
Development of a Cost-Effective Process for the Heterologous Production of SARS-CoV-2 Spike Receptor Binding Domain Using Pichia pastoris in Stirred-Tank Bioreactor
 ,
,  , , , ,
, , , , 
Abstract
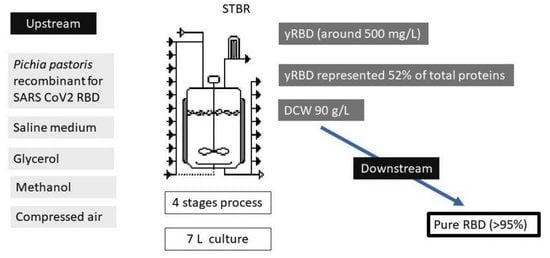
:
1. Introduction
2. Methods
2.1. Plasmid and Strain
2.2. Determination of Dry Cell Weight
2.3. Quantification of Total Proteins and RBD
2.4. Medium Composition for Flask Cultures and Bioreactor Fermentations
2.5. Inoculum Preparation
2.6. Cultivation in Erlenmeyer Flask
2.7. Fermentations in Stirred-Tank Bioreactor
2.8. Purification of Recombinant RBD and Quality Control
2.9. Statistical Analysis
3. Results
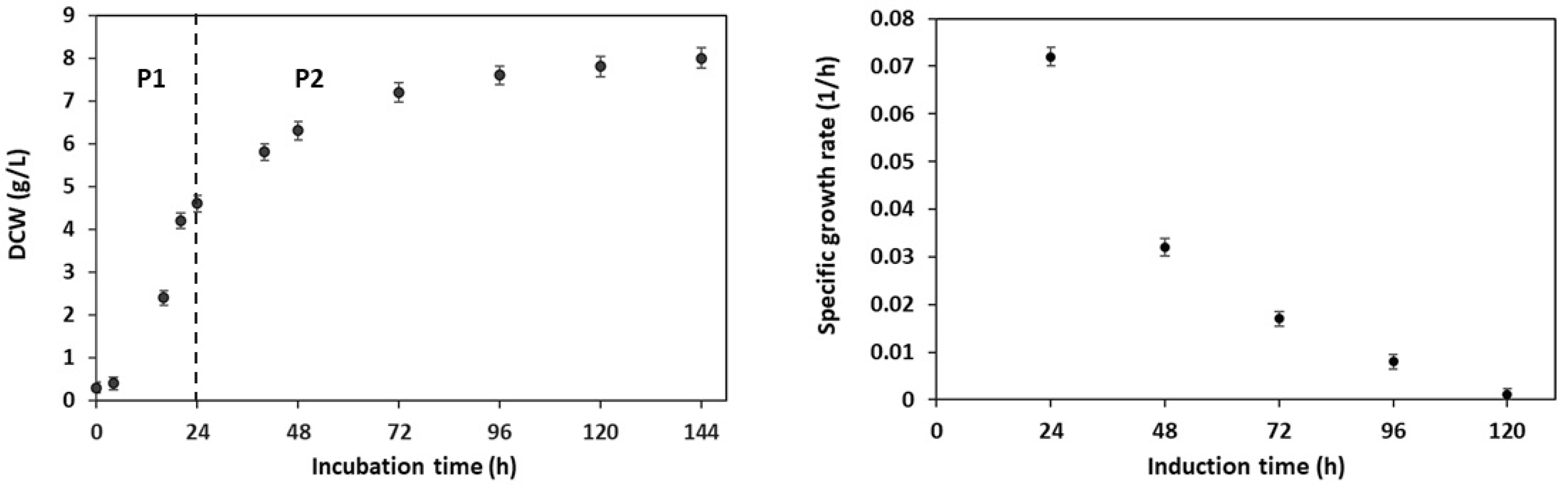
3.1. Growth Kinetics and RBD Expression at Flask Level
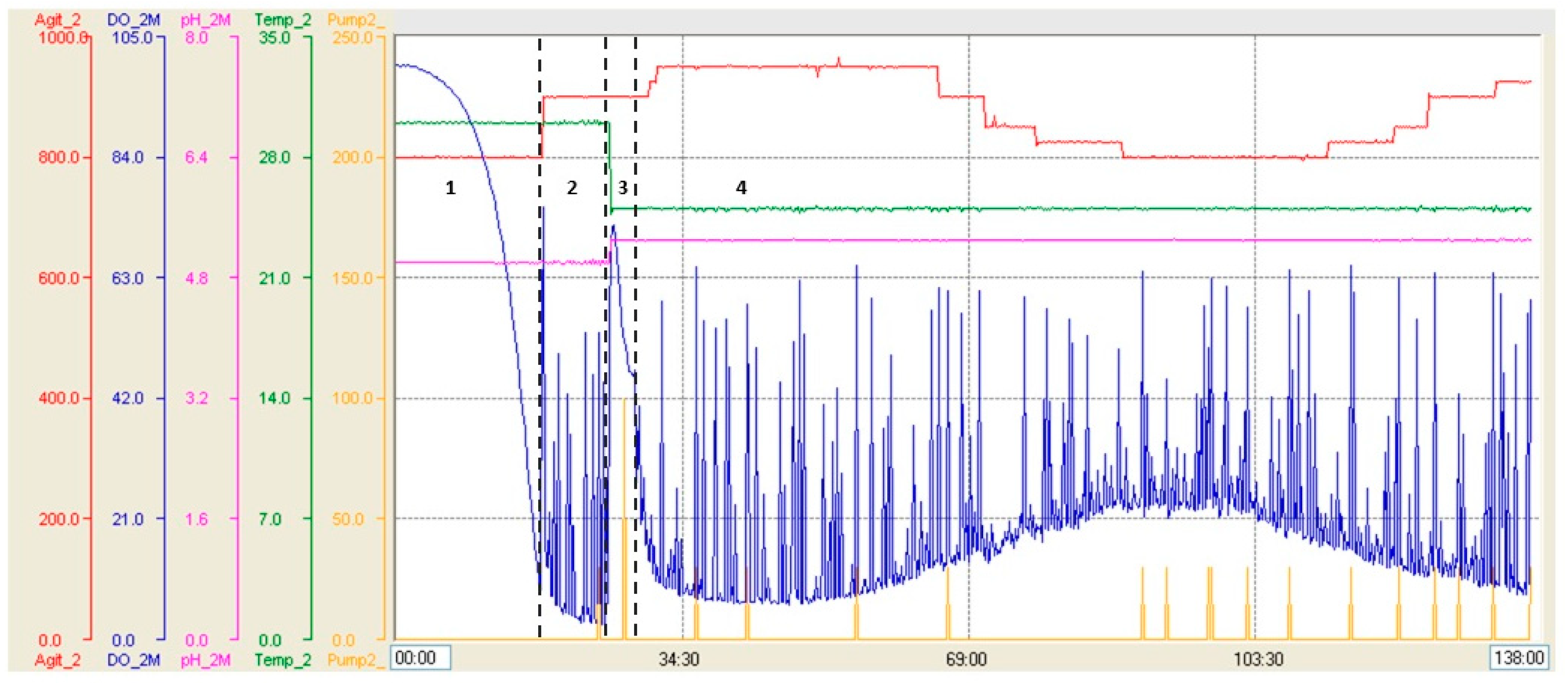
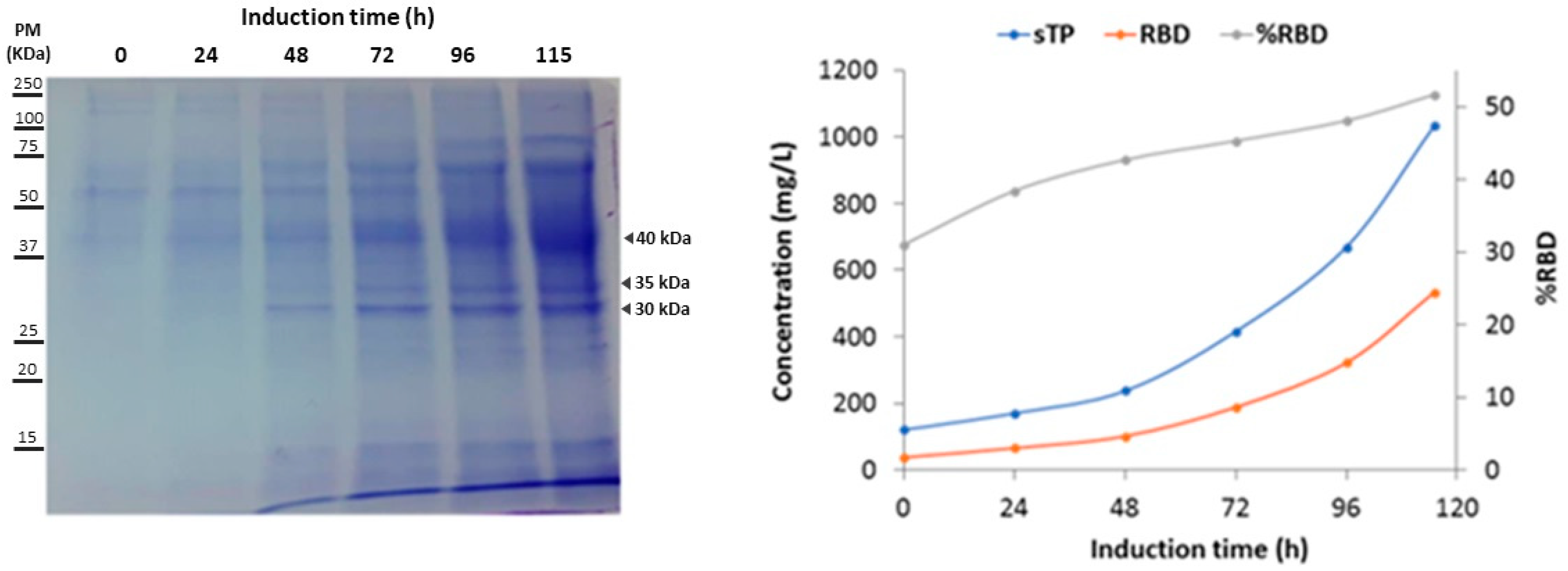
3.2. Production of Recombinant RBD in 7 L and 14 L Stirred Bioreactors
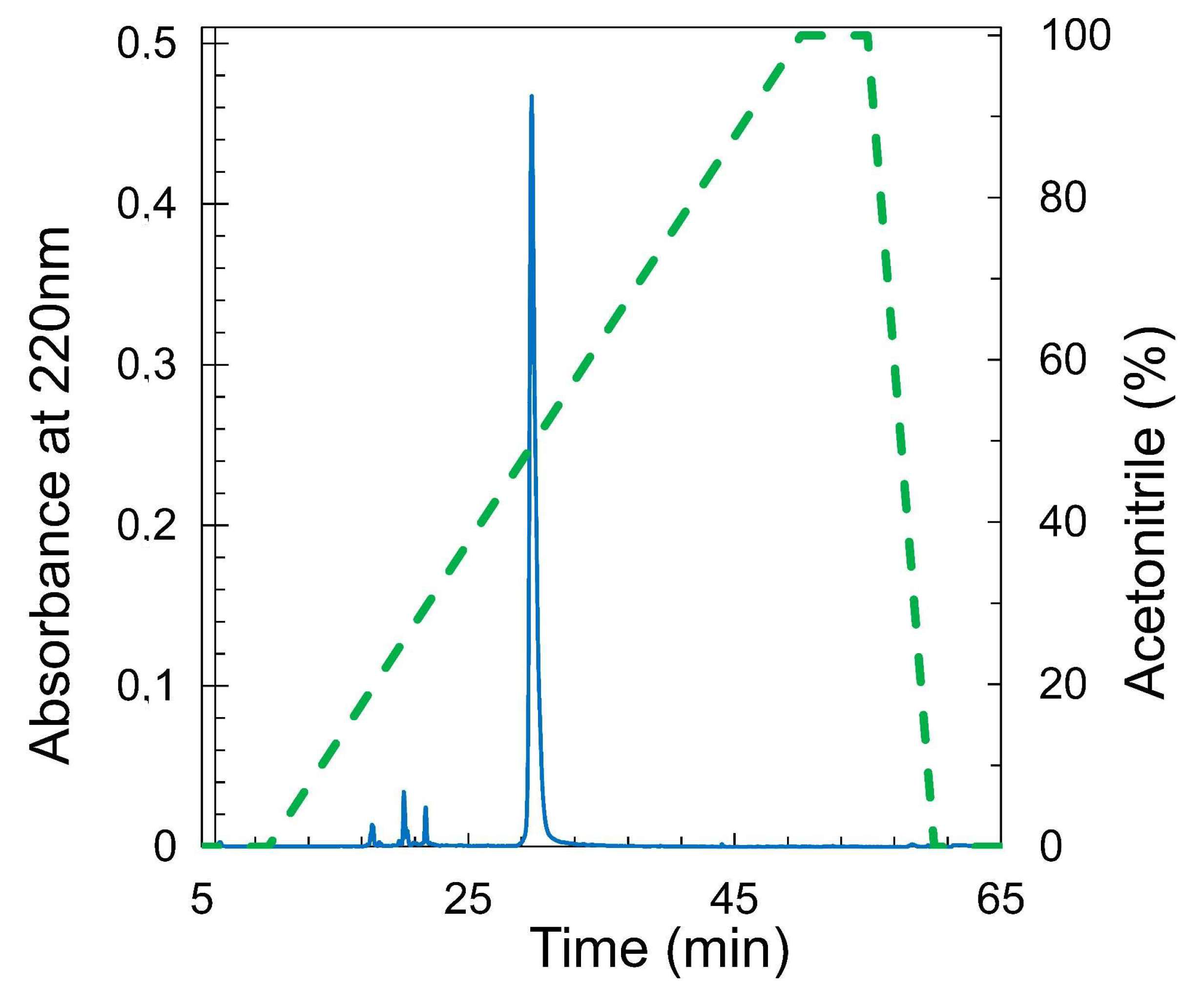
3.3. Purification and Analysis of RBD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dennehy, J.J.; Gupta, R.K.; Hanage, W.P.; Johnson, M.C.; Peacock, T.P. Where Is the next SARS-CoV-2 Variant of Concern? Lancet 2022, 399, 1938–1939. [Google Scholar] [CrossRef] [PubMed]
- Forster, P.; Forster, L.; Renfrew, C.; Forster, M. Phylogenetic Network Analysis of SARS-CoV-2 Genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 9241–9243. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-M.; Mannan, R.; Xiao, L.; Abdulfatah, E.; Qiao, Y.; Farver, C.; Myers, J.L.; Zelenka-Wang, S.; McMurry, L.; Su, F.; et al. Characterization of SARS-CoV-2 and Host Entry Factors Distribution in a COVID-19 Autopsy Series. Commun. Med. 2021, 1, 24. [Google Scholar] [CrossRef] [PubMed]
- Arimori, T.; Takagi, J. Structure of SARS-CoV-2 Spike Receptor-Binding Domain Complexed with High Affinity ACE2 Mutant 3N39 2020. Nat. Commun. 2021, 12, 3802. [Google Scholar]
- Xu, Z.P.; Liu, K.F.; Han, P.; Qi, J.X. Structure of SARS-CoV-2 Spike Receptor-Binding Domain Complexed with Its Receptor Equine ACE2 2022. Structure 2022, 30, 1432–1442.e4. [Google Scholar]
- Wang, X.; Lan, J.; Ge, J.; Yu, J.; Shan, S. Crystal Structure of SARS-CoV-2 Spike Receptor-Binding Domain Bound with ACE2 Structure summary MMDB-NCBI 2020. Available online: https://www.ncbi.nlm.nih.gov/Structure/pdb/6M0J (accessed on 8 January 2022).
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, A.; Mohammad, T.; Anwar, S.; Amani, S.; Khan, M.S.; Husain, F.M.; Rehman, M.T.; Islam, A.; Hassan, M.I. Potential Drug Targets of SARS-CoV-2: From Genomics to Therapeutics. Int. J. Biol. Macromol. 2021, 177, 1–9. [Google Scholar] [CrossRef]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human Neutralizing Antibodies Elicited by SARS-CoV-2 Infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.-W.; Sahi, V.; Figueroa, A.; et al. Potent Neutralizing Antibodies against Multiple Epitopes on SARS-CoV-2 Spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef]
- Fujita, R.; Hino, M.; Ebihara, T.; Nagasato, T.; Masuda, A.; Lee, J.M.; Fujii, T.; Mon, H.; Kakino, K.; Nagai, R.; et al. Efficient Production of Recombinant SARS-CoV-2 Spike Protein Using the Baculovirus-Silkworm System. Biochem. Biophys. Res. Commun. 2020, 529, 257–262. [Google Scholar] [CrossRef]
