48. [Google Scholar] [CrossRef]
- Salazar-Parra, C.; Aranjuelo, I.; Pascual, I.; Erice, G.; Sanz-Sáez, Á.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Araus, J.L.; Morales, F. Carbon balance, partitioning and photosynthetic acclimation in fruit-bearing grapevine (Vitis vinifera L. cv. Tempranillo) grown under simulated climate change (elevated CO2, elevated temperature and moderate drought) scenarios in temperature gradient greenhouses. J. Plant Physiol. 2015, 174, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, W.; Zhang, Q.; Lu, X.; Wei, C.; Yu, S.; Zhou, Z. Improvement of flavour quality and consumer acceptance during postharvest ripening in greenhouse peaches by carbon dioxide enrichment. Food Chem. 2014, 164, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-H.; Cho, J.G.; Son, I.-C.; Kim, S.H.; Lee, I.-B.; Choi, I.M.; Kim, D. Effects of elevated carbon dioxide and temperature on photosynthesis and fruit characteristics of ‘Niitaka’ pear (Pyrus pyrifolia Nakai). Hortic. Environ. Biotechnol. 2012, 53, 357–361. [Google Scholar] [CrossRef]
- Chitu, E.; Paltineanu, C. Timing of phenological stages for apple and pear trees under climate change in a temperate-continental climate. Int. J. Biometeorol. 2020, 64, 1263–1271. [Google Scholar] [CrossRef]
- Schattman, R.E.; Niles, M.T.; Aitken, H.M. Water use governance in a temperate region: Implications for agricultural climate change adaptation in the Northeastern United States. Ambio 2020, 50, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Fu, Y.H.; Hao, F.; Zhou, X.; Zhang, X.; Yin, G.; Vitasse, Y.; Piao, S.; Niu, K.; De Boeck, H.J. Climate warming increases spring phenological differences among temperate trees. Global Chang. Biol. 2020, 26, 5979–5987. [Google Scholar] [CrossRef]
- Haokip, S.W.; Shankar, K.; Lalrinngheta, J. Climate change and its impact on fruit crops. J. Pharmacogn. Phytochem. 2020, 9, 435–438. [Google Scholar]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Naulleau, A.; Gary, C.; Prévot, L.; Hossard, L. Evaluating strategies for adaptation to climate change in grapevine production–A systematic review. Front. Plant Sci. 2021, 11, 607859. [Google Scholar] [CrossRef]
- Parker, L.; Pathak, T.; Ostoja, S. Climate change reduces frost exposure for high-value California orchard crops. Sci. Total Environ. 2021, 762, 143971. [Google Scholar] [CrossRef]
- Jha, P.K.; Materia, S.; Zizzi, G.; Costa-Saura, J.M.; Trabucco, A.; Evans, J.; Bregaglio, S. Climate change impacts on phenology and yield of hazelnut in Australia. Agric. Syst. 2021, 186, 102982. [Google Scholar] [CrossRef]
- Vanalli, C.; Casagrandi, R.; Gatto, M.; Bevacqua, D. Shifts in the thermal niche of fruit trees under climate change: The case of peach cultivation in France. Agric. For. Meteorol. 2021, 300, 108327. [Google Scholar] [CrossRef]
- Zahoor, S.; Dutt, V.; Mughal, A.H.; Pala, N.A.; Qaisar, K.N.; Khan, P.A. Apple-based agroforestry systems for biomass production and carbon sequestration: Implication for food security and climate change contemplates in temperate region of Northern Himalaya, India. Agroforest Syst. 2021, 95, 367–382. [Google Scholar] [CrossRef]
- Anderson, J.V. Advances in Plant Dormancy; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–325. [Google Scholar] [CrossRef]
- Tanner, J.D.; Minas, I.S.; Chen, K.Y.; Jenderek, M.M.; Wallner, S.J. Antimicrobial forcing solution improves recovery of cryopreserved temperate fruit tree dormant buds. Cryobiology 2020, 92, 241–247. [Google Scholar] [CrossRef]
