
Postharvest Treatment of ‘Florida Prince’ Peaches with a Calcium Nanoparticle–Ascorbic Acid Mixture during Cold Storage and Its Effect on Antioxidant Enzyme Activities




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Materials and Postharvest Treatments
2.2. Synthesis of Metal Calcium Nanoparticles (CaNPs) with Ascorbic Acid (AA)
2.3. CaNP–AA Application Protocol
2.4. Chilling Injury Index, Water Loss%, and Fruit Skin Color
2.5. Total Soluble Solid Content (SSC%), Total Acidity (TA%), and SSC/TA Ratio
2.6. Fruit Pigments and Fruit Firmness (N)
2.7. Antioxidant Enzyme Activities (AEAs)
2.8. Lipid Peroxidation, Malondialdehyde (MDA) Accumulation, and Ion Leakage%
2.9. Ethylene and Respiration Assessment
2.10. H2O2 and O2•− Production Rate and DPPH Reduction
2.11. Statistical Analysis
3. Results
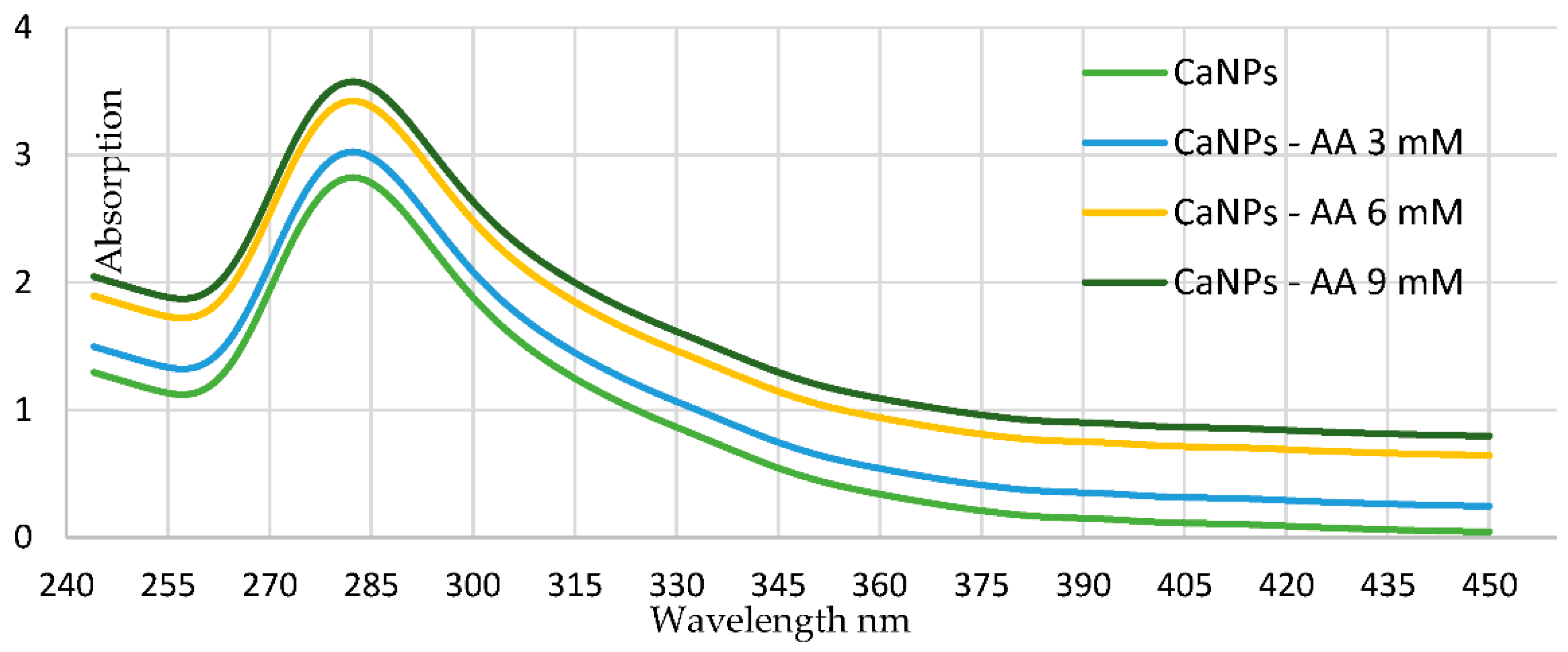
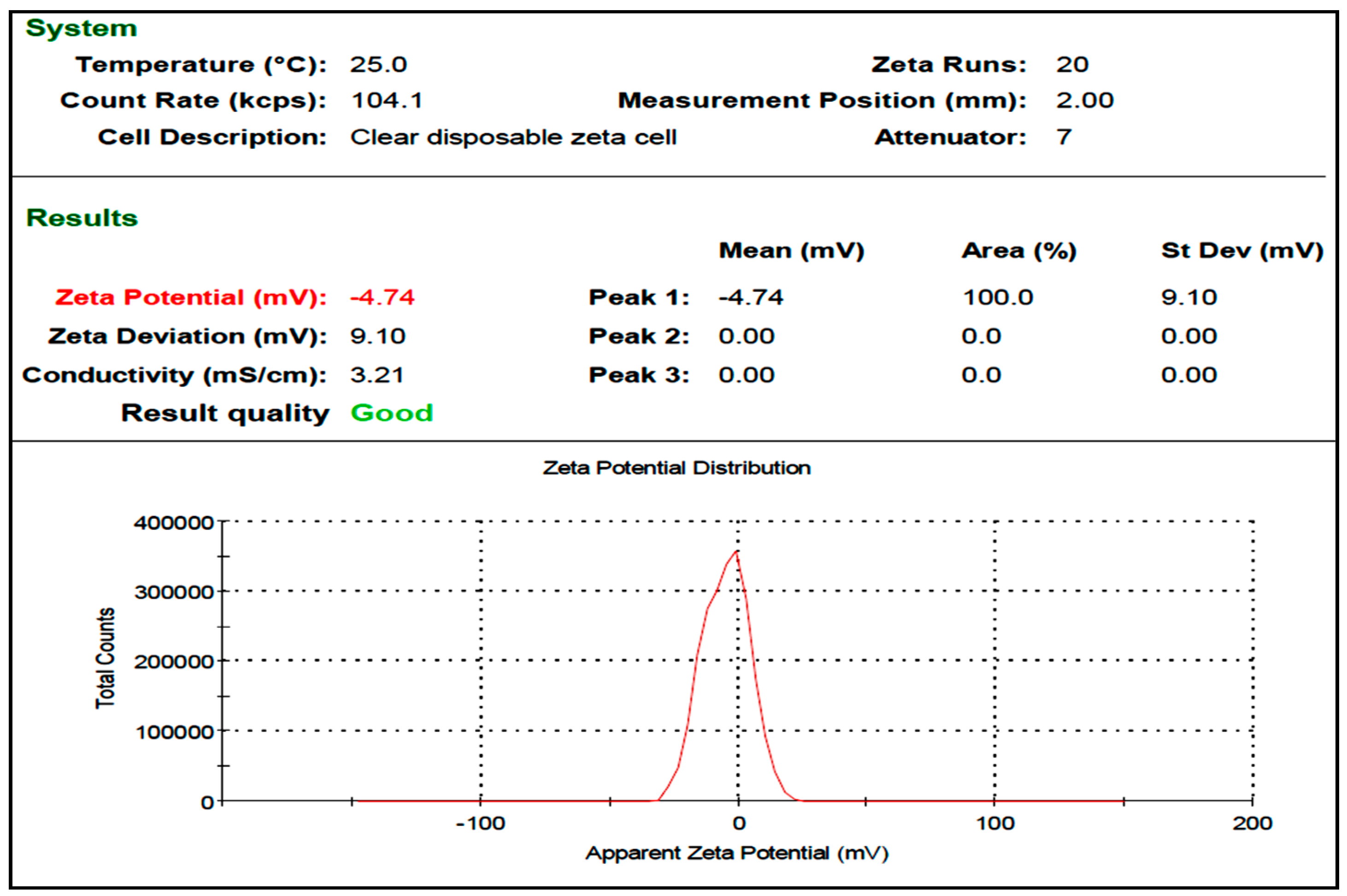
3.1. Synthesis of Metal Calcium Nanoparticles (CaNPs)
3.2. CI Index, Water Loss%, and Peach Skin Color
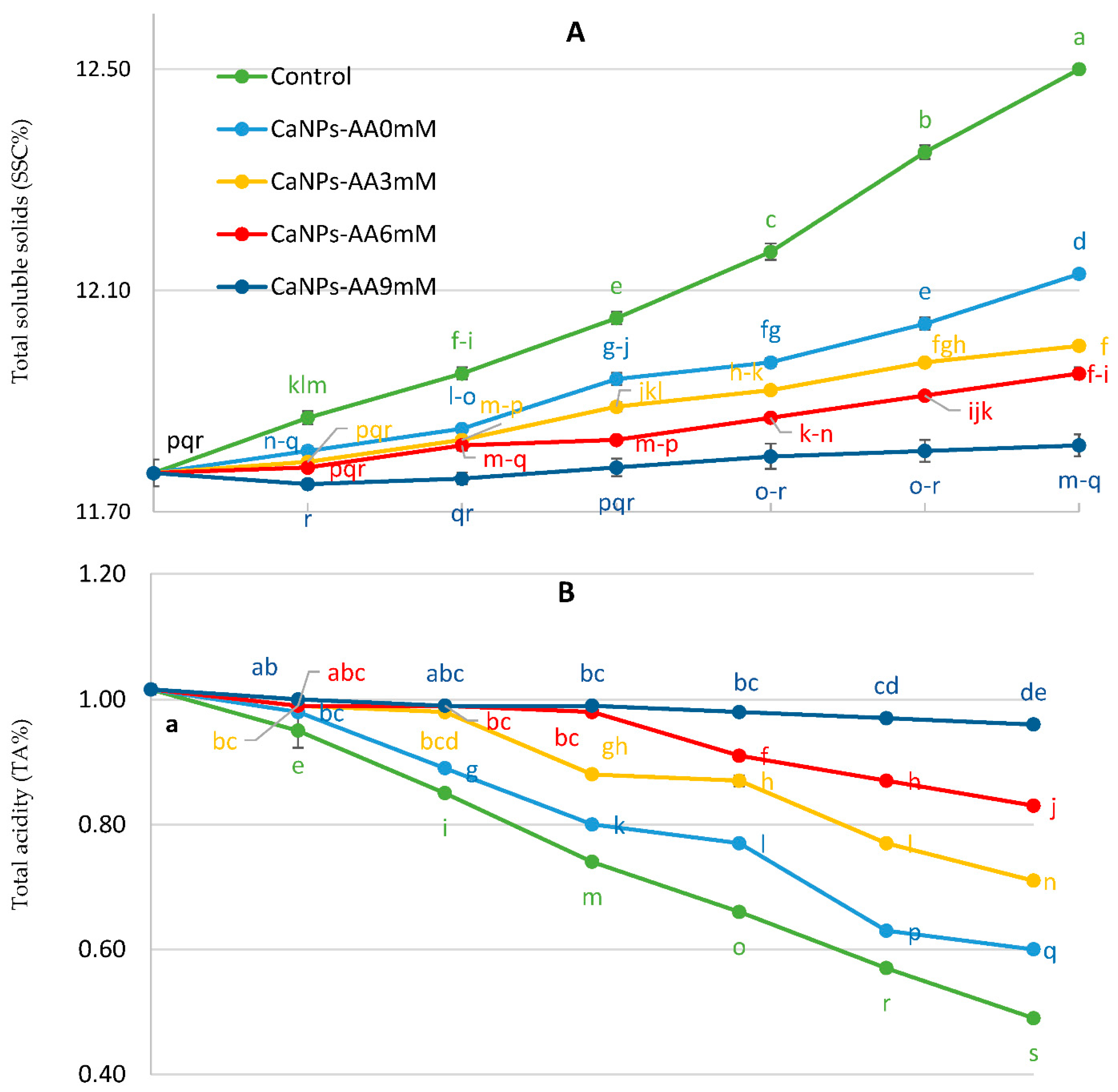
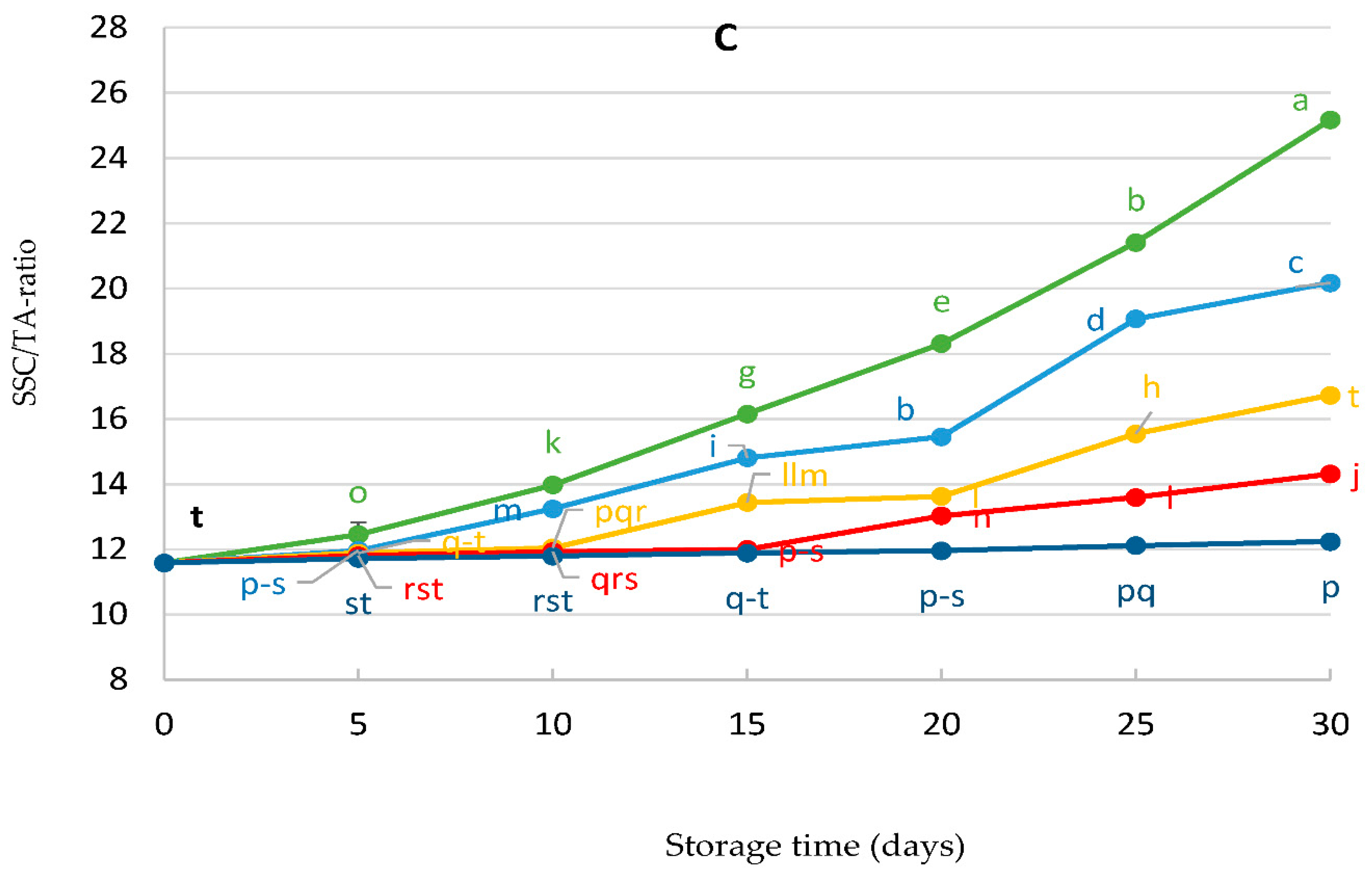
3.3. Total Soluble Solids (SSC%), Total Acidity (TA%), and SSC/TA Ratio
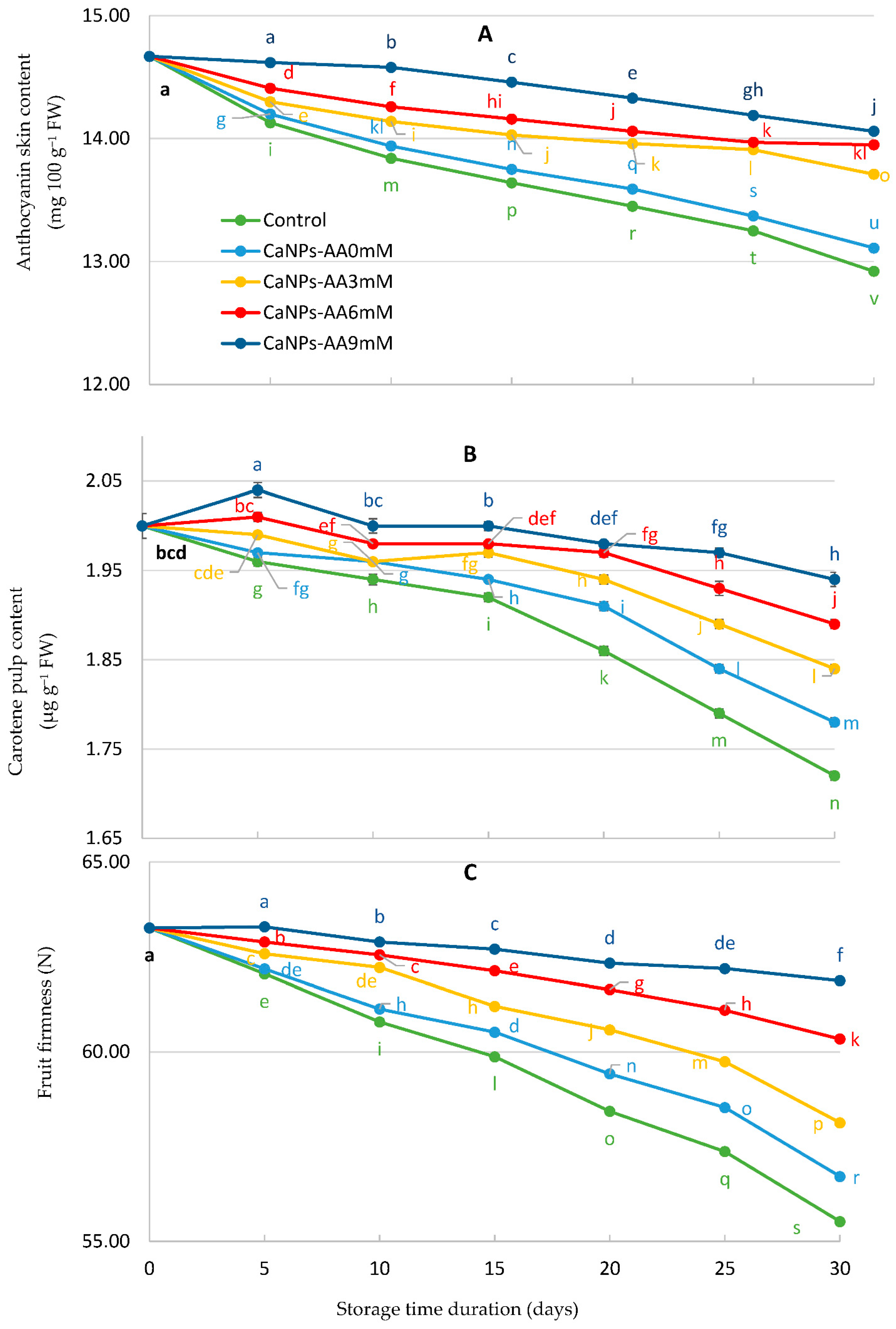
3.4. Fruit Pigments (Anthocyanin and Carotene) and Fruit Firmness (N)
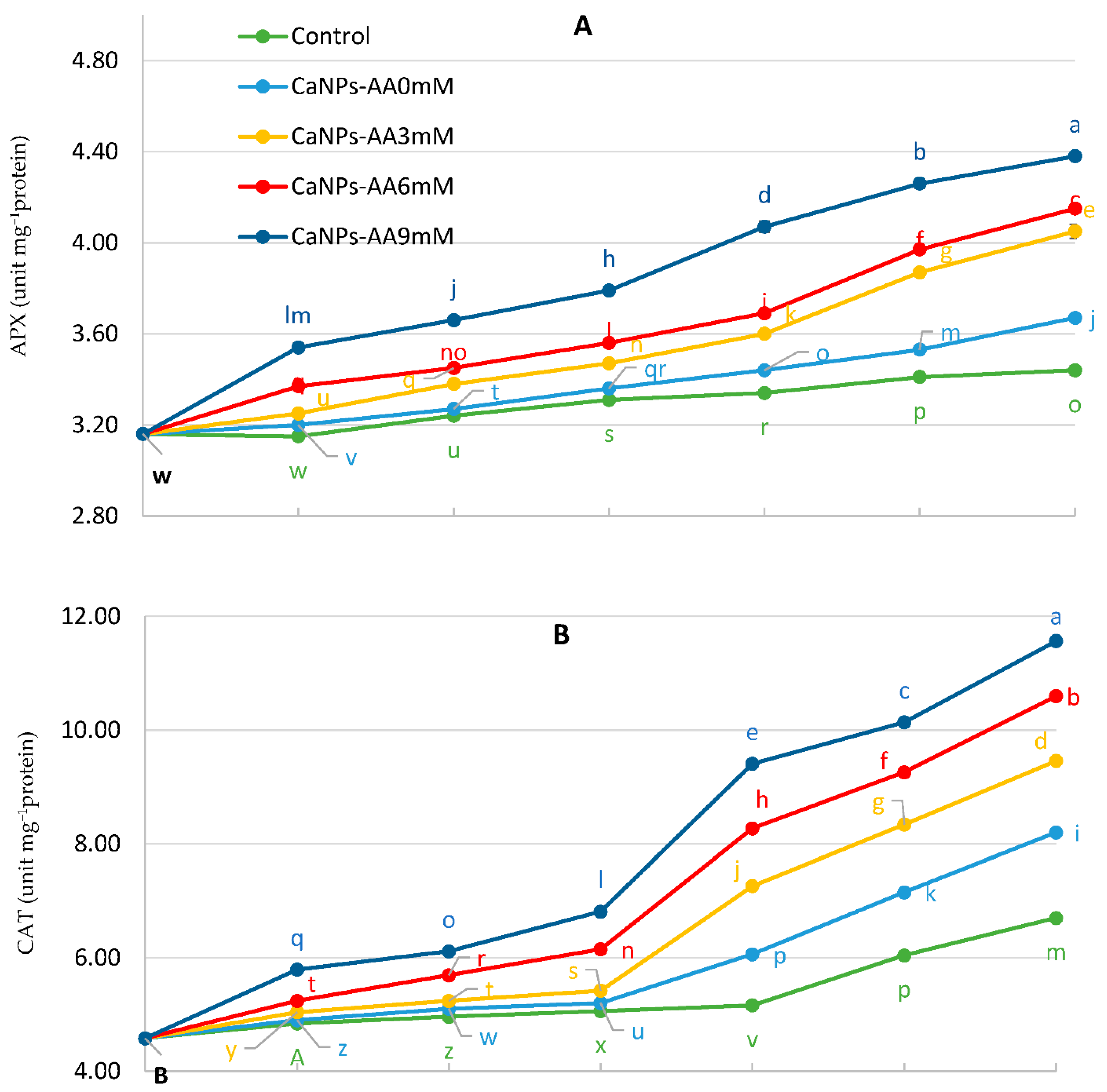
3.5. Antioxidant Enzyme Activity (AEA)
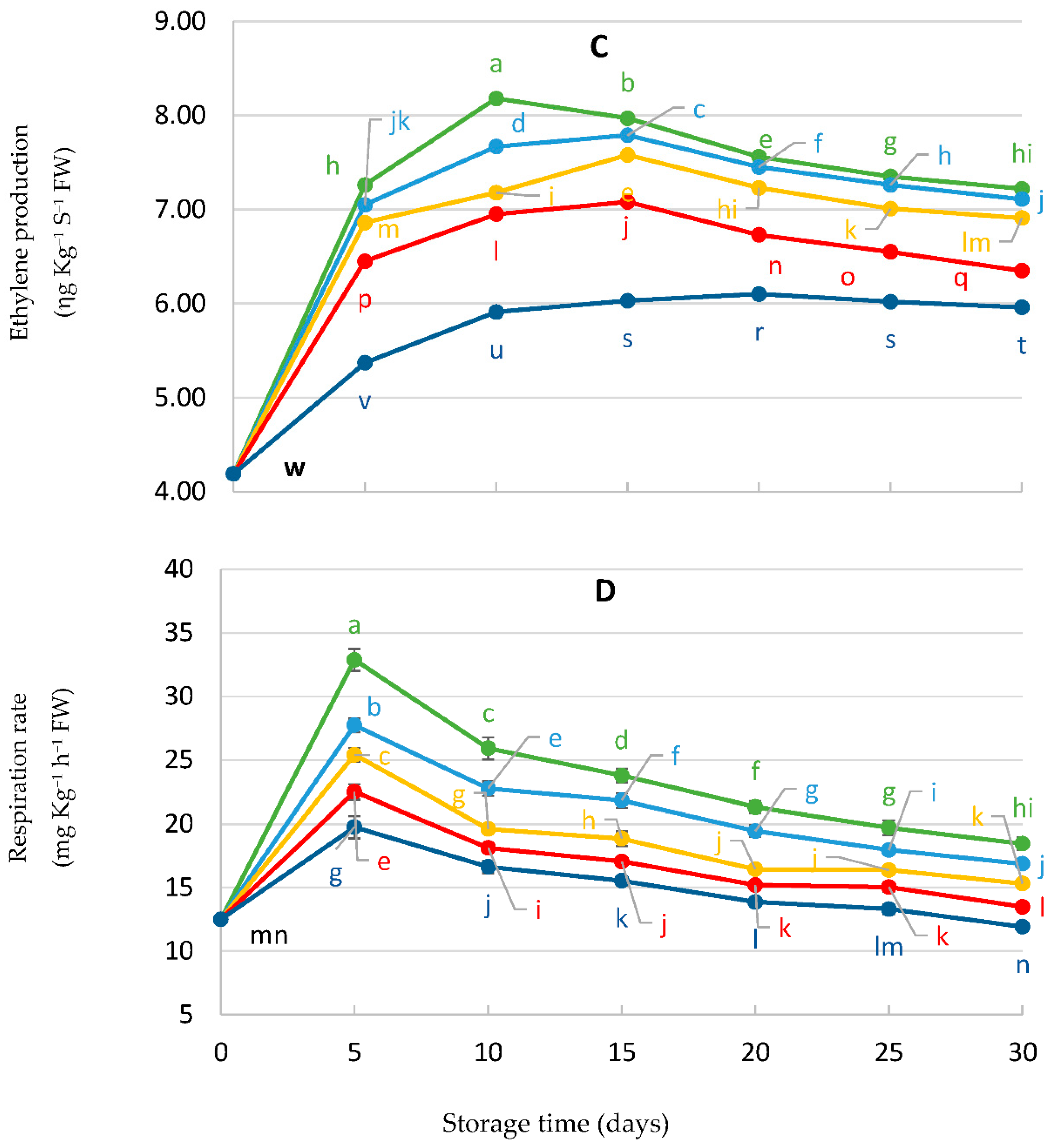
3.6. Estimation of Malondialdehyde (MDA), Ion Leakage (IL%), Ethylene Production, and Respiration Rate
3.7. H2O2 and O2•− Production and DPPH Reduction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- El-Boray, M.S.; Shalan, A.M.; Khouri, Z.M. Performance of Peach Trees cv. Florida Prince under Different Foliar Concentrations of NPK-humate in Presence or Absence of Adjuvants. Trends Hortic. Res. 2016, 6, 5–17. [Google Scholar]
- FAOSTAT. Peach Production in Egypt; FAO: Rome, Italy, 2017. [Google Scholar]
- Jin, P.; Zhu, H.; Wang, L.; Shen, T.; Zheng, Y. Oxalic acid alleviates chilling injury in peach fruit by regulating energy metabolism and fatty acid contents. Postharvest Biol. Technol. 2014, 161, 87–93. [Google Scholar] [CrossRef]
- Wang, C.Y. Chilling Injury of Horticultural Crops; CRC Press: Boca Raton, FL, USA, 1990. [Google Scholar]