- Argentinian AntiCovid Consortium. Covalent Coupling of Spike’s Receptor Binding Domain to a Multimeric Carrier Produces a High Immune Response against SARS-CoV-2. Sci. Rep. 2022, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Argentinian AntiCovid Consortium. Structural and Functional Comparison of SARS-CoV-2-Spike Receptor Binding Domain Produced in Pichia Pastoris and Mammalian Cells. Sci. Rep. 2020, 10, 21779. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.; Mc Callum, G.J.; Sabljic, A.V.; Marfía, J.I.; Bombicino, S.S.; Trabucchi, A.; Iacono, R.F.; Birenbaum, J.M.; Vázquez, S.C.; Minoia, J.M.; et al. Rapid and Cost-Effective Process Based on Insect Larvae for Scale-up Production of SARS-CoV-2 Spike Protein for Serological COVID-19 Testing. Biotechnol. Bioeng. 2021, 118, 4129–4137. [Google Scholar] [CrossRef]
- Li, T.; Zheng, Q.; Yu, H.; Wu, D.; Xue, W.; Xiong, H.; Huang, X.; Nie, M.; Yue, M.; Rong, R.; et al. SARS-CoV-2 Spike Produced in Insect Cells Elicits High Neutralization Titres in Non-Human Primates. Emerg. Microbes Infect. 2020, 9, 2076–2090. [Google Scholar] [CrossRef]
- Shajahan, A.; Supekar, N.T.; Gleinich, A.S.; Azadi, P. Deducing the N- and O-Glycosylation Profile of the Spike Protein of Novel Coronavirus SARS-CoV-2. Glycobiology 2020, 30, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Azad, T.; Singaravelu, R.; Taha, Z.; Jamieson, T.R.; Boulton, S.; Crupi, M.J.F.; Martin, N.T.; Brown, E.E.F.; Poutou, J.; Ghahremani, M.; et al. Nanoluciferase Complementation-Based Bioreporter Reveals the Importance of N-Linked Glycosylation of SARS-CoV-2 S for Viral Entry. Mol. Ther. 2021, 29, 1984–2000. [Google Scholar] [CrossRef]
- Jenkins, N.; Curling, E.M. Glycosylation of Recombinant Proteins: Problems and Prospects. Enzym. Microb. Technol. 1994, 16, 354–364. [Google Scholar] [CrossRef]
- Brooks, S.A. Appropriate Glycosylation of Recombinant Proteins for Human Use. Mol. Biotechnol. 2004, 28, 241–255. [Google Scholar] [CrossRef]
- Lingg, N.; Zhang, P.; Song, Z.; Bardor, M. The Sweet Tooth of Biopharmaceuticals: Importance of Recombinant Protein Glycosylation Analysis. Biotechnol. J. 2012, 7, 1462–1472. [Google Scholar] [CrossRef]
- Goh, J.B.; Ng, S.K. Impact of Host Cell Line Choice on Glycan Profile. Crit. Rev. Biotechnol. 2018, 38, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Casalino, L.; Gaieb, Z.; Goldsmith, J.A.; Hjorth, C.K.; Dommer, A.C.; Harbison, A.M.; Fogarty, C.A.; Barros, E.P.; Taylor, B.C.; McLellan, J.S.; et al. Beyond Shielding: The Roles of Glycans in the SARS-CoV-2 Spike Protein. ACS Cent. Sci. 2020, 6, 1722–1734. [Google Scholar] [CrossRef] [PubMed]
- Vieira Gomes, A.M.; Souza Carmo, T.; Silva Carvalho, L.; Mendonça Bahia, F.; Parachin, N.S. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernauer, L.; Radkohl, A.; Lehmayer, L.G.K.; Emmerstorfer-Augustin, A. Komagataella Phaffii as Emerging Model Organism in Fundamental Research. Front. Microbiol. 2020, 11, 607028. [Google Scholar] [CrossRef]
- Ata, Ö.; Ergün, B.G.; Fickers, P.; Heistinger, L.; Mattanovich, D.; Rebnegger, C.; Gasser, B. What Makes Komagataella Phaffii Non-Conventional? FEMS Yeast Res. 2021, 21, foab059. [Google Scholar] [CrossRef]