- Rojo, J.; Orlandi, F.; Ben Dhiab, A.; Lara, B.; Picornell, A.; Oteros, J.; Msallem, M.; Fornaciari, M.; Pérez-Badia, R. Estimation of chilling and heat accumulation periods based on the timing of olive pollination. Forests 2020, 11, 835. [Google Scholar] [CrossRef]
- Melke, A. The physiology of chilling temperature requirements for dormancy release and bud-break in temperate fruit trees grown at mild winter tropical climate. J. Plant Stud. 2015, 4, 110–156. [Google Scholar] [CrossRef]
- Vitasse, Y.; Schneider, L.; Rixen, C.; Christen, D.; Rebetez, M. Increase in the risk of exposure of forest and fruit trees to spring frosts at higher elevations in Switzerland over the last four decades. Agric. For. Meteorol. 2018, 248, 60–69. [Google Scholar] [CrossRef]
- Oliveira, I.V.M.; Lopes, P.R.C.; Silva Matos, R.R.S.; Cavalcante, I.H.L. Apple phenology, cv. ‘Countess’ in the San Francisco valley. J. Agric. Sci. 2013, 36, 23–30. (In Portuguese) [Google Scholar]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Prudencio, A.S.; Díaz-Vivancos, P.; Dicenta, F.; Hernandez, J.A.; Martínez-Gomez, P. Monitoring the transition from endodormancy to ecodormancy in almond through the analysis and expression of a specific Class III peroxidase gene. Tree Genet. Genomes 2019, 15, 44. [Google Scholar] [CrossRef]
- Singh, H. Use of chemicals in fruit crops for dormancy induction: A Review. Int. J. Ecol. Environ. Sci. 2020, 2, 249–250. [Google Scholar]
- Ionescu, I.A.; Moller, B.L.; Sánchez-Pérez, R. Chemical control of Flowering time. J. Exp. Bot. 2017, 68, 369–382. [Google Scholar] [CrossRef]
- Seif El-Yazal, M.A.S.; El-Yazal, S.A.S.; Rady, M.M. Exogenous dormancy breaking substances positively change endogenous phytohormones and amino acids during dormancy release in ‘Anna’ apple trees. Plant Growth Regul. 2014, 72, 211–220. [Google Scholar] [CrossRef]
- Seif El-Yazal, M.A.S.; Rady, M.M. Trees foliar-applied Dormex™ or thiourea-enhanced proline and biogenic amine contents and hastened breaking bud dormancy in “Ain Shemer” apple. Trees 2013, 27, 161–169. [Google Scholar] [CrossRef]
- Ahmed, F.F.; Ibrahim, H.I.M.; Abada, M.A.M.; Osman, M.M.M. Using plant extracts and chemical rest breakages for breaking bud dormancy and improving productivity of superior grapevines growing under hot climates. World Rural Obs. 2014, 6, 8–18. [Google Scholar]
- Rady, M.M.; El-Yazal, M.A.S. Garlic extract as a novel strategy to hasten dormancy release in buds of ‘Anna’ apple trees. S. Afr. J. Bot. 2014, 92, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Orrantia-Araujo, M.A.; Martínez-Téllez, M.A.; Corrales-Maldonado, C.; Rivera-Domínguez, M.; Vargas-Arispuro, I. Changes in glutathione and glutathione disulfide content in dormant grapevine buds treated with garlic compound mix to break dormancy. Sci. Hortic. 2019, 246, 407–410. [Google Scholar] [CrossRef]
- dos Santos, R.F.; Marques, L.O.D.; Mello-Farias, P.; Martins, C.R.; Konzen, L.H.; Carvalho, I.R.; Malgarim, M.B. Bud break induction in kiwifruit vines cultivated in an organic system by the biological method of single node cutting. Bragantia-Campinas 2020, 79, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Kotb, H.R.M.; El-Abd, M.A.M.; Salama, A. Response of “White Robin” peach trees cv. (Prunus persica L.) to cultivation under plastic covering conditions and foliar application by hydrogen cyanamide and garlic extract. J. Plant Prod. 2019, 10, 1187–1194. [Google Scholar] [CrossRef]