- Lurie, S.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Brummell, D.A.; Dal Cin, V.; Crisoto, C.H.; Labavith, J.M. Cell wall metabolism during the development of chilling injury in cold-stored peach fruit: Association of mealiness with arrested disassembly of cell wall pectins. J. Exp. Bot. 2004, 55, 2041–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Hu, Z.; Zheng, Y.; Lu, B. Synergistic effect of heat treatment and salicylic acid on alleviating internal browning in cold-stored peach fruit. Postharvest Biol. Technol. 2010, 58, 93–97. [Google Scholar] [CrossRef]
- Fernandez-Trujillo, J.P.; Martinez, J.A.; Artes, F. Modified atmosphere pack affect the incidence of cold storage disorder and keep ‘flat’ peach quality. Food Res. Int. 1998, 31, 571–579. [Google Scholar] [CrossRef]
- Ebel, R.C.; Woods, F.M.; Himelrick, D. Effect of UV-C on ripening and postharvest quality of peach. HortScience 1999, 34, 504. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Zheng, Y.H.; Zhang, Y.F.; Wang, F.; Zhang, L.; Lu, Z.X. Methyl jasmonate reduces chilling injury and maintains postharvest quality in peaches. Agric. Sci. China 2003, 11, 1246–1252. [Google Scholar]
- Girardi, C.L.; Corrent, A.R.; Lucchetta, L.; Zanuzo, M.R.; Da Costa, T.S.; Brachmann, A.; Twymand, R.M.; Nora, F.R.; Nora, L.; Silva, J.A.; et al. Effect of ethylene, intermittent warming and controlled atmosphere on postharvest quality and occurrence of woolliness in peach (Prunus persic cv. Chiripa) during cold storage. Postharvest Biol. Technol. 2005, 38, 25–33. [Google Scholar] [CrossRef]
- Wang, L.; Chen, S.; Kong, W.; Li, S.; Archbold, D.D. Salicylic acid pretreatment alleviates chilling injury and affects the antioxidant system and heat shock proteins of peaches during cold storage. Postharvest Biol. Technol. 2006, 41, 244–251. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; Ameer, N.M. Performance of calcium nanoparticles blending with ascorbic acid and alleviation internal browning of ‘Hindi Be-Sennara’ mango fruit at a low temperature. Sci. Hortic. 2019, 254, 199–207. [Google Scholar] [CrossRef]
- Wismer, W.V.; Worthing, W.M.; Yada, R.Y.; Marangoni, A.G. Membrane lipid dynamics and lipid peroxidation in the early stages of low-temperature sweetening in tubers of Solanum tuberosum. Physiol. Plant. 1998, 102, 396–410. [Google Scholar] [CrossRef]
