Author Contributions
Conceptualization, S.M. and S.K.; Data curation, S.S.; Formal analysis, S.S.; Funding acquisition, S.K.; Investigation, S.S. and D.B.; Methodology, S.S., S.M. and S.K.; Project administration, S.K.; Resources, S.K.; Supervision, S.M. and S.K.; Visualization, S.S. and A.S.; Writing—original draft, S.S.; Writing—review & editing, S.S., S.M., A.S. and S.K. All authors have read and agreed to the published version of the manuscript.
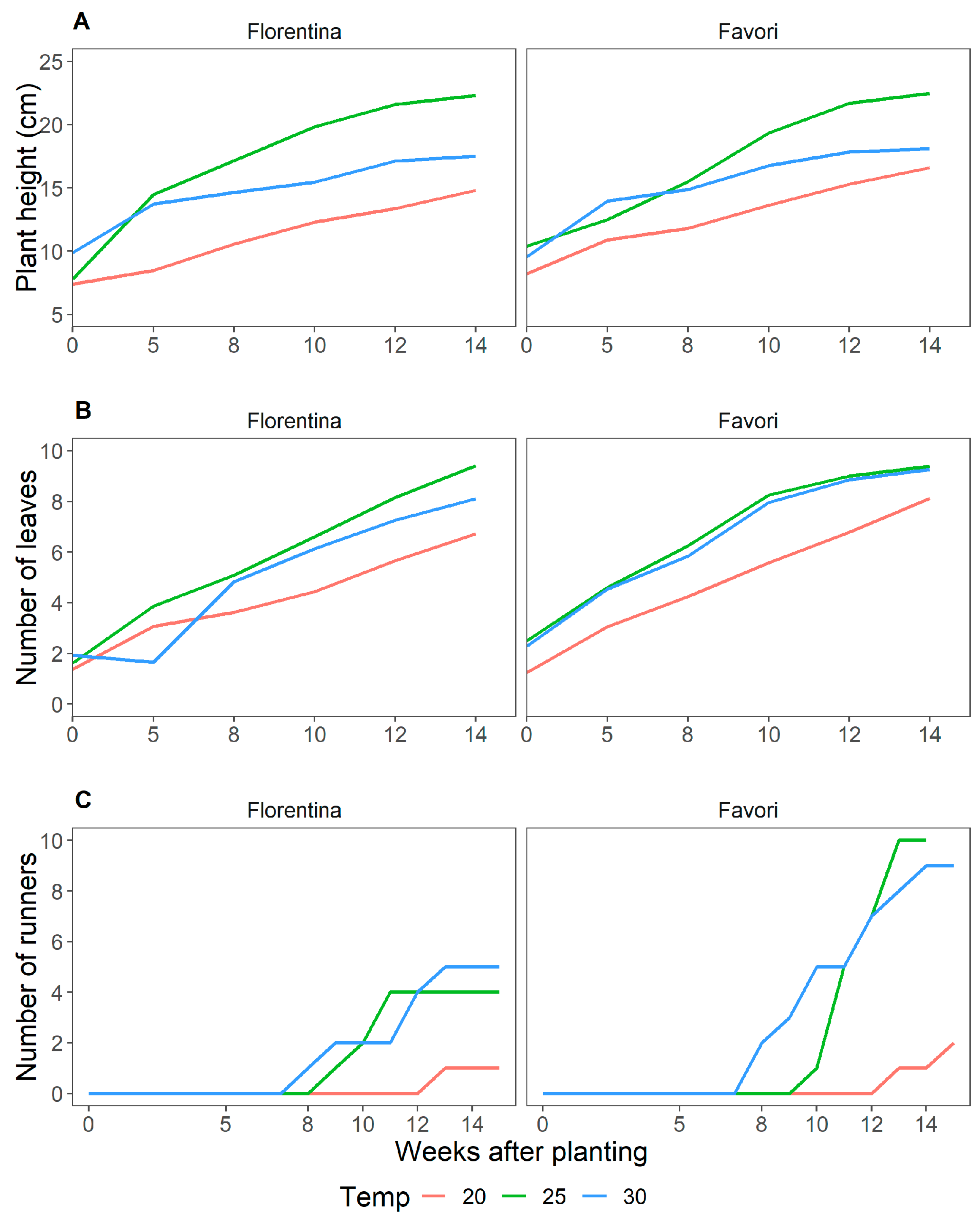
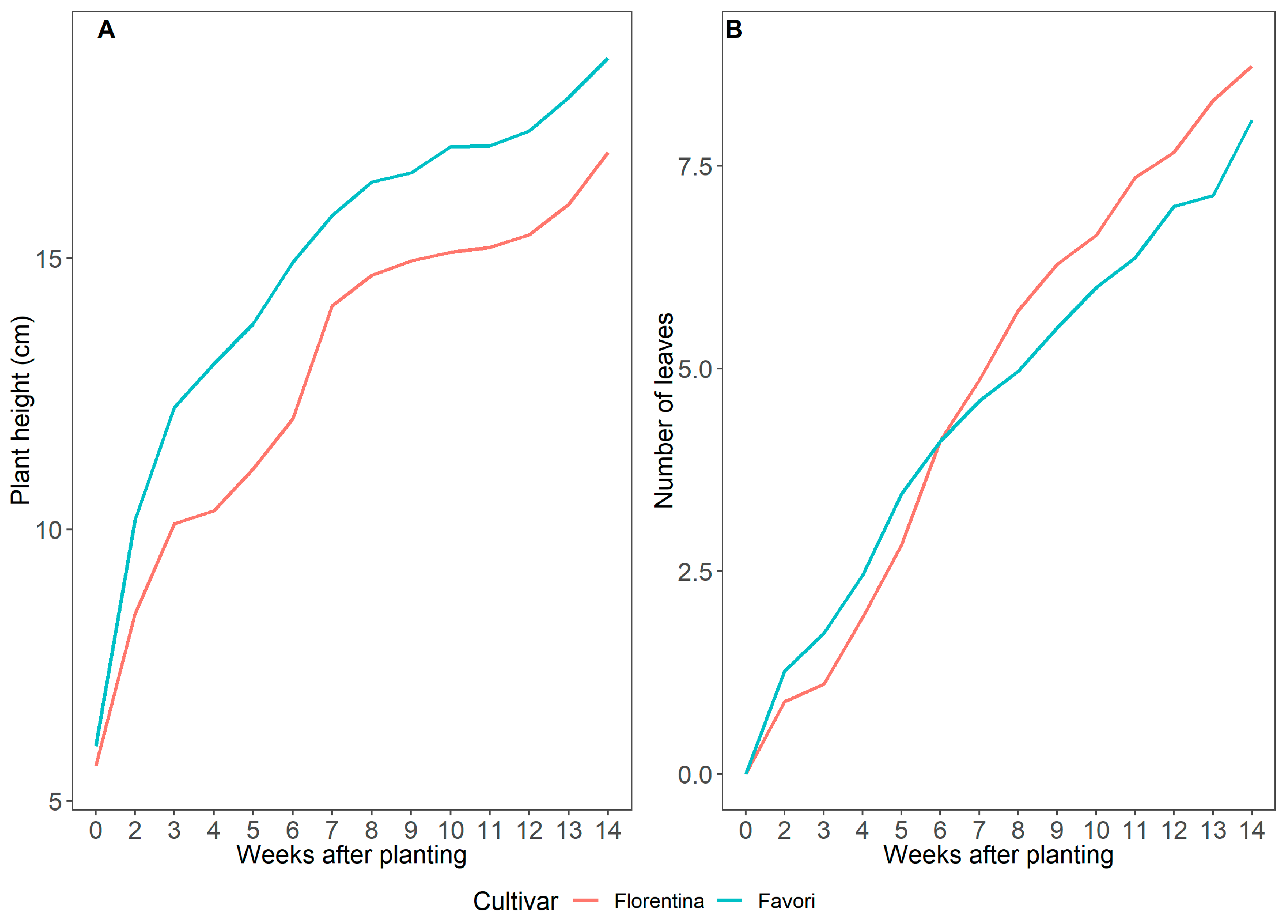
Figure 1.