- Pérez, F.J.; Vergara, R.; Or, E. On the mechanism of dormancy release in grapevine-buds: A comparative study between hydrogen cyanamide and sodium azide. Plant Growth Regul. 2009, 59, 145–152. [Google Scholar] [CrossRef]
- Tohbe, M.; Mochioka, R.; Horiuchi, S.; Ogata, T.; Shiozaki, S.; Kurooka, H. Roles of ACC and glutathione during breaking of dormancy in grapevine buds by high temperature treatment. Engei Gakkai Zasshi 1998, 67, 897–901. [Google Scholar] [CrossRef]
- Vargas-Arispuro, I.; Corrales-Maldonado, C.; Martínez-Téllez, M.Á. Compounds derived from garlic as bud induction agent in organic farming of table grape. Chil. J. Agric. Res. 2008, 68, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Ziosi, V.; Giovannetti, G.; Vitali, F.; Di Nardo, A.; Costa, G.; Laghezza, L. Bluprins® induces advanced bud release from dormancy in sweet cherry. Acta Hortic. 2013, 1009, 71–77. [Google Scholar] [CrossRef]
- Ziosi, V.; Di Nardo, A.; Fontana, A.; Vitali, F.; Costa, G. Effect of Bluprins® application on bud release from dormancy in kiwifruit, cherry, and table grape. In Advances in Plant Dormancy; Anderson, J.V., Ed.; Springer International Publishing AG: Cham, Switzerland, 2015; pp. 301–308. [Google Scholar]
- Saffari, P.; Majd, A.; Jonoubi, P.; Najafi, F. Effect of treatments on seed dormancy breaking, seedling growth, and seedling antioxidant potential of Agrimonia eupatoria L. J. Appl. Res. Medic. Aromat. Plants 2021, 20, 100282. [Google Scholar] [CrossRef]
- Khalil, U.; Rehman, K.; Wang, W.; Zheng, H.; Faheem, M.; Iqbal, S.; Shen, Z.G.; Tao, J. Role of hydrogen cyanamide (HC) in grape bud dormancy release: Proteomic approach. Biotech 2020, 10, 229. [Google Scholar] [CrossRef]
- Urtasun, M.M.; Giamminola, E.M.; Baskin, C.C.; de Viana, M.L.; Morandini, M.N.; Lamas, C.Y. Dormancy release, germination and ex situ conservation of the southern highland papaya (Vasconcellea quercifolia, Caricaceae), a wild crop relative. Sci. Hortic. 2020, 263, 109134. [Google Scholar] [CrossRef]
- Golmohammadzadeh, S.; Zaefarian, F.; Rezvani, M. Priming techniques, germination and seedling emergence in two Papaver species (P. rhoeas L. and P. dubium L., Papaveraceae). Braz. J. Bot. 2020, 43, 503–512. [Google Scholar] [CrossRef]
- Guijarro-Real, C.; Adalid-Martínez, A.M.; Gregori-Montaner, A.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Factors affecting germination of Diplotaxis erucoides and their effect on selected quality properties of the germinated products. Sci. Hortic. 2020, 261, 109013. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Walck, J.L.; Tao, J. Non-deep simple morphophysiological dormancy in seeds of Angelica keiskei (Apiaceae). Sci. Hortic. 2019, 255, 202–208. [Google Scholar] [CrossRef]
- Vajari, M.A.; Moghadam, J.F.; Eshghi, S. Influence of late season foliar application of urea, boric acid and zinc sulfate on nitrogenous compounds concentration in the bud and flower of Hayward kiwifruit. Sci. Hortic. 2018, 242, 137–145. [Google Scholar] [CrossRef]
- Babita Rana, V.S. Influence of Dormex on the phenology of flowering and fruit production of Hayward kiwifruit. Int. J. Chem. Stud. 2018, 6, 2941–2945. [Google Scholar]
- Hoeberichts, F.A.; Povero, G.; Ibañez, M.; Strijker, A.; Pezzolato, D.; Mills, R.; Piaggesi, A. Next Generation Sequencing to characterise the breaking of bud dormancy using a natural biostimulant in kiwifruit (Actinidia deliciosa). Sci. Hortic. 2017, 225, 252–263. [Google Scholar] [CrossRef]