- Wegner, G.H. Emerging Applications of the Methylotrophic Yeasts. FEMS Microbiol. Rev. 1990, 7, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.; Hearn, M.T.W. Expression of Heterologous Proteins in Pichia Pastoris: A Useful Experimental Tool in Protein Engineering and Production. J. Mol. Recognit. 2005, 18, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Ergün, B.G.; Berrios, J.; Binay, B.; Fickers, P. Recombinant Protein Production in Pichia Pastoris: From Transcriptionally Redesigned Strains to Bioprocess Optimization and Metabolic Modelling. FEMS Yeast Res. 2021, 21, foab057. [Google Scholar] [CrossRef] [PubMed]
- Hartner, F.S.; Glieder, A. Regulation of Methanol Utilisation Pathway Genes in Yeasts. Microb. Cell Fact. 2006, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Hartner, F.S.; Ruth, C.; Langenegger, D.; Johnson, S.N.; Hyka, P.; Lin-Cereghino, G.P.; Lin-Cereghino, J.; Kovar, K.; Cregg, J.M.; Glieder, A. Promoter Library Designed for Fine-Tuned Gene Expression in Pichia Pastoris. Nucleic Acids Res. 2008, 36, e76. [Google Scholar] [CrossRef] [Green Version]
- Lin Cereghino, G.P.; Sunga, A.J.; Cereghino, J.L.; Cregg, J.M. Expression of Foreign Genes in the Yeast Pichia Pastoris. Genet. Eng. Princ. Methods 2001, 23, 157–169. [Google Scholar]
- Cregg, J.M.; Cereghino, J.L.; Shi, J.; Higgins, D.R. Recombinant Protein Expression in Pichia Pastoris. Mol. Biotechnol. 2000, 16, 23–52. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, G.P.L.; Cereghino, J.L.; Ilgen, C.; Cregg, J.M. Production of Recombinant Proteins in Fermenter Cultures of the Yeast Pichia Pastoris. Curr. Opin. Biotechnol. 2002, 13, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, J.L.; Cregg, J.M. Heterologous Protein Expression in the Methylotrophic Yeast Pichia Pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Shental-Bechor, D.; Levy, Y. Effect of Glycosylation on Protein Folding: A Close Look at Thermodynamic Stabilization. Proc. Natl. Acad. Sci. USA 2008, 105, 8256–8261. [Google Scholar] [CrossRef] [Green Version]
- Jayaprakash, N.G.; Surolia, A. Role of Glycosylation in Nucleating Protein Folding and Stability. Biochem. J. 2017, 474, 2333–2347. [Google Scholar] [CrossRef]
- Roth, J.; Zuber, C.; Park, S.; Jang, I.; Lee, Y.; Kysela, K.G. Protein N-Glycosylation, Protein Folding, and Protein Quality Control. Molecules 2010, 30, 497–506. [Google Scholar] [CrossRef]
- Gao, J.; Jiang, L.; Lian, J. Development of Synthetic Biology Tools to Engineer Pichia Pastoris as a Chassis for the Production of Natural Products. Synth. Syst. Biotechnol. 2021, 6, 110–119. [Google Scholar] [CrossRef]
- Gaboardi, G.C.; Alves, D.; Gil de Los Santos, D.; Xavier, E.; Nunes, A.P.; Finger, P.; Griep, E.; Roll, V.; Oliveira, P.; Silva, A.; et al. Influence of Pichia Pastoris X-33 Produced in Industrial Residues on Productive Performance, Egg Quality, Immunity, and Intestinal Morphometry in Quails. Sci. Rep. 2019, 9, 15372. [Google Scholar] [CrossRef] [Green Version]