Average plant height in cm (A); average number of leaves (B), starting at W0; and number of runners (C), starting at W8 for strawberry plants of cvs. ‘Florentina’ (N = 30–32) and ‘Favori’ (N = 20–34) during 14 weeks at ambient natural short day at 20 °C, 25 °C, and 30 °C (Expt. 1).
Figure 1.
Average plant height in cm (A); average number of leaves (B), starting at W0; and number of runners (C), starting at W8 for strawberry plants of cvs. ‘Florentina’ (N = 30–32) and ‘Favori’ (N = 20–34) during 14 weeks at ambient natural short day at 20 °C, 25 °C, and 30 °C (Expt. 1).
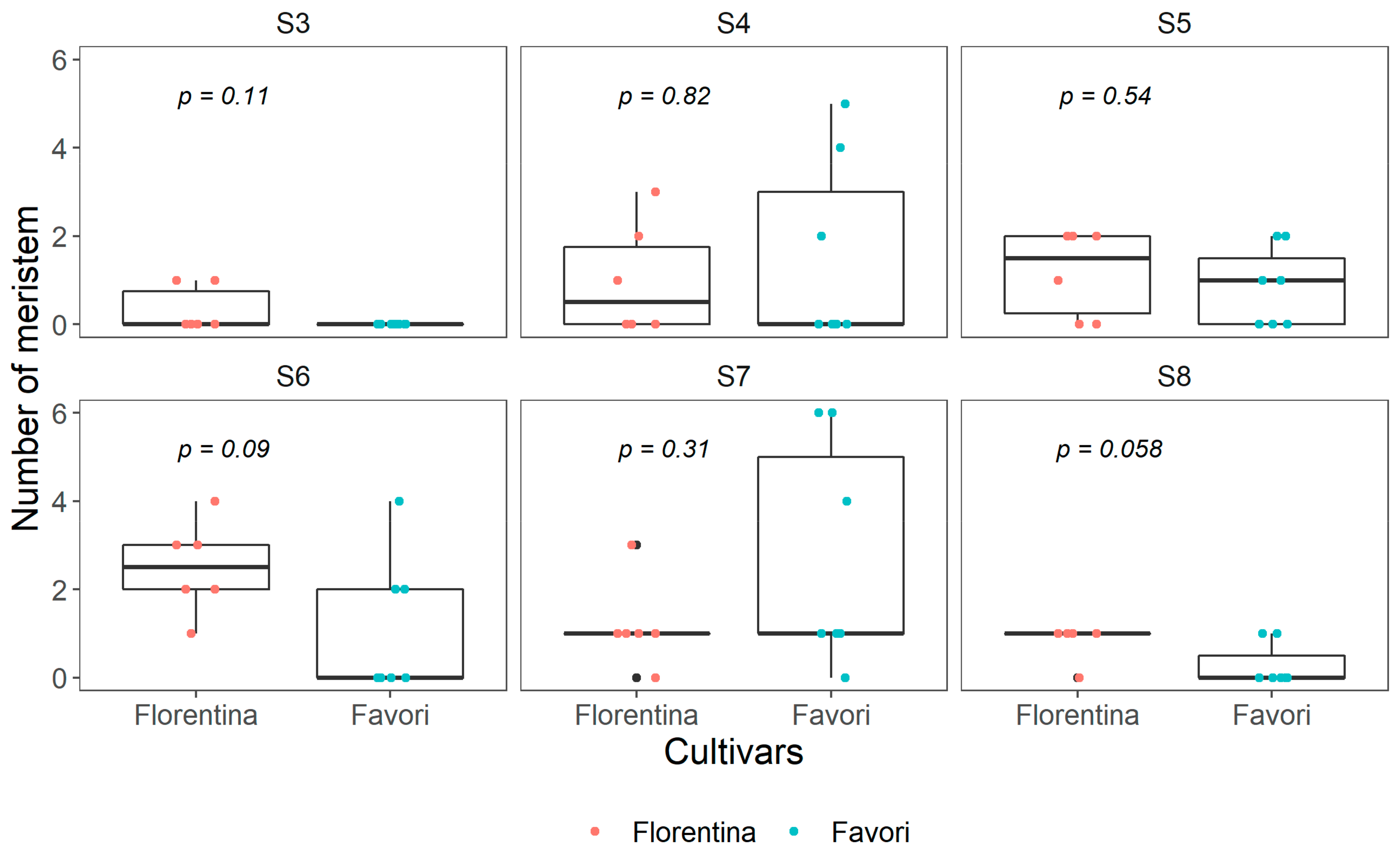
Figure 2.
Boxplot of number of meristems at each developmental stage (S3–S8) in ‘Florentina’ and ‘Favori’ strawberry plants at start of experiment (W0), N = 7, where any bud below S4 was pooled as vegetative and categorized as S3. The p-value is adjusted using pairwise Kruskal−Wallis test.
Figure 2.
Boxplot of number of meristems at each developmental stage (S3–S8) in ‘Florentina’ and ‘Favori’ strawberry plants at start of experiment (W0), N = 7, where any bud below S4 was pooled as vegetative and categorized as S3. The p-value is adjusted using pairwise Kruskal−Wallis test.