- Grzesik, M.; Górnik, K.; Janas, R.; Lewandowki, M.; Romanowska-Duda, Z.; van Duijn, B. High efficiency stratification of apple cultivar Ligol seed dormancy by phytohormones, heat shock and pulsed radio frequency. J. Plant Physiol. 2017, 219, 81–90. [Google Scholar] [CrossRef]
- Sudawan, B.; Chang, C.-S.; Chao, H.-F.; Ku, M.S.B.; Yen, Y.-F. Hydrogen cyanamide breaks grapevine bud dormancy in the summer through transient activation of gene expression and accumulation of reactive oxygen and nitrogen species. BMC Plant Biol. 2016, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Jang, B.K.; Lee, C.H. Breaking combinational dormancy of Rhus javanica L. seeds in South Korea: Effect of mechanical scarification and cold-moist stratification. S. Afr. J. Bot. 2020, 133, 174–177. [Google Scholar] [CrossRef]
- Kimura, E.; Fransen, S.C.; Collins, H.P.; Guy, S.O.; Johnston, W.J. Breaking seed dormancy of switchgrass (Panicum virgatum L.): A review. Biomass Bioenergy 2015, 80, 94–101. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, X.; Gao., Y.; Li, K.; Guo, W. Transcriptome-based identification of AP2/ERF family genes and their cold-regulated expression during the dormancy phase transition of Chinese cherry flower buds. Sci. Hortic. 2021, 275, 109666. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Zhang, Y.; Baskin, J.M.; Baskin, C.C.; Xiong, Z.; Tao, J. A review of the seed biology of Paeonia species (Paeoniaceae), with particular reference to dormancy and germination. Planta 2019, 249, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Svystun, T.; Al-Dahmash, B.; Jonsson, A.M.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Considine, M.J.; Foyer, C.H. Redox regulation of plant development. Antioxid. Redox Signal. 2014, 21, 1305–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Halaly, T.; Acheampong, A.K.; Takebayashi, Y.; Jikumaru, Y.; Kamiya, Y.; Or, E. Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism. J. Exp. Bot. 2015, 66, 1527–1542. [Google Scholar] [CrossRef]
- Ben Mohamed, H.; Vadel, A.M.; Geuns, J.M.C.; Khemira, H. Carbohydrate changes during dormancy release in Superior Seedless grapevine cuttings following hydrogen cyanamide treatment. Sci. Hortic. 2012, 140, 19–25. [Google Scholar] [CrossRef]
- Rinne, P.L.H.; Welling, A.; Vahala, J.; Ripel, L.; Ruonala, R.; Kangasjärvi, J.; van der Schoot, C. Chilling of dormant buds hyperinduces flowering locus T and recruits GA-inducible 1, 3-β-glucanases to reopen signal conduits and release dormancy in Populus. Plant Cell 2011, 23, 130–146. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, W.; Gao, Z.; Wang, L.; Zhong, W.; Ni, Z.; Zhang, Z. Comparative proteomic and transcriptomic approaches to address the active role of GA4 in Japanese apricot flower bud dormancy release. J. Ex. Bot. 2013, 64, 4953–4966. [Google Scholar] [CrossRef]
- Zhuang, W.; Gao, Z.; Wen, L.; Huo, X.; Cai, B.; Zhang, Z. Metabolic changes upon flower bud break in Japanese apricot are enhanced by exogenous GA4. Hortic. Res. 2015, 2, 15046. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, L.M.; Volaire, F.A. Are winter and summer dormancy symmetrical seasonal adaptive strategies? The case of temperate herbaceous perennials. Ann. Bot. 2017, 119, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Lloret, A.; Badenes, M.L.; Ríos, G. Modulation of dormancy and growth responses in reproductive buds of temperate trees. Front. Plant Sci. 2018, 9, 1368. [Google Scholar] [CrossRef] [Green Version]
- Martín-Fontecha, E.S.; Tarancon, C.; Cubas, P. To grow or not to grow, a power-saving program induced in dormant buds. Curr. Opin. Plant Biol. 2018, 41, 102–109. [Google Scholar] [CrossRef]