- Foyer, C.H.; Ruban, A.V.; Noctor, G. Viewing oxidative stress through the lens of oxidative signaling rather than damage. Biochem. J. 2017, 474, 877–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, Z.; Nouri, M.Z.; Komatsu, S. Plant Cell Organelle Proteomics in Response to Abiotic Stress. J. Proteome Res. 2012, 11, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Lo’ay, A.A.; El-Khateeb, A.Y. Antioxidant enzyme activities and exogenous ascorbic acid treatment of ‘Williams’ banana during long-term cold storage stress. Sci. Hortic. 2018, 234, 210–219. [Google Scholar] [CrossRef]
- Purvis, A.C. Regulatin of oxidative stress in horticultural crops. HortiScience 2004, 39, 930–932. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Vasilakakis, M.; Diamantidis, G.; Mignani, I. Effect of calcium additives on physicochemical aspects of cell wall pectin and sensory attributes of canned peach (Prunus persica Batsch cv Andross). J. Sci. Food Agric. 2005, 85, 1773–1777. [Google Scholar] [CrossRef]
- Fan, X.; Niemera, B.A.; Matthheis, J.P.; Zhuang, H.; Olson, D.W. Quality of fresh-cut apple slices as affected by low-dose ionizing radiation and calcium ascorbate treatment. J. Food Sci. 2005, 70, 143–149. [Google Scholar] [CrossRef]
- Luna-Guzman, I.; Cantwell, M.; Barrett, D.M. Fresh-cut cantaloupe: Effects of calcium chloride dips and heat treatments on firmness and metabolic activity. Postharvest Biol. Technol. 1999, 17, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S. Calcium Chloride Treatment of Fruits and Vegetables; Tetre Technologies Inc.: The Woodlands, TX, USA, 2002. [Google Scholar]
- Yugandhar, P.; Savithramma, N. Biosynthesis, characterization and antimicrobial studies of green synthesized silver nanoparticles from fruit extract of Syzygium alternifolium (Wt.) Walp. an endemic, endangered medicinal tree taxon. Appl. Nanosci. 2016, 6, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Schirra, M. Behaviour of ‘star Ruby’ grapefruit under chilling and non-chilling storage temperature. Postharvest Biol. Technol. 1992, 2, 315–327. [Google Scholar] [CrossRef]
- Khojastehnazhand, M.; Omid, M.; Tabatabaeefar, A. Development of a lemon sorting system based on color and size. Afr. J. Plant Sci. 2010, 4, 122–127. [Google Scholar]
- Lo’ay, A.A.; Dawood, H.D. Active chitosan/PVA with ascorbic acid and berry quality of ‘Superior seedless’ grapes. Sci. Hortic. 2017, 224, 286–292. [Google Scholar] [CrossRef]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis Transcription Factor MYB12 Is a Flavonol-Specific Regulator of Phenylpropanoid Biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mínguez-Mosquera, M.I.; Jarén-Galán, M.; Hornero-Méndez, D.; Garrido-Fernández, J.; Gallardo-Guerrero, M.L.; Gandul-Rojas, B. Decoloration of vegetable oils and oleoresins with recovery of unaltered pigments. J. Am. Oil Chem. Soc. 1991, 68, 809–813. [Google Scholar] [CrossRef]