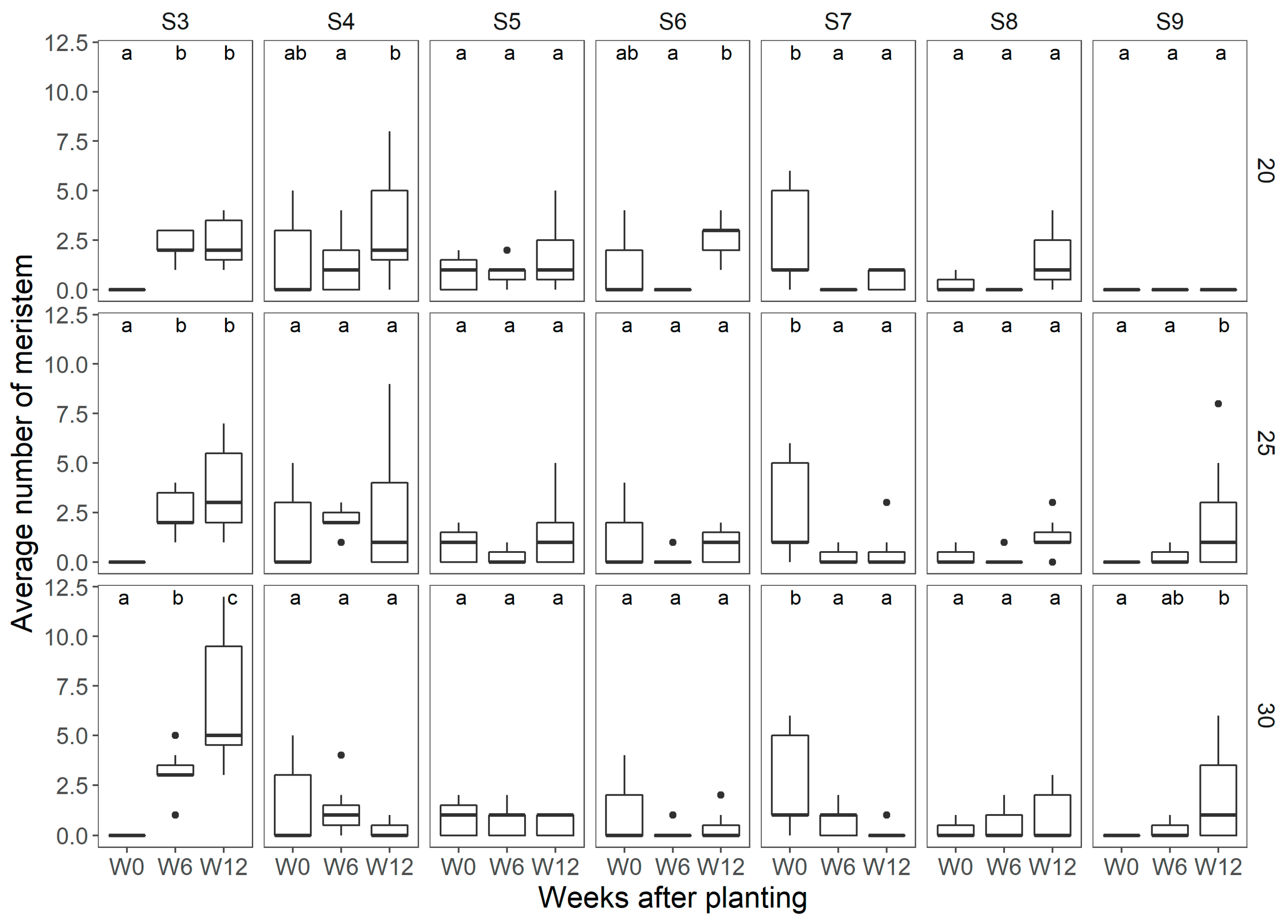
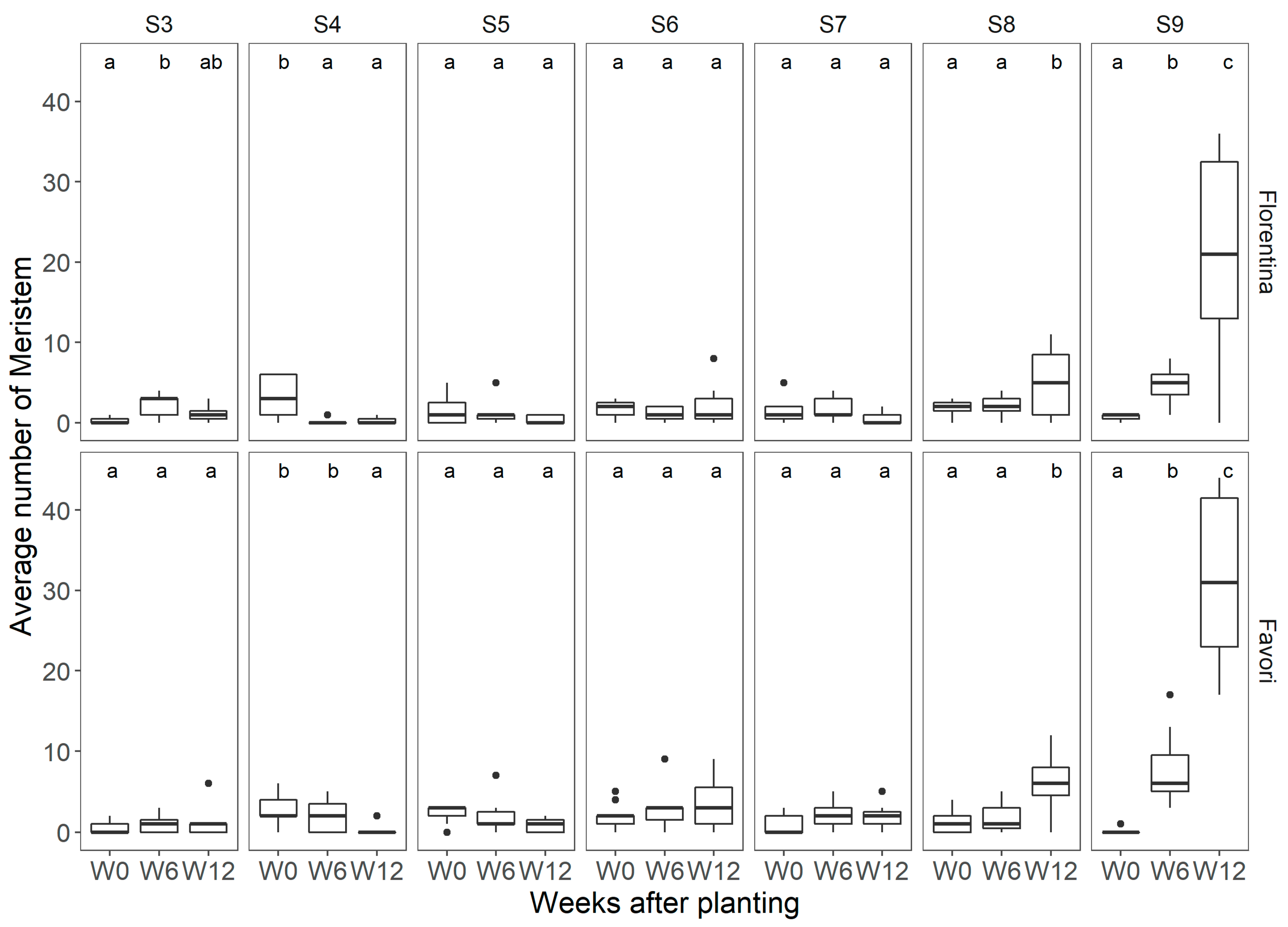
Figure 3.