- Falavigna, V.S.; Guitton, B.; Costes, E.; Andrés, F.I. Want to (bud) break free: The potential role of DAM and SVP-like genes in regulating dormancy cycle in temperate fruit trees. Front. Plant Sci. 2019, 9, 1990. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Vimont, N.; Fouché, M.; Campoy, J.A.; Tong, M.; Arkoun, M.; Yvin, J.-C.; Wigge, P.A.; Dirlewanger, E.; Cortijo, S.; Wenden, B. From bud formation to flowering: Transcriptomic state defines the cherry developmental phases of sweet cherry bud dormancy. BMC Genom. 2019, 20, 974. [Google Scholar] [CrossRef] [Green Version]
- Buijs, G.A. Perspective on Secondary Seed Dormancy in Arabidopsis thaliana. Plants 2020, 9, 749. [Google Scholar] [CrossRef] [PubMed]
- Goeckeritz, C.; Hollender, C.A. There is more to flowering than those DAM genes: The biology behind bloom in rosaceous fruit trees. Curr. Opin. Plant Biol. 2021, 59, 101995. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Darbyshire, R.; Dirlewanger, E.; Quero-García, J.; Wenden, B. Yield potential definition of the chilling requirement reveals likely underestimation of the risk of climate change on winter chill accumulation. Int. J. Biometeorol. 2019, 63, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Dai, J.; Ranjitkar, S.; Yu, H.; Xu, J.; Luedeling, E. Chilling and heat requirements for flowering in temperate fruit trees. Int. J. Biometeorol. 2014, 58, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Ruiz, D.; Allderman, L.; Cook, N.; Egea, J. The fulfilment of chilling requirements and the adaptation of apricot (Prunus armeniaca L.) in warm winter climates: An approach in Murcia (Spain) and the Western Cape (South Africa). Eur. J. Agron. 2012, 37, 43–55. [Google Scholar] [CrossRef]
- El Yaacoubi, A.; Malagi, G.; Oukabli, A.; Citadin, I.; Hafidi, M.; Bonhomme, M.; Legave, J.-M. Differentiated dynamics of bud dormancy and growth in temperate fruit trees relating to bud phenology adaptation, the case of apple and almond trees. Int. J. Biometeorol. 2016, 60, 1695–1710. [Google Scholar] [CrossRef]
- Kaufmann, H.; Blanke, M. Performance of three numerical models to assess winter chill for fruit trees—A case study using cherry as model crop in Germany. Reg. Environ. Chang. 2017, 17, 715–723. [Google Scholar] [CrossRef]
- Nasrabadi, M.; Ramezanian, A.; Eshghi, S.; Sarkhosh, A. Chilling and heat requirement of pomegranate (Punica granatum L.) trees grown under sustained deficit irrigation. Sci. Hortic. 2020, 263, 109117. [Google Scholar] [CrossRef]
- Kühn, N.; Ormeño-Nuñez, J.; Jaque-Zamora, G.; Pérez, F.J. Photoperiod modifies the diurnal expression profile of VvPHYA and VvPHYB transcripts in field-grown grapevine leaves. J. Plant Physiol. 2009, 166, 1172–1180. [Google Scholar] [CrossRef]
- Javanshah, A.; Alipour, H.; Hadavi, F. A model for assessing the chill units received in Kerman and Rafsanjan areas. Acta Hortic. 2006, 726, 221–225. [Google Scholar] [CrossRef]
- Jangra, M.S.; Sharma, J.P. Climate resilient apple production in Kullu valley of Himachal Pradesh. Int. J. Farm Sci. 2013, 3, 91–98. [Google Scholar]
- Esper, J.; Cook, E.R.; Schweingruber, F.H. Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 2002, 295, 2250–2253. [Google Scholar] [CrossRef] [Green Version]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. IPCC (2013): Climate Change The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; pp. 1–1535. [Google Scholar]