- Sturmberger, L.; Chappell, T.; Geier, M.; Krainer, F.; Day, K.J.; Vide, U.; Trstenjak, S.; Schiefer, A.; Richardson, T.; Soriaga, L.; et al. Refined Pichia Pastoris Reference Genome Sequence. J. Biotechnol. 2016, 235, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Sinha, J.; Plantz, B.A.; Zhang, W.; Gouthro, M.; Schlegel, V.; Liu, C.-P.; Meagher, M.M. Improved Production of Recombinant Ovine Interferon-Tau by Mut(+) Strain of Pichia Pastoris Using an Optimized Methanol Feed Profile. Biotechnol. Prog. 2003, 19, 794–802. [Google Scholar] [CrossRef]
- Files, D.; Ogawa, M.; Scaman, C.H.; Baldwin, S.A. A Pichia Pastoris Fermentation Process for Producing High-Levels of Recombinant Human Cystatin-C. Enzym. Microb. Technol. 2001, 29, 335–340. [Google Scholar] [CrossRef]
- Cos, O.; Ramón, R.; Montesinos, J.L.; Valero, F. Operational Strategies, Monitoring and Control of Heterologous Protein Production in the Methylotrophic Yeast Pichia Pastoris under Different Promoters: A Review. Microb. Cell Fact. 2006, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamawaki, S.; Matsumoto, T.; Ohnishi, Y.; Kumada, Y.; Shiomi, N.; Katsuda, T.; Lee, E.K.; Katoh, S. Production of Single-Chain Variable Fragment Antibody (ScFv) in Fed-Batch and Continuous Culture of Pichia Pastoris by Two Different Methanol Feeding Methods. J. Biosci. Bioeng. 2007, 104, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Looser, V.; Bruhlmann, B.; Bumbak, F.; Stenger, C.; Costa, M.; Camattari, A.; Fotiadis, D.; Kovar, K. Cultivation Strategies to Enhance Productivity of Pichia Pastoris: A Review. Biotechnol. Adv. 2015, 33, 1177–1193. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.R.; Ataíde, F.; von Stosch, M.; Dias, J.M.L.; Clemente, J.J.; Cunha, A.E.; Oliveira, R. Application of Adaptive DO-Stat Feeding Control to Pichia Pastoris X33 Cultures Expressing a Single Chain Antibody Fragment (ScFv). Bioprocess Biosyst. Eng. 2012, 35, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Trinh, L.B.; Phue, J.N.; Shiloach, J. Effect of Methanol Feeding Strategies on Production and Yield of Recombinant Mouse Endostatin from Pichia Pastoris. BioTechnology 2003, 82, 438–444. [Google Scholar]
- Liu, W.; Xiang, H.; Zhang, T.; Pang, X.; Su, J.; Liu, H. Development of a New High-Cell Density Fermentation Strategy for Enhanced Production of a Fungus β-Glucosidase in Pichia Pastoris. Front. Microbiol. 2020, 11, 1988. [Google Scholar] [CrossRef] [PubMed]
- Dalvie, N.C.; Rodriguez-Aponte, S.A. Engineered SARS-CoV-2 Receptor Binding Domain Improves Manufacturability in Yeast and Immunogenicity in Mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2106845118. [Google Scholar] [CrossRef]
- Liu, Y.; Ye, Q. Nucleic Acid Vaccines against SARS-CoV-2. Vaccines 2022, 10, 1849. [Google Scholar] [CrossRef]
- Jain, S.; Venkataraman, A.; Wechsler, M.E.; Peppas, N.A. Messenger RNA-Based Vaccines: Past, Present, and Future Directions in the Context of the COVID-19 Pandemic. Adv. Drug Deliv. Rev. 2021, 179, 114000. [Google Scholar] [CrossRef]
- Heidary, M.; Kaviar, V.H.; Shirani, M.; Ghanavati, R.; Motahar, M.; Sholeh, M.; Ghahramanpour, H.; Khoshnood, S. A Comprehensive Review of the Protein Subunit Vaccines Against COVID-19. Front. Microbiol. 2022, 13, 927306. [Google Scholar] [CrossRef] [PubMed]