- Biehler, E.; Mayer, F.; Hoffmann, L.; Krause, E.; Bohn, T. Comparison of 3 Spectrophotometric Methods for Carotenoid Determination in Frequently Consumed Fruits and Vegetables. J. Food Sci. 2010, 75, C55–C61. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Teng, B.; Yu, Y. The firmness detection by excitation dynamic characteristics for peach. Food Control 2006, 17, 353–358. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. Assay of ascorbate-specific peroxidase. Methods Enzymol. 1984, 105, 427–429. [Google Scholar]
- Smith, I.S.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5%-dithiobis (2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar]
- Oberley, L.W.; Spitz, D.R. Nitroblue tetrazolium. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1986. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Iturbe-Ormaetxe, I.; Escuredo, P.R.; Arrese-Igor, C.; Becana, M. Oxidative damage in pea plants exposed to water deficit or paraquat. Plant Physiol. 1998, 116, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Lo’ay, A.A.; Taha, N.A.; El-Khateeb, Y.A. Storability of ‘Thompson Seedless’ grapes: Using biopolymer coating chitosan and polyvinyl alcohol blending with salicylic acid and antioxidant enzymes activities during cold storage. Sci. Hortic. 2019, 249, 314–321. [Google Scholar] [CrossRef]
- Lo’ay, A.A. Chilling Injury in Mangoes. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2005. [Google Scholar]
- Yang, H.; Wu, F.; Cheng, J. Reduced chilling injury in cucumber by nitricoxide and the antioxidant response. Food Chem. 2011, 127, 1237–1242. [Google Scholar] [CrossRef]
- Xu, M.; Dong, J.; Zhang, M.; Xu, X.; Sun, L. Cold-induced endogenous nitric oxide generation plays a role in chilling tolerance of loquat fruit during postharvest storage. Postharvest Biol. Technol. 2012, 65, 5–12. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Chen, L.-L.; Shan, W.; Cai, D.-L.; Chen, J.-Y.; Lu, W.-J.; Su, X.-G.; Kuang, J.-F. Postharvest application of glycine betaine ameliorates chilling injury in cold-stored banana fruit by enhancing antioxidant system. Sci. Hortic. 2021, 287, 110264. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; Doaa, M.H. The potential of vine rootstocks impacts on ‘Flame Seedless’ bunches behavior under cold storage and antioxidant enzyme activity performance. Sci. Hortic. 2020, 260, 108844. [Google Scholar] [CrossRef]