- Perez-Lopez, D.; Ribas, F.; Moriana, A.; Rapoport, H.F.; De Juan, A. Influence oftemperature on the growth and development of olive (Olea europaea L.) trees. J. Hortic. Sci. Biotechnol. 2008, 83, 171–176. [Google Scholar] [CrossRef]
- Orlandi, F.; Garcia-Mozo, H.; Galán, C.; Romano, B.; de la Guardia, C.D.; Ruiz, L.; del Mar Trigo, M.; Dominguez-Vilches, E.; Fornaciari, M. Olive flowering trends in a large Mediterranean area (Italy and Spain). Int. J. Biometeorol. 2010, 54, 151–163. [Google Scholar] [CrossRef]
- Ezzat, A.; Amriskó, L.G.; Mikita, G.; Nyéki, T.; Soltész, J.; Szabó, Z.M. Variation between some apricot varieties in regard to flowering phenology in Boldogkôváralja, Hungary. Int. J. Hortic. Sci. 2012, 18, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Jana, B.R.; Das, B. Effect of dormancy breaking chemicals on flowering, fruit set and quality in Asian pear (Pyrus pyrifolia L.). Afr. J. Agric. Res. 2014, 9, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Yuri, J.A.; Moggia, C.; Sepulveda, A.; Poblete-Echeverría, C.; Valdés-Gómez, H.; Torres, C.A. Effect of cultivar, rootstock, and growing conditions on fruit maturity and postharvest quality as part of a six-year apple trial in Chile. Sci. Hortic. 2019, 253, 70–79. [Google Scholar] [CrossRef]
- Rana, R.S.; Bhagat, R.M.; Kalia, V.; Lal, H. Impact of climate change on shift of apple belt in Himachal Pradesh. In Proceedings of the Workshop on Impact of Climate Change on Agriculture, Ahmedabad, India, 17–18 December 2009; ISPRS Ar-chives 38-8/3. Panigrahy, S., Ray, S.S., Parihar, J.S., Eds.; Excel India Publishers: New Delhi, India, 2009; pp. 131–137. [Google Scholar]
- Stockle, C.O.; Nelson, R.L.; Higgins, S.; Brunner, J.; Grove, G. Assessment of climate change impact on Eastern Washington agriculture. Clim. Chang. 2010, 102, 77–102. [Google Scholar] [CrossRef] [Green Version]
- Byrne, D.H.; Sherman, W.B.; Bacon, T.A. Stone fruit genetic pool and its exploitation for growing under warm climatic conditions. In Temperate Fruit Crops in Warm Climates; Erez, A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 157–230. [Google Scholar]
- Mohamed, A.K.A. The effect of chilling, defoliation and hydrogen cyanamide on dormancy release, bud break and fruiting of Anna apple cultivar. Sci. Hortic. 2008, 118, 25–32. [Google Scholar] [CrossRef]
- Fernandez, E.; Whitney, C.; Luedeling, E. The importance of chill model selection—A multi-site analysis. Eur. J. Agron. 2020, 119, 126103. [Google Scholar] [CrossRef]
- Weinberger, J. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Richardson, E.; Seeley, S.D.; Walker, D. A model for estimating the completion of rest for Redhaven and Elberta peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Cabré, F.; Nuñez, M. Impacts of climate change on viticulture in Argentina. Reg. Environ. Chang. 2020, 20, 12. [Google Scholar] [CrossRef]
- Benmoussa, H.; Luedeling, E.; Ghrab, M.; Ben Mimoun, M. Severe winter chill decline impacts Tunisian fruit and nut orchards. Clim. Chang. 2020, 162, 1249–1267. [Google Scholar] [CrossRef]
- Landsberg, J.J. Apple fruit bud development and growth; analysis and an empirical model. Ann. Bot. 1974, 38, 1013–1023. [Google Scholar] [CrossRef]
- Gilreath, P.R.; Buchanan, D.W. Rest prediction model for low-chilling Sungold nectarine. J. Am. Soc. Hortic. Sci. 1981, 106, 426–429. [Google Scholar]