- Dalvie, N.C.; Tostanoski, L.H.; Rodriguez-Aponte, S.A.; Kaur, K.; Bajoria, S.; Kumru, O.S.; Martinot, A.J.; Chandrashekar, A.; McMahan, K.; Mercado, N.B.; et al. SARS-CoV-2 Receptor Binding Domain Displayed on HBsAg Virus-like Particles Elicits Protective Immunity in Macaques. Sci. Adv. 2022, 8, eabl6015. [Google Scholar] [CrossRef] [PubMed]
- Ghasemiyeh, P.; Mohammadi-Samani, S.; Firouzabadi, N.; Dehshahri, A.; Vazin, A. A Focused Review on Technologies, Mechanisms, Safety, and Efficacy of Available COVID-19 Vaccines. Int. Immunopharmacol. 2021, 100, 108162. [Google Scholar] [CrossRef] [PubMed]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In Protein Protocols Handbook; Humana: Louisville, KY, USA, 2009; pp. 15–22. [Google Scholar]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2009; pp. 17–24. ISBN 9781603274746. [Google Scholar]
- Chen, W.-H.; Chag, S.M.; Poongavanam, M.V.; Biter, A.B.; Ewere, E.A.; Rezende, W.; Seid, C.A.; Hudspeth, E.M.; Pollet, J.; McAtee, C.P.; et al. Optimization of the Production Process and Characterization of the Yeast-Expressed SARS-CoV Recombinant Receptor-Binding Domain (RBD219-N1), a SARS Vaccine Candidate. J. Pharm. Sci. 2017, 106, 1961–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Xiong, F.; Lin, Q.; d’Anjou, M.; Daugulis, A.J.; Yang, D.S.; Hew, C.L. Low-Temperature Increases the Yield of Biologically Active Herring Antifreeze Protein in Pichia Pastoris. Protein Expr. Purif. 2001, 21, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Karkut, T.; Chamankhah, M.; Alting-Mees, M.; Hemmingsen, S.M.; Hegedus, D. Optimal Conditions for the Expression of a Single-Chain Antibody (ScFv) Gene in Pichia Pastoris. Protein Expr. Purif. 2003, 28, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Celik, E.; Calık, P. Production of Recombinant Proteins by Yeast Cells. Biotechnol. Adv. 2012, 30, 1108–1118. [Google Scholar] [CrossRef]
- Noseda, D.G.; Recúpero, M.N.; Blasco, M.; Ortiz, G.E.; Galvagno, M.A. Cloning, Expression and Optimized Production in a Bioreactor of Bovine Chymosin B in Pichia (Komagataella) Pastoris under AOX1 Promoter. Protein Expr. Purif. 2013, 92, 235–244. [Google Scholar] [CrossRef]
- Noseda, D.G.; Recúpero, M.; Blasco, M.; Bozzo, J.; Galvagno, M.Á. Production in Stirred-Tank Bioreactor of Recombinant Bovine Chymosin B by a High-Level Expression Transformant Clone of Pichia Pastoris. Protein Expr. Purif. 2016, 123, 112–121. [Google Scholar] [CrossRef]
- Picotto, L.D.; Sguazza, G.H.; Tizzano, M.A.; Galosi, C.M.; Cavalitto, S.F.; Pecoraro, M.R. An Effective and Simplified DO-Stat Control Strategy for Production of Rabies Glycoprotein in Pichia Pastoris. Protein Expr. Purif. 2017, 132, 124–130. [Google Scholar] [CrossRef]
- Kim, S.; Warburton, S.; Boldogh, I.; Svensson, C.; Pon, L.; d’Anjou, M.; Stadheim, T.A.; Choi, B.-K. Regulation of Alcohol Oxidase 1 (AOX1) Promoter and Peroxisome Biogenesis in Different Fermentation Processes in Pichia Pastoris. J. Biotechnol. 2013, 166, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Looser, V.; Lüthy, D.; Straumann, M.; Hecht, K.; Melzoch, K.; Kovar, K. Effects of Glycerol Supply and Specific Growth Rate on Methanol-Free Production of CALB by P. Pastoris: Functional Characterisation of a Novel Promoter. Appl. Microbiol. Biotechnol. 2017, 101, 3163–3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capone, S.; Horvat, J.; Herwig, C.; Spadiut, O. Development of a Mixed Feed Strategy for a Recombinant Pichia Pastoris Strain Producing with a De-Repression Promoter. Microb. Cell Fact. 2015, 14, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, M.-M.; Rieley, W. Medical Gases, Their Storage and Delivery. Anaesth. Intensive Care Med. 2012, 13, 533–538. [Google Scholar] [CrossRef]