Average number of meristems observed per temperature treatment at each of the flower mapping occasions in ‘Florentina’, where W0 = week 0, W6 = week 6, and W12 = week 12 after start of treatment, at 20 °C, 25 °C, and 30 °C, N = 7. The different letters above the boxplots in each box denote differences among means between the flower mapping occasions per temperature treatment, and similar lowercase letters indicate non-significant difference by the Holm−Sidak post-hoc test at 5% of error probability (p < 0.05).
Figure 3.
Average number of meristems observed per temperature treatment at each of the flower mapping occasions in ‘Florentina’, where W0 = week 0, W6 = week 6, and W12 = week 12 after start of treatment, at 20 °C, 25 °C, and 30 °C, N = 7. The different letters above the boxplots in each box denote differences among means between the flower mapping occasions per temperature treatment, and similar lowercase letters indicate non-significant difference by the Holm−Sidak post-hoc test at 5% of error probability (p < 0.05).
Figure 4.
Average number of meristems observed per temperature treatment at each of the flower mapping occasions for ‘Favori’, where W0 = week 0, W6 = week 6, and W12 = week 12 after start of treatment, at 20 °C 25 °C, and 30 °C, N = 7. The different letters above the boxplots in each box denote differences among means between the flower mapping occasions per temperature treatment and similar lowercase letters indicate non-significant difference by the Holm−Sidak post-hoc at 5% of error probability (p < 0.05).
Figure 4.
Average number of meristems observed per temperature treatment at each of the flower mapping occasions for ‘Favori’, where W0 = week 0, W6 = week 6, and W12 = week 12 after start of treatment, at 20 °C 25 °C, and 30 °C, N = 7. The different letters above the boxplots in each box denote differences among means between the flower mapping occasions per temperature treatment and similar lowercase letters indicate non-significant difference by the Holm−Sidak post-hoc at 5% of error probability (p < 0.05).
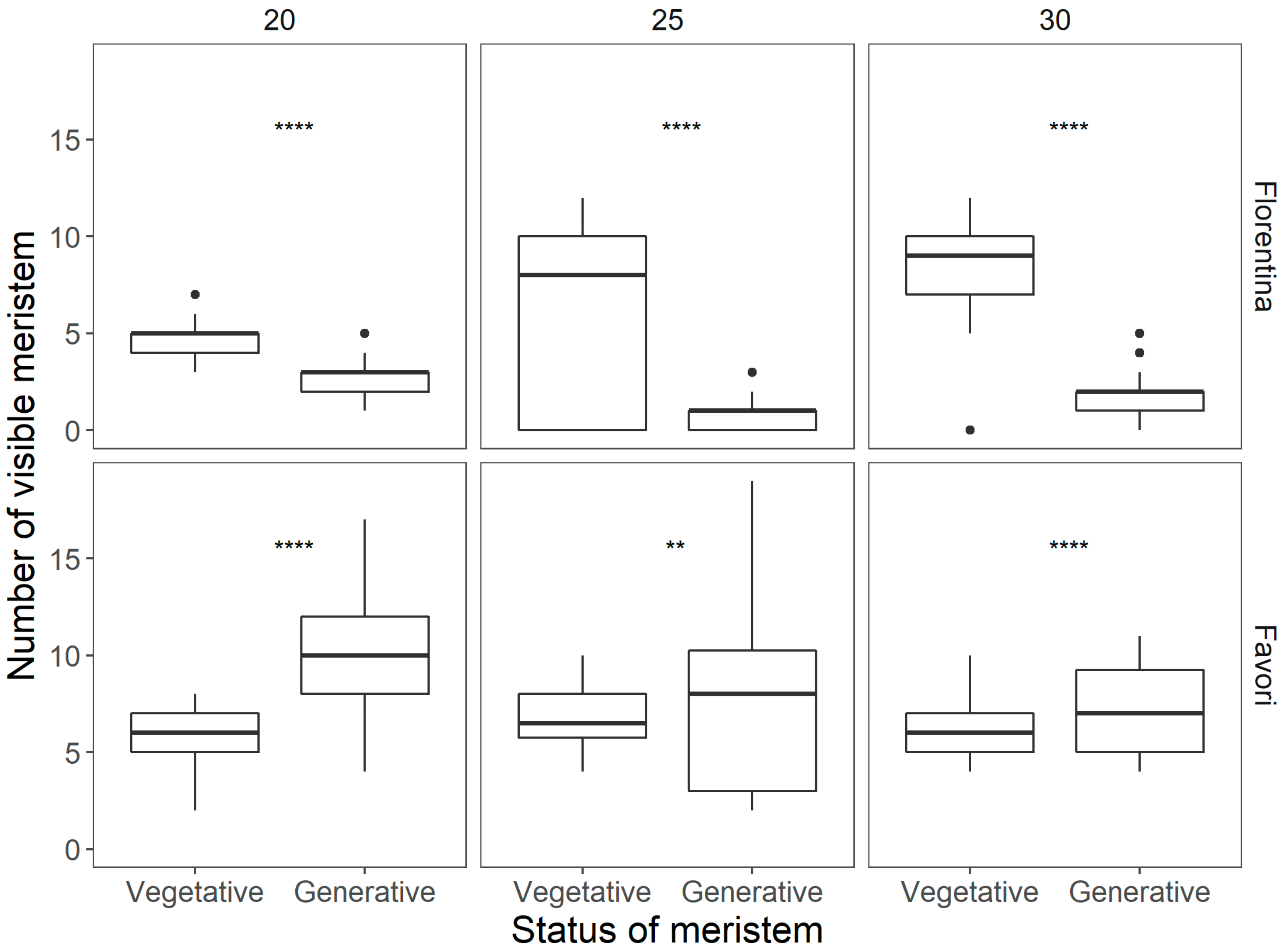
Figure 5.
Differences in the status of the meristem at the end of 15 weeks of temperature treatments for each cultivar Statistically significant differences between the vegetative and generative stages were calculated by ANOVA and are indicated by asterisks (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001).
Figure 5.
Differences in the status of the meristem at the end of 15 weeks of temperature treatments for each cultivar Statistically significant differences between the vegetative and generative stages were calculated by ANOVA and are indicated by asterisks (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001).