- Hodges, D.M.; Lester, G.E.; Munro, K.D.; Toivonen, P.M.A. Oxidative stress: Importance for postharvest quality. HortScience 2004, 39, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y. Reducing chilling injury and maintaining quality of horticultural crops with natural products and their derivatives. Acta Hortic. 2006, 712, 285–290. [Google Scholar] [CrossRef]
- Kondo, S.; Kittikorn, M.; Kanlayanarat, S. Preharvest antioxidant activities of tropical fruit and the effect of low temperature storage on antioxidants and jasmonates. Postharvest Biol. Technol. 2005, 36, 309–318. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, W.; Cao, J.; Ma, L. A combination of 1-methylcyclopropene treatment and intermittent warming alleviates chilling injury and affects phenolics and antioxidant activity of peach fruit during storage. Sci. Hortic. 2018, 229, 175–181. [Google Scholar] [CrossRef]
- Zhu, L.Q.; Zhou, J.; Zhu, S.H. Effect of a combination of nitric oxide treatment and intermittent warming on prevention of chilling injury of ‘Feicheng’ peach fruit during storage. Food Chem. 2010, 121, 165–170. [Google Scholar] [CrossRef]
- Giménez, M.; Olarte, C.; Sanz, S.; Lomas, C.; Echávarri, J.F.; Ayala, F. Influence of Packaging Films on the Sensory and Microbiological Evolution of Minimally Processed Borage (Borrago officinalis). J. Food Sci. 2003, 68, 1051–1058. [Google Scholar] [CrossRef]
- De Reuck, K.; Sivakumar, D.; Korsten, L. Integrated application of 1-methylcyclopropene and modified atmosphere packaging to improve quality retention of litchi cultivars during storage. Postharvest Biol. Technol. 2009, 52, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, E.A.; Niesperos, M.O.; Shaw, P.E.; Burns, J.K. Effect of coatings and prolonged storage conditions on fresh orange flavour volatiles, degrees brix and ascorbic acid levels. J. Agric. Food Chem. 1995, 43, 1321–1331. [Google Scholar] [CrossRef]
- Kader, A.A. Postharvest Technology of Horticulture Crops, 3rd ed.; University of California Agriculture & Natural Resources: Irvine, CA, USA, 2002. [Google Scholar]