- Malayaman, S.N.; Mychaskiw, G.I.I.; Berry, J.M.; Ehrenwerth, J. Medical Gases: Storage and Supply. In Anesthesia Equipment; Elsevier: Amsterdam, The Netherlands, 2021; pp. 3–24. [Google Scholar]
- Farnós, O.; Venereo-Sánchez, A.; Xu, X.; Chan, C.; Dash, S.; Chaabane, H.; Sauvageau, J.; Brahimi, F.; Saragovi, U.; Leclerc, D.; et al. Rapid High-Yield Production of Functional SARS-CoV-2 Receptor Binding Domain by Viral and Non-Viral Transient Expression for Pre-Clinical Evaluation. Vaccines 2020, 8, 654. [Google Scholar] [CrossRef]
- Li, W.; Fan, Z.; Lin, Y.; Wang, T.-Y. Serum-Free Medium for Recombinant Protein Expression in Chinese Hamster Ovary Cells. Front. Bioeng. Biotechnol. 2021, 9, 646363. [Google Scholar] [CrossRef]
- Zhang, J.; Greasham, R. Chemically Defined Media for Commercial Fermentations. Appl. Microbiol. Biotechnol. 1999, 51, 407–421. [Google Scholar] [CrossRef]
- Orman, M.A.; Çalık, P.; Özdamar, T.H. The Influence of Carbon Sources on Recombinant-human-growth-hormone Production by Pichia Pastoris Is Dependent on Phenotype: A Comparison of Muts and Mut+ strains. Biotechnol. Appl. Biochem. 2009, 52, 245–255. [Google Scholar] [CrossRef]
- Pla, I.A.; Damasceno, L.M.; Vannelli, T.; Ritter, G.; Batt, C.A.; Shuler, M.L. Evaluation of Mut+ and MutS Pichia Pastoris Phenotypes for High Level Extracellular ScFv Expression under Feedback Control of the Methanol Concentration. Biotechnol. Prog. 2006, 22, 881–888. [Google Scholar] [CrossRef]
- Singh, A.; Narang, A. The Mut+ Strain of Komagataella Phaffii (Pichia Pastoris) Expresses PAOX1 5 and 10 Times Faster than Muts and Mut− Strains: Evidence That Formaldehyde or/and Formate Are True Inducers of PAOX1. Appl. Microbiol. Biotechnol. 2020, 104, 7801–7814. [Google Scholar] [CrossRef]
- Barrigón, J.M.; Montesinos, J.L.; Valero, F. Searching the Best Operational Strategies for Rhizopus Oryzae Lipase Production in Pichia Pastoris Mut+ Phenotype: Methanol Limited or Methanol Non-Limited fed-batch cultures? Biochem. Eng. J. 2013, 75, 47–54. [Google Scholar] [CrossRef]
- Garrigós-Martínez, J.; Nieto-Taype, M.A. Specific Growth Rate Governs AOX1 Gene Expression, Affecting the Production Kinetics of Pichia Pastoris (Komagataella Phaffii) PAOX1-Driven Recombinant producer strains with different target gene dosage. Microb. Cell Fact. 2019, 18, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayson, B.E.; Kilburn, D.G.; Zamost, B.L.; Raymond, C.K.; Lesnicki, G.J. Effects of Methanol Concentration on Expression Levels of Recombinant Protein in Fed-Batch Cultures of Pichia Methanolica. Biotechnol. Bioeng. 2003, 81, 291–298. [Google Scholar] [CrossRef]