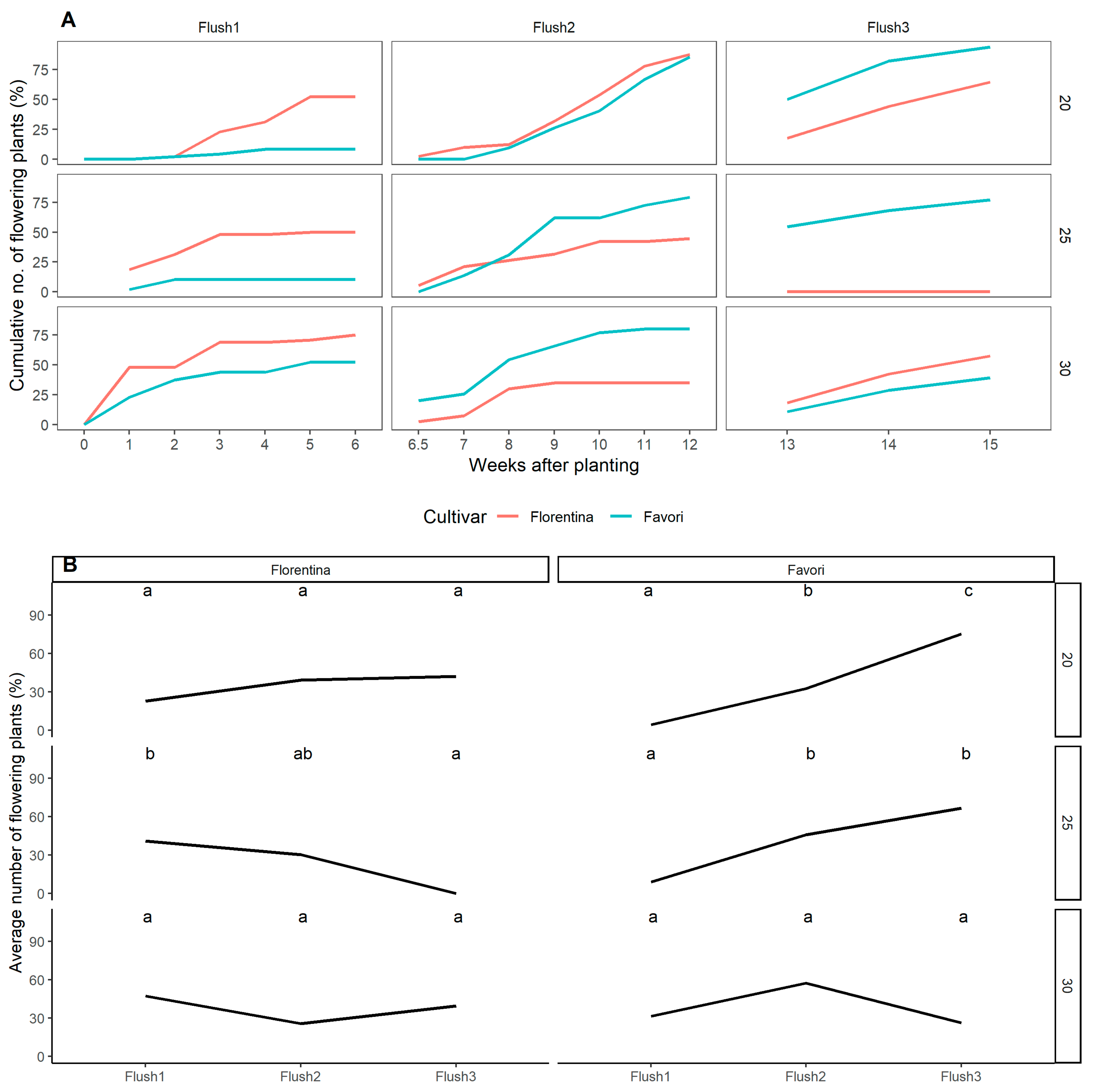
Figure 6.
Cumulative number of flowering plants (%), recorded at anthesis, at each flush during the three temperature treatments for ‘Florentina’ and ‘Favori’ (A). The data was divided into three flushes, where flush 1 = W0–W6, flush 2 = W7–W12, and flush 3 = W13–W15, correlating with the flower mapping data. The number of plants that flushed were compared for each cultivar at each temperature treatment using pairwise comparison and letters were calculated using Tukey’s post-hoc method for multiple comparison. Similar letters indicate no significance at p = 0.05 (B).
Figure 6.
Cumulative number of flowering plants (%), recorded at anthesis, at each flush during the three temperature treatments for ‘Florentina’ and ‘Favori’ (A). The data was divided into three flushes, where flush 1 = W0–W6, flush 2 = W7–W12, and flush 3 = W13–W15, correlating with the flower mapping data. The number of plants that flushed were compared for each cultivar at each temperature treatment using pairwise comparison and letters were calculated using Tukey’s post-hoc method for multiple comparison. Similar letters indicate no significance at p = 0.05 (B).
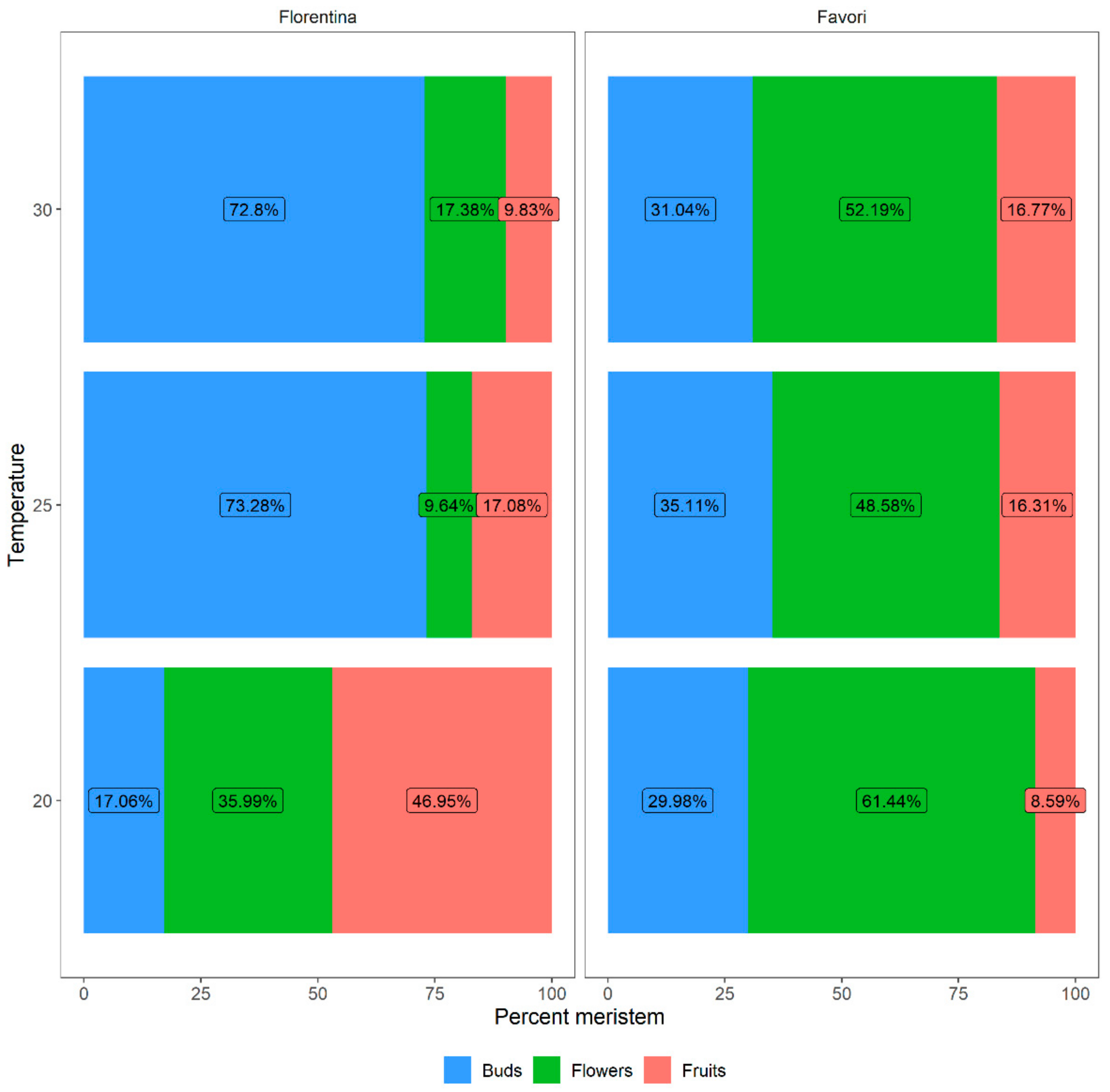
Figure 7.