- Shahid, M.N.; Abbasi, N.A. Effect of bee wax coatings on physiological changes in fruits of sweet orange CV. “Blood Red”. Sarhad J. Agric. 2011, 27, 385–394. [Google Scholar]
- Verma, P.; Dashora, L.K. Post-harvest physiconutritional changes in Kagzi Limes (Citrus aurantifolia S.) treated with selected oil emulsions and diphenyl. Plant Food Hum. Nutr. 2000, 55, 279–284. [Google Scholar] [CrossRef]
- Galli, F.; Archbold, D.D.; Pomper, K.W. Pawpaw fruit chilling injury and antioxidant protection. J. Am. Soc. Hortic. Sci. 2009, 134, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Long, L.E. Physiological and biochemical changes relating to postharvest splitting of sweet cherries affected by calcium application in hydrocooling water. Food Chem. 2015, 181, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Naser, F.; Rabiei, V.; Razavi, F.; Khademi, O. Effect of calcium lactate in combination with hot water treatment on the nutritional quality of persimmon fruit during cold storage. Sci. Hortic. 2018, 233, 114–123. [Google Scholar] [CrossRef]
- Gerasopoulos, D.; Drogoudi, P.D. Summer-pruning and preharvest calcium chloride sprays affect storability and low temperature breakdown incidence in kiwifruit. Postharvest Biol. Technol. 2005, 36, 303–308. [Google Scholar] [CrossRef]
- Sugar, D.; Basile, S.R. Orchard calcium and fungicide treatments mitigate effects of delayed postharvest fungicide applications for control of postharvest decay of pear fruit. Postharvest Biol. Technol. 2011, 60, 52–56. [Google Scholar] [CrossRef]
- Madani, B.; Mohamed, M.T.M.; Watkins, C.B.; Kadir, J.; Awang, Y.; Shojaei, T.R. Preharvest calcium chloride sprays affect ripening of Eksotika II’papaya fruits during cold storage. Sci. Hortic. 2014, 171, 6–13. [Google Scholar] [CrossRef]
- Fang, T.; Zhen, Q.; Liao, L.; Owiti, A.; Zhao, L.; Korban, S.S.; Han, Y. Variation of ascorbic acid concentration in fruits of cultivated and wild apples. Food Chem. 2017, 225, 132–137. [Google Scholar] [CrossRef]
- Lin, L.; Li, Q.P.; Wang, B.G.; Cao, J.K.; Jiang, W.B. Inhibition of core browning in ‘Yali’ pear fruit by post-harvest treatment with ascorbic acid. J. Hortic. Sci. Biotechnol. 2007, 82, 397–402. [Google Scholar] [CrossRef]
- Sogvar, O.B.; Koushesh Saba, M.; Emamifar, A. Aloe vera and ascorbic acid coatings maintain postharvest quality and reduce microbial load of strawberry fruit. Postharvest Biol. Technol. 2016, 114, 29–35. [Google Scholar] [CrossRef]