- Shaltout, A.D.; Unrath, C.R. Rest completion prediction model for Starkrimson Delicious apples. J. Am. Soc. Hortic. Sci. 1983, 108, 957–961. [Google Scholar]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants - mathematical-analysis of a 2-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Linsley-Noakes, G.; Allan, P.; Matthee, G. Modification of rest completion prediction models for improved accuracy in South African stone fruit orchards. J. S. Afr. Soc. Hort. Sci. 1994, 4, 13–15. [Google Scholar]
- Guak, S.; Neilsen, D. Chill Unit models for predicting dormancy completion of floral buds in apple and sweet cherry. Hort. Environ. Biotechnol. 2013, 54, 29–36. [Google Scholar] [CrossRef]
- Jones, H.G.; Hillis, R.M.; Gordon, S.L.; Brennan, R.M. An approach to the determination of winter chill requirements for different Ribes cultivars. Plant Biol. 2013, 15, 18–27. [Google Scholar] [CrossRef]
- Jones, H.G.; Gordon, S.L.; Brennan, R.M. Chilling requirement of Ribes cultivars. Front. Plant Sci. 2015, 5, 767. [Google Scholar] [CrossRef]
- Luedeling, E. chillR: Statistical Methods for Phenology Analysis in Temperate Fruit Trees. R Package Version 0.70.21. Available online: https://cran.r-project.org/web/packages/chillR/ (accessed on 18 April 2021).
- Parkes, H.; Darbyshire, R.; White, N. Chilling requirements of apple cultivars grown in mild Australian winter conditions. Sci. Hortic. 2020, 260, 108858. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Ben Mimoun, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Egea, J.A.; Egea, J.; Ruiz, D. Reducing the uncertainty on chilling requirements for endodormancy breaking of temperate fruits by data-based parameter estimation of the dynamic model: A test case in apricot. Tree Physiol. 2020, 41, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Dai, J.; Ge, Q.; Wang, H.; Tao, Z. Comparison of chilling and heat requirements for leaf unfolding in deciduous woody species in temperate and subtropical China. Int. J. Biometeorol. 2020, 65, 393–403. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, T.; Frauenfeld, O.W.; Wang, K.; Sun, W.; Luo, J. Evaluation and quantification of surface air temperature over Eurasia based on CMIP5 models. Clim. Res. 2019, 77, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Paltineanu, C.; Chitu, E. Climate change impact on phenological stages of sweet and sour cherry trees in a continental climate environment. Sci. Hortic. 2020, 261, 109011. [Google Scholar] [CrossRef]
- Boudichevskaia, A.; Kumar, G.; Sharma, Y.; Kapoor, R.; Singh, A.K. Challenges and strategies for developing climate-smart apple varieties through genomic approaches. In Genomic Designing of Climate-Smart Fruit Crops; Kole, C., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 23–71. [Google Scholar] [CrossRef]


| Crop | Country | Impact | Ref. |
|---|---|---|---|
| Apple (Malus × domestica Borkh) | Morocco | Late blooming, extended blooming period, extended period from blooming to harvest, and reduced fruit quality were recorded. | [29] |
| Apple (Malus × domestica Borkh) | India | Negative effects on cumulative chilling requirements and final fruit set as well as variation in flowering onset and duration were recorded. | [30] |
| Apple (Malus × domestica Borkh) and Almond (Prunus dulcis (Mill.) Webb) | Spain | A phenology disruption was recorded due to changes in chilling requirements and mean temperature threshold for spring growth. | [31] |
| Apple (Malus × domestica Borkh) | Germany | Insufficient chilling requirements negatively affected flowering date and quantity. Early spring frost damages the new flush. | [32] |
| Apple (Malus × domestica Borkh) | Iran | Expected temperature range of tree growth will change from 11.6–27.3 to 16.7–33.4 °C by 2090, and 46.7% of orchards will be lost. | [33] |