The percentages of meristems that differentiated into flowers and fruits for plants of the strawberry cultivars ‘Florentina’ and ‘Favori’ treated at three temperatures (20 °C, 25 °C, and 30 °C). The buds that were below S9 (fully differentiated flower) were compiled as “Buds”.
Figure 7.
The percentages of meristems that differentiated into flowers and fruits for plants of the strawberry cultivars ‘Florentina’ and ‘Favori’ treated at three temperatures (20 °C, 25 °C, and 30 °C). The buds that were below S9 (fully differentiated flower) were compiled as “Buds”.
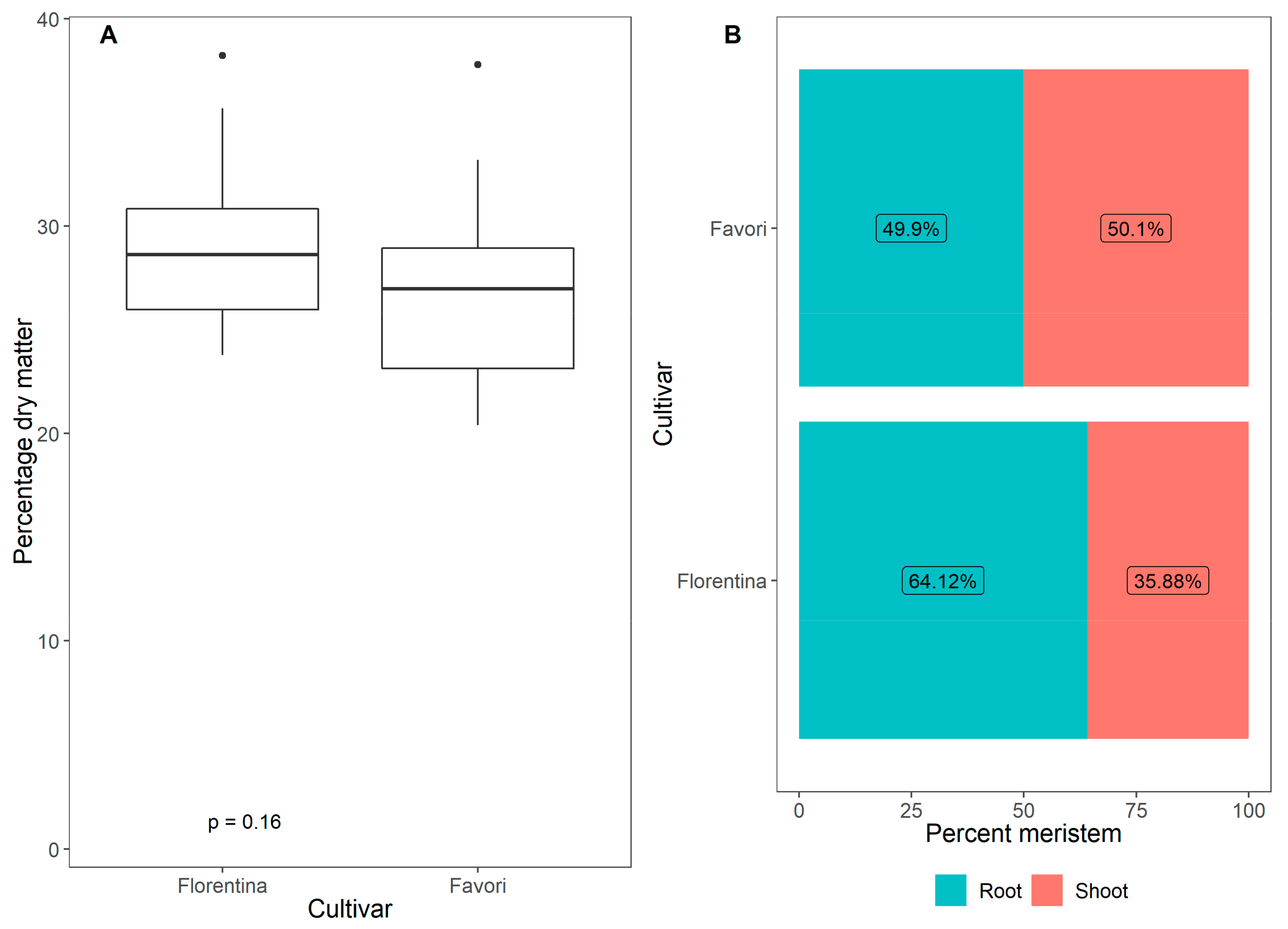
Figure 8.
Average plant height (A) and total number of leaves (B) for ‘Florentina’ and ‘Favori’ strawberry plants grown at ambient long day at 20 °C for 14 weeks (N = 29–32).
Figure 8.
Average plant height (A) and total number of leaves (B) for ‘Florentina’ and ‘Favori’ strawberry plants grown at ambient long day at 20 °C for 14 weeks (N = 29–32).