- Campos-Vargas, R.; Becerra, O.; Baeza-Yates, R.; Cambiazo, V.; González, M.; Meisel, L.; Orellana, A.; Retamales, J.; Silva, H.; Defilippi, B.G. Seasonal variation in the development of chilling injury in ‘O’Henry’ peaches. Sci. Hortic. 2006, 110, 79–83. [Google Scholar] [CrossRef]
- Gupta, A.S.; Webb, R.P.; Holaday, A.S.; Allen, R.D. Overexpression of Superoxide Dismutase Protects Plants from Oxidative Stress (Induction of Ascorbate Peroxidase in Superoxide Dismutase-Overexpressing Plants). Plant Physiol. 1993, 103, 1067–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elia, M.R.; Borraccino, G.; Dipierro, S. Soluble ascorbate peroxidase from potato tubers. Plant Sci. 1992, 85, 17–21. [Google Scholar] [CrossRef]
- Duan, X.; Liu, T.; Zhang, D.; Su, X.; Lin, H.; Jiang, Y. Effect of pure oxygen atmosphere on antioxidant enzyme and antioxidant activity of harvested litchi fruit during storage. Food Res. Int. 2011, 44, 1905–1911. [Google Scholar] [CrossRef]
- Ali, S.; Khan, A.S.; Malik, A.U.; Shahid, M. Effect of controlled atmosphere storage on pericarp browning, bioactive compounds and antioxidant enzymes of litchi fruits. Food Chem. 2016, 206, 18–29. [Google Scholar] [CrossRef]
- Rapisarda, P.; Bianco, M.L.; Pannuzzo, P.; Timpanaro, N. Effect of cold storage on vitamin C, phenolics and antioxidant activity of five orange genotypes [Citrus sinensis (L.) Osbeck]. Postharvest Biol. Technol. 2008, 49, 348–354. [Google Scholar] [CrossRef]
- Hou, Y.; Li, Z.; Zheng, Y.; Jin, P. Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis. Foods 2021, 10, 1662. [Google Scholar] [CrossRef]
- Niazi, Z.; Razavi, F.; Khademi, O.; Aghdam, M.S. Exogenous application of hydrogen sulfide and γ-aminobutyric acid alleviates chilling injury and preserves quality of persimmon fruit (Diospyros kaki, cv. Karaj) during cold storage. Sci. Hortic. 2021, 285, 110198. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Calcium: Transport and Physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef] [Green Version]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.-P.; Thuleau, P.; Mazars, C. Calcium Sensors as Key Hubs in Plant Responses to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium Signaling-Mediated Plant Response to Cold Stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, J.-W.; Zhou, B.; Li, G.-D.; Liu, Y.-F.; Xia, X.-L.; Xiao, Z.-G.; Fei, L.; Ji, S.-J. Calcium inhibited peel browning by regulating enzymes in membrane metabolism of ‘Nanguo’ pears during post-ripeness after refrigerated storage. Sci. Hortic. 2019, 244, 15–21. [Google Scholar] [CrossRef]