- Katakura, Y.; Zhang, W.; Zhuang, G.; Omasa, T. Effect of Methanol Concentration on the Production of Human Β2-Glycoprotein I Domain V by a Recombinant Pichia Pastoris: A Simple System for the Control of Methanol concentration using a semiconductor gas sensor. J. Ferment. Bioeng. 1998, 86, 482–487. [Google Scholar] [CrossRef]
- Guarna, M.M.; Lesnicki, G.J.; Tam, B.M.; Robinson, J.; Radziminski, C.Z.; Hasenwinkle, D.; Boraston, A.; Jervis, E.; MacGillivray, R.T.; Turner, R.F.; et al. On-Line Monitoring and Control of Methanol Concentration in Shake-Flask Cultures of Pichia Pastoris. Biotechnol. Bioeng. 1997, 56, 279–286. [Google Scholar] [CrossRef]
- Schenk, J.; Marison, I.W.; von Stockar, U. A Simple Method to Monitor and Control Methanol Feeding of Pichia Pastoris Fermentations Using Mid-IR Spectroscopy. J. Biotechnol. 2007, 128, 344–353. [Google Scholar] [CrossRef]
- Limonta-Fernández, M.; Chinea-Santiago, G.; Martín-Dunn, A.M.; Gonzalez-Roche, D.; Bequet-Romero, M.; Marquez-Perera, G.; González-Moya, I.; Canaan-Haden-Ayala, C.; Cabrales-Rico, A.; Espinosa-Rodríguez, L.A.; et al. An Engineered SARS-CoV-2 Receptor-Binding Domain Produced in Pichia Pastoris as a Candidate Vaccine Antigen. New Biotechnol. 2022, 72, 11–21. [Google Scholar] [CrossRef]
- Cardoso, V.M.; Campani, G.; Santos, M.P.; Silva, G.G. Cost Analysis Based on Bioreactor Cultivation Conditions: Production of a Soluble Recombinant Protein Using Escherichia Coli BL21 (DE3). Biotechnology 2020, 26, e00441. [Google Scholar] [CrossRef]
- He, Y.; Qi, J.; Xiao, L.; Shen, L.; Yu, W.; Hu, T. Purification and Characterization of the Receptor-Binding Domain of SARS-CoV-2 Spike Protein from Escherichia Coli. Eng. Life Sci. 2021, 21, 453–460. [Google Scholar] [CrossRef]
- Meena, J.; Singhvi, P.; Srichandan, S.; Dandotiya, J.; Verma, J.; Singh, M.; Ahuja, R.; Panwar, N.; Wani, T.Q.; Khatri, R.; et al. RBD Decorated PLA Nanoparticle Admixture with Aluminum Hydroxide Elicit Robust and Long Lasting Immune Response against SARS-CoV-2. Eur. J. Pharm. Biopharm. 2022, 176, 43–53. [Google Scholar] [CrossRef]
- Hossler, P.; Khattak, S.F.; Li, Z.J. Optimal and Consistent Protein Glycosylation in Mammalian Cell Culture. Glycobiology 2009, 19, 936–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyoncu, S.; Yilmaz, S.; Kuyucu, A.Z.; Sayili, D.; Mert, O.; Soyturk, H.; Gullu, S.; Akinturk, H.; Citak, E.; Arslan, M.; et al. Process Development for an Effective COVID-19 Vaccine Candidate Harboring Recombinant SARS-CoV-2 Delta plus Receptor Binding Domain Produced by Pichia Pastoris. Sci. Rep. 2023, 13, 5224. [Google Scholar] [CrossRef] [PubMed]
- Dumont, J.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R. Human Cell Lines for Biopharmaceutical Manufacturing: History, Status, and Future Perspectives. Crit. Rev. Biotechnol. 2016, 36, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, B.; Gottschalk, U.; Pohlscheidt, M. New Bioprocessing Strategies: Development and Manufacturing of Recombinant Antibodies and Proteins; Springer: Berlin/Heidelberg, Germany, 2018; ISBN 9783319971100. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Induction Time (h) | Total Protein Concentration (mg/L) | RBD Concentration (mg/L) | RBD Increase (fold) | Y RBD/x (mg/g) | vP (mg/L h) | sP (µg/g h) |