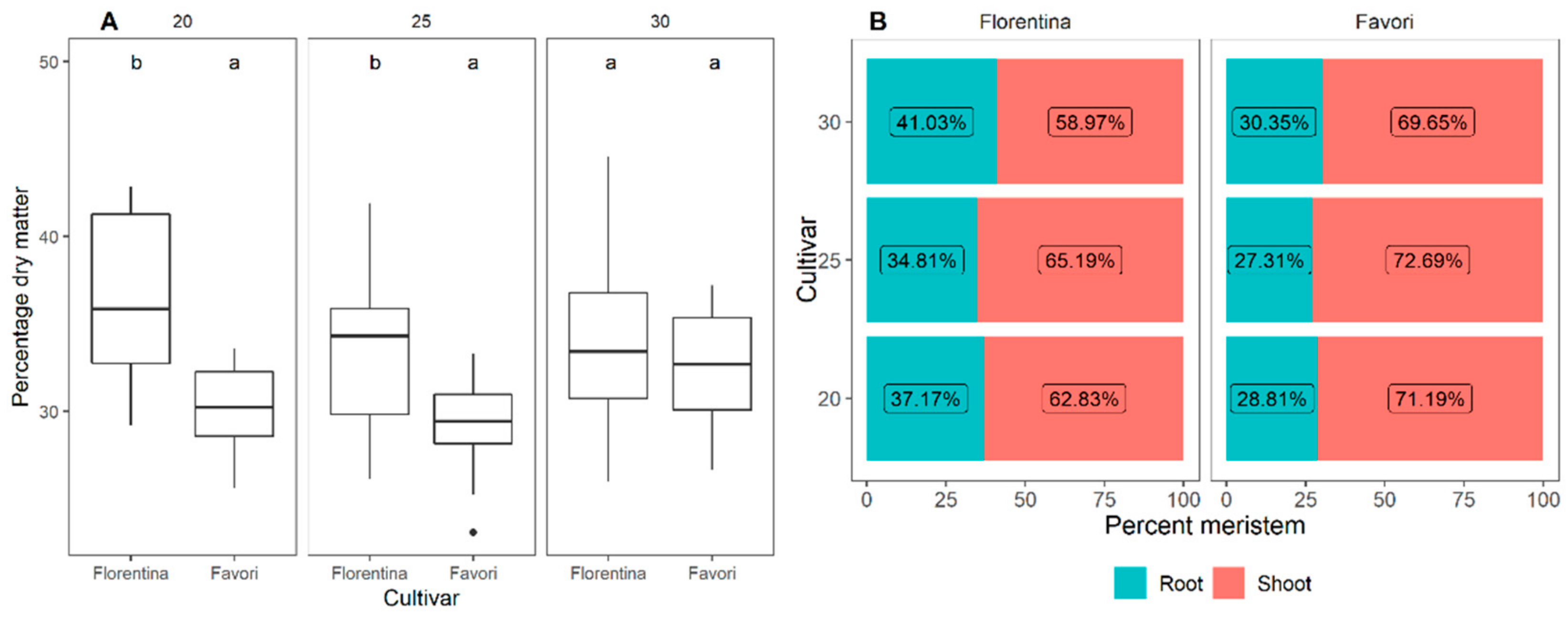
Figure 9.
Boxplots of the meristem development stages (S3–S9) at three timepoints (W0, W6, and W12) for plants of the strawberry cultivar ‘Florentina’ and ‘Favori’ grown in a greenhouse at ambient long photoperiod at 20 °C for 14 weeks, where N = 7. Pairwise comparison between the week, where the different letters above the boxplots represent statistical differences, while similar letters indicate no significant difference between cultivars by the Tukey test at 5% of error probability (p < 0.05).
Figure 9.
Boxplots of the meristem development stages (S3–S9) at three timepoints (W0, W6, and W12) for plants of the strawberry cultivar ‘Florentina’ and ‘Favori’ grown in a greenhouse at ambient long photoperiod at 20 °C for 14 weeks, where N = 7. Pairwise comparison between the week, where the different letters above the boxplots represent statistical differences, while similar letters indicate no significant difference between cultivars by the Tukey test at 5% of error probability (p < 0.05).
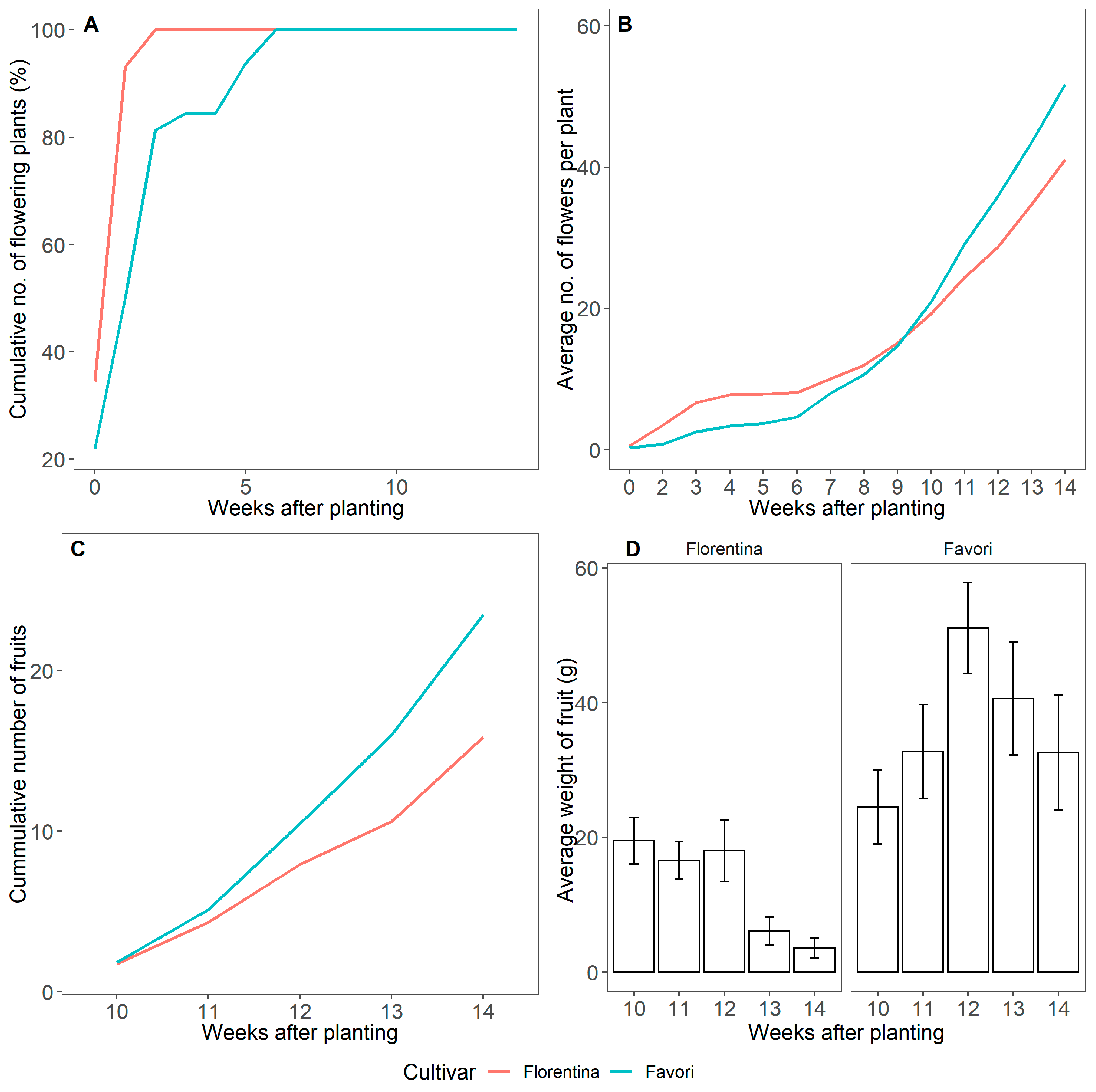
Figure 10.