- Wei, D.; Zhao, X.-h. Calcium maintained higher quality and enhanced resistance against chilling stress by regulating enzymes in reactive oxygen and biofilm metabolism of Chinese winter jujube fruit. J. Food Biochem. 2020, 44, 13161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ma, M.; Ye, B.; Liu, L.; Ji, S. Calcium ion improves cold resistance of green peppers (Capsicum annuum L.) by regulating the activity of protective enzymes and membrane lipid composition. Sci. Hortic. 2021, 277, 109789. [Google Scholar] [CrossRef]
- Li, Z.; Wang, L.; Xie, B.; Hu, S.; Zheng, Y.; Jin, P. Effects of exogenous calcium and calcium chelant on cold tolerance of postharvest loquat fruit. Sci. Hortic. 2020, 269, 109391. [Google Scholar] [CrossRef]
- Wang, C.F.; Cheng, Z.M.; Li, Y. Effect of postharvest treatments on physiology and quality of litchi and their economics. Acta Hortic. 1996, 429, 503–507. [Google Scholar] [CrossRef]
- Shewfelt, R.L.; del Rosario, B.A. The role of lipid peroxidation in storage disorders of fresh fruits and vegetables. HortScience 2000, 35, 575–579. [Google Scholar] [CrossRef]
- Tang, T.; Huang, D.W.; Zhou, C.Q.; Li, X.; Xie, Q.J.; Liu, F.S. Molecular cloning and expression patterns of copper/zinc superoxide dismutase and manganese superoxide dismutase in Musca domestica. Gene 2012, 505, 211–220. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
A. A., L.; Ismail, H.; Kassem, H.S. Postharvest Treatment of ‘Florida Prince’ Peaches with a Calcium Nanoparticle–Ascorbic Acid Mixture during Cold Storage and Its Effect on Antioxidant Enzyme Activities. Horticulturae 2021, 7, 499. https://doi.org/10.3390/horticulturae7110499
A. A. L, Ismail H, Kassem HS. Postharvest Treatment of ‘Florida Prince’ Peaches with a Calcium Nanoparticle–Ascorbic Acid Mixture during Cold Storage and Its Effect on Antioxidant Enzyme Activities. Horticulturae. 2021; 7(11):499. https://doi.org/10.3390/horticulturae7110499
Chicago/Turabian StyleA. A., Lo’ay, Hamed Ismail, and Hazem S. Kassem. 2021. "Postharvest Treatment of ‘Florida Prince’ Peaches with a Calcium Nanoparticle–Ascorbic Acid Mixture during Cold Storage and Its Effect on Antioxidant Enzyme Activities" Horticulturae 7, no. 11: 499. https://doi.org/10.3390/horticulturae7110499