| Apple (Malus × domestica Borkh) and other fruit crops | USA | Increased frequencies of longer warmer summers can increase the insect generations during the season and the over-wintering survival of insect pests and their natural enemies. | [34] |
| Apple (Malus × domestica Borkh) | Spain | Continuous vulnerability to insufficient chilling will lead to late blooming by the end of the 21st century. | [35] |
| Apple (Malus × domestica Borkh) | India | A negative fruit production rate was shown from 2005 to 2014 by 0.183 ton ha−1 year−1. | [36] |
| Apple (Malus × domestica Borkh) | India | Insufficient chilling negatively affected flower bud induction, fruit set, yield, color, firmness, and taste. Cold temperature during flowering affected pollination. Insufficient chilling also posed serious problems such asapple scab epidemics, red spider gradation, and premature leaf fall. | [10] |
| Crop | Country | Impact | Ref. |
|---|---|---|---|
| Grapevine (Vitis vinifera L.) | Spain | Fruit quality (mainly contents of phenolic compounds and total soluble solids) of some old genotypes was quite stable at CO2 (700 ppm) and temperature (4 °C). | [37] |
| Peach Prunus persica L. | Spain | Decreased stomatal conductance, alleviated drought stress effects, and an increased photosynthetic rate were recorded. | [38] |
| Grapevine: (Vitis vinifera L.), cv. Tempranillo | Spain | CO2 (700 ppm) decreased the combining effects of temperature on the anthocyanin content and sugar ratio. Elevated CO2 and air temperature was expected to advance grape maturity and reduce the elapsed period between fruit set and maturity. | [39] |
| Grapevine (Vitis vinifera L.) | Croatia | Grapevine phenology was advanced by 4.1 and 6.8 days at 4 °C and ambient temperature, respectively, and early fruit maturity and ripening were reported. | [40] |
| Grapevine (Vitis vinifera L.) | Spain | Elevated CO2 (700 ppm) for 10 days decreased chlorophyll content, but the effect dissipated after longer exposure for 20 days. Decreased N content and no effect on chlorophyll a/b ratio were reported. | [41] |
| Peach (Prunus persica L.) Batsch | China | Elevated CO2 (360 ppm) increased fruit sucrose, lactones, and flavor constituents (linoleic and linolenic acid, pyruvic acid, and the precursor of volatile compounds) but decreased malic acid. | [42] |
| Pears (Pyrus pyrifolia Nakai), cv. Niitaka | Korea | Elevated CO2 (700 ppm) increased fruit weight, photosynthetic rate, and total soluble solids content at harvest. | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salama, A.-M.; Ezzat, A.; El-Ramady, H.; Alam-Eldein, S.M.; Okba, S.K.; Elmenofy, H.M.; Hassan, I.F.; Illés, A.; Holb, I.J. Temperate Fruit Trees under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions. Horticulturae 2021, 7, 86. https://doi.org/10.3390/horticulturae7040086
Salama A-M, Ezzat A, El-Ramady H, Alam-Eldein SM, Okba SK, Elmenofy HM, Hassan IF, Illés A, Holb IJ. Temperate Fruit Trees under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions. Horticulturae. 2021; 7(4):86. https://doi.org/10.3390/horticulturae7040086
Chicago/Turabian StyleSalama, Abdel-Moety, Ahmed Ezzat, Hassan El-Ramady, Shamel M. Alam-Eldein, Sameh K. Okba, Hayam M. Elmenofy, Islam F. Hassan, Attila Illés, and Imre J. Holb. 2021. "Temperate Fruit Trees under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions" Horticulturae 7, no. 4: 86. https://doi.org/10.3390/horticulturae7040086