|---|---|---|---|---|---|---|
| 0 | 5.2 | 1.6 | 1.0 | 0.34 | 0.07 | 14.1 |
| 24 | 12.8 | 3.8 | 2.5 | 0.61 | 0.08 | 12.7 |
| 48 | 26.4 | 7.9 | 5.7 | 1.12 | 0.11 | 15.5 |
| 72 | 41.8 | 12.5 | 8.0 | 1.67 | 0.13 | 17.4 |
| 96 | 57.1 | 17.1 | 11.0 | 2.17 | 0.14 | 18.1 |
| 120 | 72.3 | 21.7 | 13.9 | 2.71 | 0.15 | 18.8 |
| Strategy 1 | Strategy 2 | |
|---|---|---|
| Final biomass level (g DCW/L) | 78.2 | 89.2 |
| Total protein concentration (mg/L) | 296.3 | 1378.5 |
| RBD concentration (mg/L) | 98.4 | 519.6 |
| Total RBD (mg) | 255.8 | 1402.9 |
| YRBD/Biomass (mg/g) | 1.3 | 5.8 |
| Volumetric RBD productivity (mg/L h) | 1.0 | 3.8 |
| Total RBD productivity (mg/h) | 2.6 | 10.3 |
| Specific RBD productivity (µg/g DCW h) | 12.8 | 42.2 |
| YRBD/Methanol (mg/g) | 0.5 | 1.8 |
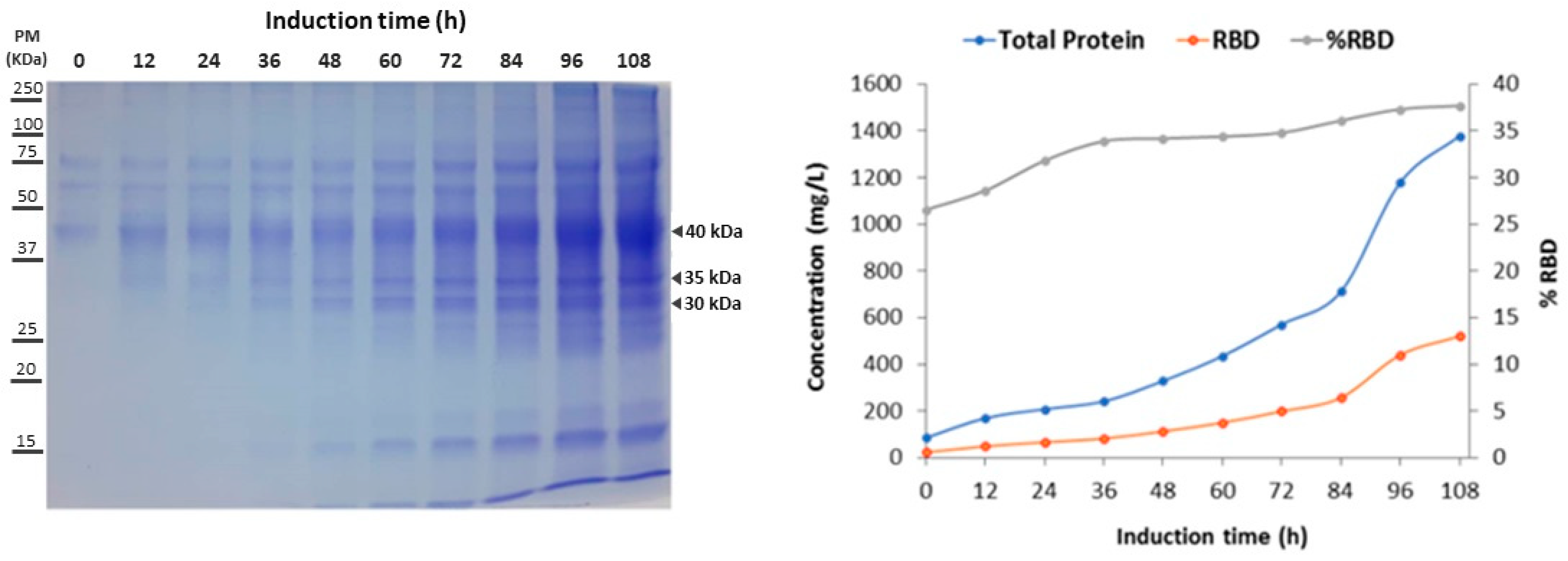
| Induction Time (h) | Total Protein Concentration (mg/L) | RBD Concentration (mg/L) | RBD Percentage (%) | RBD Increase (fold) | Y RBD/x (mg/g) vP | vP (mg/L h) |
|---|---|---|---|---|---|---|
| 0 | 85.0 | 22.6 | 26.5 | 1.0 | 0.5 | 0.8 |
| 12 | 168.7 | 48.2 | 28.6 | 2.1 | 0.8 | 1.2 |
| 24 | 207.7 | 66.0 | 31.8 | 2.9 | 1.0 | 1.2 |
| 36 | 241.0 | 81.7 | 33.9 | 3.6 | 1.1 | 1.3 |
| 48 | 328.5 | 112.3 | 34.2 | 5.0 | 1.4 | 1.5 |
| 60 | 435.0 | 149.6 | 34.4 | 6.6 | 1.9 | 1.7 |
| 72 | 570.6 | 198.6 | 34.8 | 8.8 | 2.4 | 2.0 |
| 84 | 711.4 | 256.8 | 36.1 | 11.4 | 2.9 | 2.3 |
| 96 | 1178.3 | 439.8 | 37.3 | 19.5 | 4.9 | 3.5 |
| 108 | 1378.5 | 519.6 | 37.7 | 23.0 | 5.8 | 3.8 |
| Induction Time (h) | Total Protein Concentration (mg/L) | RBD Concentration (mg/L) | RBD Percentage (%) | RBD Increase (fold) | Y RBD/x (mg/g) vP | vP (mg/L h) |
|---|---|---|---|---|---|---|
| 0 | 120.3 | 37.3 | 31.0 | 1.0 | 1.0 | 1.2 |
| 24 | 169.7 | 65.2 | 38.4 | 1.7 | 1.1 | 1.2 |
| 48 | 237.0 | 101.2 | 42.7 | 2.7 | 1.4 | 1.3 |
| 72 | 415.3 | 188.1 | 45.3 | 5.0 | 2.4 | 1.8 |
| 96 | 669.2 | 321.8 | 48.1 | 8.6 | 3.9 | 2.6 |
| 115 | 1032.7 | 533.4 | 51.7 | 14.3 | 5.9 | 3.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noseda, D.G.; D’Alessio, C.; Santos, J.; Idrovo-Hidalgo, T.; Pignataro, F.; Wetzler, D.E.; Gentili, H.; Nadra, A.D.; Roman, E.; Paván, C.; et al. Development of a Cost-Effective Process for the Heterologous Production of SARS-CoV-2 Spike Receptor Binding Domain Using Pichia pastoris in Stirred-Tank Bioreactor. Fermentation 2023, 9, 497. https://doi.org/10.3390/fermentation9060497
Noseda DG, D’Alessio C, Santos J, Idrovo-Hidalgo T, Pignataro F, Wetzler DE, Gentili H, Nadra AD, Roman E, Paván C, et al. Development of a Cost-Effective Process for the Heterologous Production of SARS-CoV-2 Spike Receptor Binding Domain Using Pichia pastoris in Stirred-Tank Bioreactor. Fermentation. 2023; 9(6):497. https://doi.org/10.3390/fermentation9060497
Chicago/Turabian StyleNoseda, Diego G., Cecilia D’Alessio, Javier Santos, Tommy Idrovo-Hidalgo, Florencia Pignataro, Diana E. Wetzler, Hernán Gentili, Alejandro D. Nadra, Ernesto Roman, Carlos Paván, and et al. 2023. "Development of a Cost-Effective Process for the Heterologous Production of SARS-CoV-2 Spike Receptor Binding Domain Using Pichia pastoris in Stirred-Tank Bioreactor" Fermentation 9, no. 6: 497. https://doi.org/10.3390/fermentation9060497