Cumulative number of plants that flowered (%) (A), cumulative number of flowers per plant (B), cumulative number of fruits collected (C), and average weight of the total number of fruits collected on the given week with the error bars representing the standard error (D) over the course of 14 weeks at 20 °C under LD condition for ‘Florentina’ and ‘Favori’.
Figure 10.
Cumulative number of plants that flowered (%) (A), cumulative number of flowers per plant (B), cumulative number of fruits collected (C), and average weight of the total number of fruits collected on the given week with the error bars representing the standard error (D) over the course of 14 weeks at 20 °C under LD condition for ‘Florentina’ and ‘Favori’.
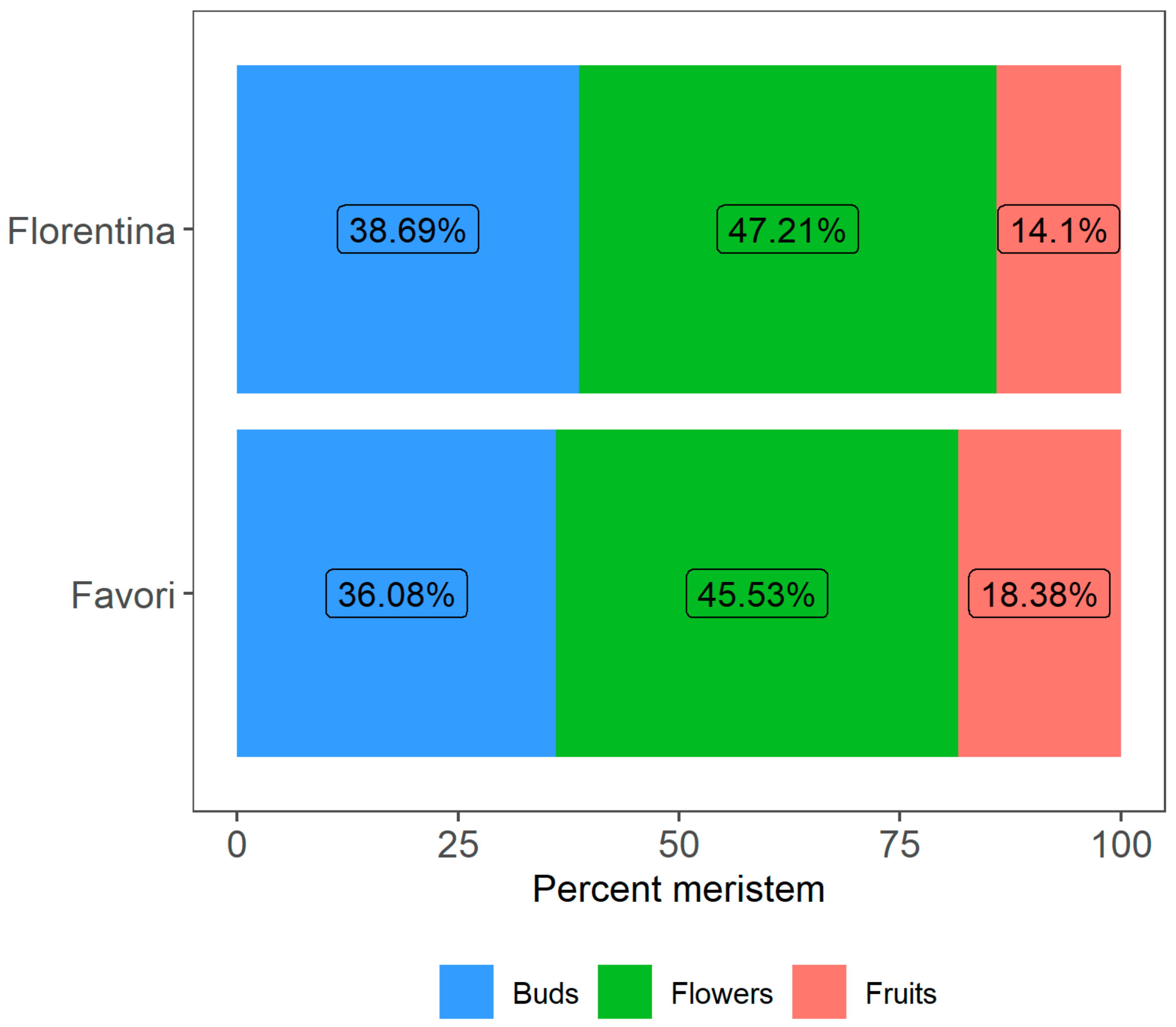
Figure 11.
Percentage of meristems that differentiated into flowers and fruits for plants of the strawberry cultivars ‘Florentina’ and ‘Favori’ after 14 weeks at 20 °C under LD conditions. The buds that were below S9 (fully differentiated flower) were compiled as buds.
Figure 11.
Percentage of meristems that differentiated into flowers and fruits for plants of the strawberry cultivars ‘Florentina’ and ‘Favori’ after 14 weeks at 20 °C under LD conditions. The buds that were below S9 (fully differentiated flower) were compiled as buds.
Table 1.
Experimental set up with the two everbearing strawberry cultivars ‘Florentina’and ‘Favori’, conducted in a greenhouse at Alnarp, Sweden, under ambient short day (SD), long day (LD) and temperatures conditions as indicated.
Table 1.
Experimental set up with the two everbearing strawberry cultivars ‘Florentina’and ‘Favori’, conducted in a greenhouse at Alnarp, Sweden, under ambient short day (SD), long day (LD) and temperatures conditions as indicated.
| | SD | LD |
|---|
| Temperature (day/night) | 20/17 °C, 25/24 °C and 30/27 °C | 20/20 °C |
| Day length | 7–10 h | 16–18 h |
| Duration | 15 weeks | 14 weeks |
| Substrate | Peat (WeibullHorto- Sweden) |
| Fertilization | Kristalon superba Red (NPK 12-5-30; EC = 1.3 mS/cm) and Calcinit (15% N, 14.4% NO3−, 1.1% Al, 19% Ca; EC = 1.1 mS/cm) |
| Plants per pot | 4 plants per 11 L pot |
| Number of replicates | 12 pots per cultivar per temperature treatment |
Table 2.
Description of floral developmental stages (S3 to S9) for strawberry plants assessed under a stereo light microscope. Stages below S4 were regarded as vegetative, while stages from 4 to 9 were regarded as reproductive.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



















